
Abstract
Osteosarcoma (OS) is the most common primary bone tumor in children and adolescents. Long noncoding RNAs (lncRNAs) are a class of transcriptional products of the genome without protein-coding potential. Recently, lncRNA Ewing sarcoma-associated transcript 1 (EWSAT1) was functionally identified in Ewing sarcoma, a highly aggressive primary pediatric bone tumor. However, whether EWSAT1 plays a role in OS remains unclear. In the present study, gain- and loss-of-function assays demonstrated that EWSAT1 enhanced OS cell proliferation, migration, and invasion. Further mechanistic studies found that EWSAT1 positively regulated lncRNA MEG3 expression in the transcriptional level. Finally, we observed that EWSAT1 facilitates OS cell growth and metastasis through regulation of MEG3, suggesting that EWSAT1-MEG3 axis might be a promising target for OS treatment.
Introduction
O
Long noncoding RNAs (lncRNAs) are a class of transcriptional products of the genome without protein-coding potential and composed of more than 200 nucleotides in length (Serghiou et al., 2016). lncRNAs play a critical role in the regulation of gene expression in various manners, such as transcription control, chromatin remodeling, and post-transcriptional regulation through interaction with protein, microRNAs, or mRNAs (Bartonicek et al., 2016; Sullenger and Nair, 2016). Recent comprehensive studies have demonstrated the gene expression profiles of lncRNAs in cancers through microarray or RNA sequencing. For example, downregulation of lncRNA MEG3 has been observed in various cancers and was associated with poor prognosis (Lu et al., 2013; Qin et al., 2013; Sun et al., 2014). lncRNA MEG3 functions as a tumor suppressor gene, which regulated p53 expression and the TGF-β pathway genes (Lu et al., 2013; Mondal et al., 2015). However, only a few lncRNAs have been functionally annotated in OS (Li et al., 2016; Ruan et al., 2016). Recently, lncRNA Ewing sarcoma-associated transcript 1 (EWSAT1) was functionally identified in Ewing sarcoma, a highly aggressive primary pediatric bone tumor. EWSAT1 was upregulated by EWS-FLI1 fusion protein in human mesenchymal progenitor cells. Silence of EWSAT1 expression suppressed the proliferation of Ewing sarcoma cells. EWSAT1 regulated target genes through interaction with heterogeneous nuclear ribonucleoprotein (HNRNPK), which was an RNA-binding protein (Marques Howarth et al., 2014). However, whether EWSAT1 plays a role in OS remains unclear.
In the present study, we demonstrated that knockdown of EWSAT1 suppressed cell proliferation, migration, and invasion in OS cells. In contrast, overexpression of EWSAT1 promoted cell growth, migration, and invasion in OS cells. Furthermore, we found that EWSAT1 positively regulated lncRNA MEG3 expression in the transcriptional level. Finally, we observed that EWSAT1 facilitates OS cell growth and metastasis through regulation of MEG3. We supposed that the EWSAT1-MEG3 axis might be a promising target for OS treatment.
Materials and Methods
Cell culture
The human osteosarcoma (HOS) cell lines, MG63 and HOS cells, were purchased from American Type Culture Collection (ATCC). Cells were cultured in Dulbecco's Modified Eagle's Medium (DMEM) (Gibco, America) supplemented with 10% fetal bovine serum (FBS) (Gibco, America), 100 U penicillin, 100 μg/mL streptomycin, and 100 U pen/100 μg strep/2 mM glutamine (Invitrogen).
Knockdown and overexpression experiments
The pLKO.1 shRNA lentiviral system was used to knockdown genes of interest. pLKO.1-shScramble, pLKO.1-shEWSAT1-1, and pLKO.1-shEWSAT1-2 were purchased from Open Biosystems (America). The target sequences of EWSAT1 were as follows: sh1: TTGGGCTCTCAATGGT ATCAT and sh2: AAGGGAGGGTTACTAACTTTA. Lentivirus expressing these shRNAs was produced as previously described (Wang et al., 2015b). Cells were infected and then selected with 3 μg/mL puromycin for 7 days.
For the case of overexpression experiments, cells were infected with lentivirus expressing full-length EWSAT1 and selected with 3 μg/mL puromycin for 7 days.
siRNA transfection
siRNAs against MEG3 were purchased from RiboBio company (Guangzhou, China) and transfected using Lipofectamine 3000 (Invitrogen) according to the manufacturer's instructions. Cells were transfected with 100 nM siRNA for 48 h before following assays. The sequences of siRNAs were as follows: siMEG3: GGAAUGAGCAUGCUACUGAAU and siHNRNPK: GCAGUAUUCUGGAAAGUUUUU.
Luciferase assays
The promoter region of MEG3 was cloned into pGL3-basic vector. Cells were grown in 24-well plates to ∼60% confluence and then transfected with luciferase reporter construct containing MEG3 promoter region and EWSAT1 expression plasmid or EWSAT1 shRNA plasmid by TurboFect transfection reagent (Thermo). At 48 h after transfection, cells were lysed and luciferase activities determined by a dual-luciferase reporter gene assay system (Promega, America) according to the manufacturer's instructions.
RNA isolation and real-time polymerase chain reaction
Total RNA was isolated with the TRIzol (Invitrogen, America) reagent according to the manufacturer's instructions. Approximately 1 μg RNA was reverse transcribed using the Reverse EasyScript One-Step gDNA Removal and cDNA Synthesis SuperMix (Trangene, Beijing, China) following the manufacturer's instructions. Real-time polymerase chain reaction (RT-PCR) was performed using the SYBR Green Master Mixture (Roche, America) reagent in ABI 7500 Real-time PCR instrument. GAPDH was used as an endogenous control. The relative expression of genes was calculated by the 2−ΔΔCt method. Primers used for RT-RCR are listed as follows: EWSAT1-F: GTGTCTGGCAAGGAA CACTA, EWSAT1-R: GGTGGAGAAGAGGGACAATAAG; MEG3-F: CCTCACCTCCAATTTCCTCTTC, MEG3-R: CTTCCATCCGCAGTTCTTCA; and HNRNPK-F: TGAT TGGTGTGCCCGTTTA, HNRNPK-R: GCAGGGCGTTT ACCAAATTC.
RNA immunoprecipitation
RNA immunoprecipitation (RIP) assays were performed by using an EZ-Magna RIP RNA-binding protein immunoprecipitation kit (Millipore, America) according to the standard protocol. Cells were mixed with 2 mL nuclear lysis buffer. Also, then nuclei were pelleted by centrifugation at 800 g for 10 min. Pellets were resuspended in freshly prepared RIP buffer and then incubated on ice for 20 min with frequent mixing. The nuclear membranes and debris were pelleted by centrifugation at 12,000 g for 10 min at 4°C. Five micrograms HNRNPK (ab70492; Abcam) or control IgG (Cell Signaling Technology) antibodies and beads were used for RIP reaction. After wash, the RIP products were extracted and analyzed by RT-PCR.
Colony formation assay
Cells (2 × 103) per well were seeded in 6-well plates. The cells were cultured for 14 days. The cells were then fixed and staining by crystal violet. All experiments were performed at least three times in triplicate.
CCK-8 assays
Cells (3 × 103) per well were seeded in 96-well plates, and the viability of the cells was detected from three replicates by the CCK-8 assay kit (Dojindo, Japan) at indicated time point.
Migration and invasion assay
For migration assay, 1.0 × 105 cells were seeded in FBS-free DMEM in the upper chamber of a 24-well transwell (Corning, America). For invasion assay, 3.0 × 105 cells were seeded in FBS-free DMEM in the upper chamber of a 24-well Matrigel transwell invasion insert (BD bioscience, America). The lower chamber was filled with DMEM supplemented with 20% FBS. After 24 h of incubation, cells in the upper chamber were removed, and the cells that had traversed the membrane were staining by crystal violet. All experiments were performed at least three times in triplicate.
Statistics
p-Values were calculated using two-tailed Student's t-test or one-way ANOVA test by SPSS 19.0 software. A p-value of less than 0.05 was considered significant.
Results
EWSAT1 promotes proliferation of OS cells
To determine the biological function of EWSAT1 in OS cells in vitro, we constructed OS cell lines (MG63 and HOS) with stable overexpression or knockdown of EWSAT1 using the lentiviral expressing system (Fig. 1A and B). First, we evaluated the effect of EWSAT1 on OS cell growth by performing colony formation assay. The results demonstrated that knockdown of EWSAT1 significantly decreased the clonogenic survival of MG63 and HOS cells compared with their control cells (Fig. 1C). In contrast, EWSAT1-overexpressed cells showed a significant increase in the clonogenic survival of both MG63 and HOS cells compared with their control cells (Fig. 1D). For further confirmation, we performed the CCK8 assay to detect the effect of EWSAT1 on cell proliferation, and the results showed that knockdown of EWSAT1 significantly suppressed MG63 and HOS cell vitality (Fig. 1E), while EWSAT1 overexpression markedly enhanced the vitality of MG63 and HOS cells (Fig. 1F). Taken together, EWSAT1 promotes cell growth of OS cells in vitro.

EWSAT1 promotes proliferation of OS cells.
EWSAT1 promotes OS cell metastasis in vitro
The poor prognosis of OS patients is mainly due to the cancer cell metastasis. To investigate whether the EWSAT1 plays a critical role in OS cell metastasis, we performed transwell assays. We found that overexpression of EWSAT1 promoted cell migration, while knockdown of EWSAT1 suppressed cell migration in both MG63 and HOS cells (Fig. 2A and B). Similarly, the invasion was significantly suppressed in EWSAT1 knockdown cells compared with their control cells, while EWSAT1 overexpression elevated cell invasion in MG63 and HOS cells (Fig. 2C and D). These data suggested that EWSAT1 promoted OS migration and invasion.
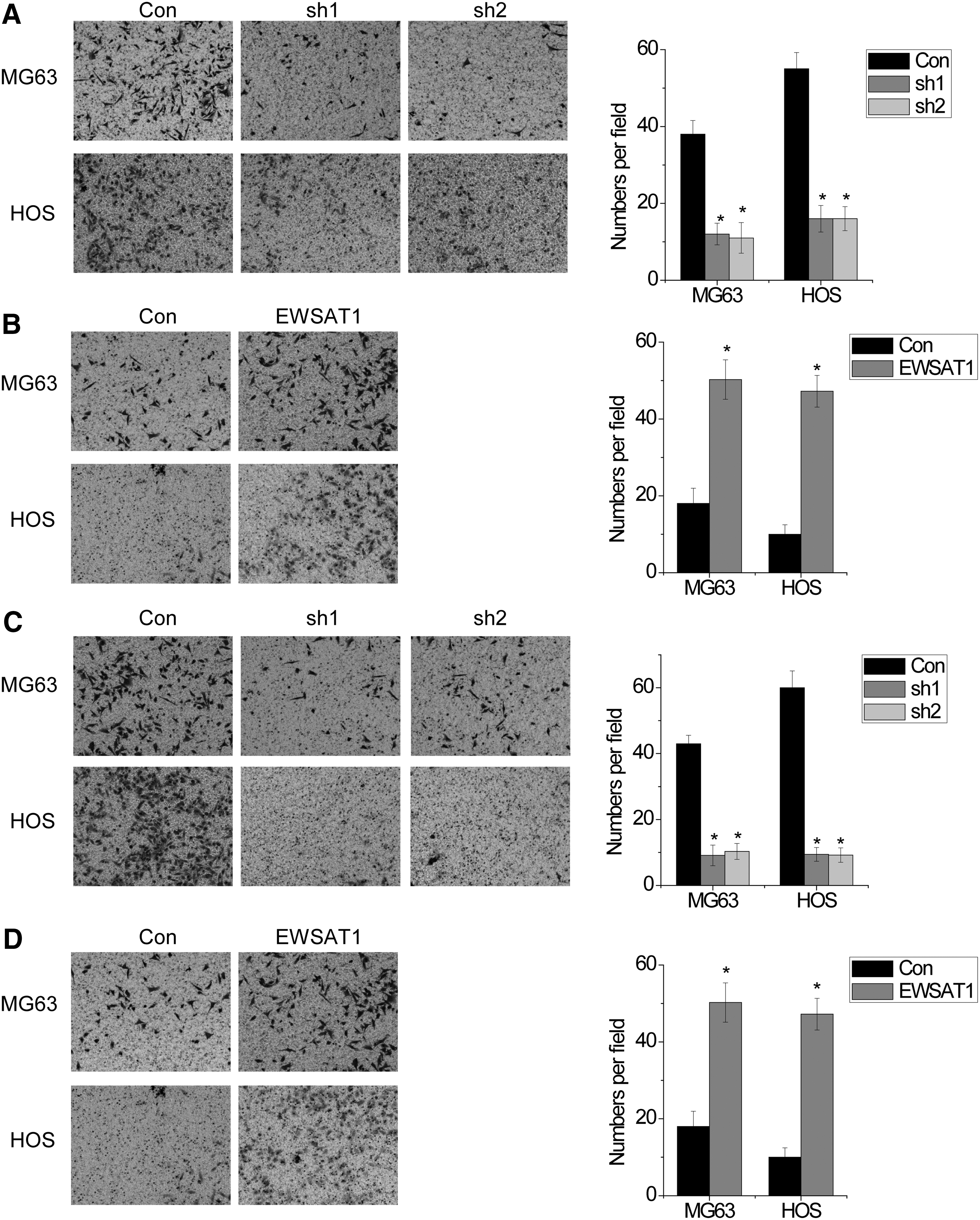
EWSAT1 promotes OS cell metastasis in vitro.
lncRNA MEG3 is a direct target gene of EWSAT1
Next, we explored the exact mechanism by which EWSAT1 promotes malignant phenotypes of OS cells. We analyzed the gene expression profiles mediated by EWSAT1 from the previous study (Marques Howarth et al., 2014). We found that lncRNA MEG3 might be a direct target gene of EWSAT1. To confirm these hypotheses, we detected the MEG3 expression in EWSAT1-overexpressed or EWSAT1-silenced cells by RT-PCR. We found that MEG3 expression was significantly downregulated by EWSAT1 overexpression, while markedly increased by EWSAT1 knockdown (Fig. 3A and B). Furthermore, we cloned the promoter region of MEG3 into luciferase reporter vector. We found that overexpression of EWSAT1 significantly suppressed the luciferase activity of MEG3 promoter (Fig. 3C). In contrast, silence of EWSAT1 increased the luciferase activity of MEG3 promoter (Fig. 3D). These data indicated that EWSAT1 regulated MEG3 expression in the transcriptional level.

lncRNA MEG3 is a direct target gene of EWSAT1.
Next, we determined the mechanism by which EWSAT1 regulates MEG3 expression. Previous study demonstrated that HNRNPK was associated with EWSAT1 in Ewing sarcoma (Marques Howarth et al., 2014). We suspected that MEG3 was also regulated by EWSAT1 through HNRNPK in OS. Consistent with this study, RIP assays showed that EWSAT1 was significantly enriched by the HNRNPK antibody in both MG63 and HOS cells compared with IgG and negative control GAPDH (Fig. 3E). Furthermore, we determined whether MEG3 expression regulated by EWSAT1 depended on HNRNPK. We found that HNRNPK silencing abrogated suppression of MEG3 expression and luciferase activity of MEG3 promoter mediated by EWSAT1 overexpression in MG63 and HOS cells (Fig. 3F–H). These data demonstrated that MEG3 was regulated by EWSAT1 through the interaction of EWSAT1 and HNRNPK.
EWSAT1 promotes malignant phenotypes of OS cells through regulation of MEG3
Finally, we explored whether EWSAT1 facilitated malignant phenotypes of OS cells through MEG3. Full-length MEG3 was transfected into MG63 cells with EWSAT1 overexpression (Fig. 4A). Interestingly, we found that MEG3 overexpression abrogated the effect of EWSAT1 overexpression in inducing cell proliferation, migration, and invasion in MG63 cells (Fig. 4B and C). In contrast, MEG3 silencing rescued the inhibition of cell proliferation, migration, and invasion mediated by EWSAT1 knockdown in MG63 cells, indicating that the function of EWSAT1 in OS cells depends on MEG3 (Fig. 4D–F). Collectively, these data indicated that EWSAT1 promotes malignant phenotypes in a MEG3-dependent manner.
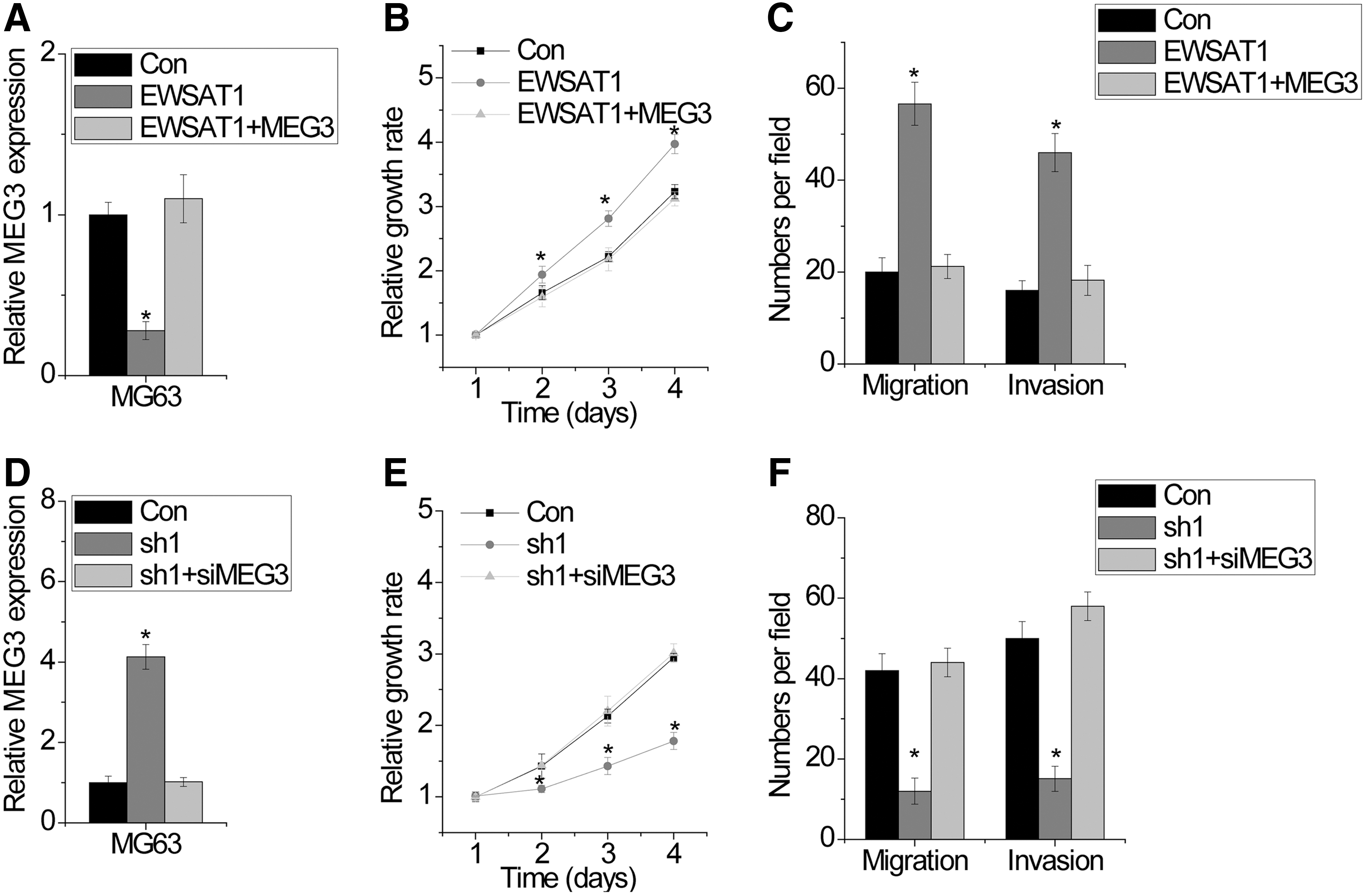
EWSAT1 promotes malignant phenotypes of OS cells through MEG3.
Discussion
Increasing evidence suggests that lncRNAs play important roles in tumorigenesis and cancer progression (Schmitt and Chang, 2016; Serghiou et al., 2016). For example, several lncRNAs have been shown to play precise roles in regulating target gene expression in different kind of cancers, including OS, breast cancer, and hepatocellular carcinoma (Dhamija and Diederichs, 2016; Fang and Fullwood, 2016). In hepatocellular carcinoma, lncRNA-HEIH regulated cell cycle through association with EZH2, which was required for the repression of PRC2 target genes (Yang et al., 2011). The copy number and expression level of lncRNA-FAL1 are correlated with clinical prognosis of patients with ovarian cancer, which is associated with the epigenetic repressor BMI1 and regulates its protein stability to regulate the transcription of various genes through epigenetic mechanism (Hu et al., 2014). These and our findings indicated that lncRNAs are critical for oncogenic processes. LncRNAs may be a potential target for cancer therapy.
To the best of our knowledge, the role of lncRNA EWSAT1 in OS has not been described. EWSAT1 promoted growth of Ewing sarcoma cells (Marques Howarth et al., 2014). In this study, we demonstrated a similar function of EWSAT1 in the proliferation of OS cells. EWSAT1 overexpression resulted in significant increases in cell proliferation and colony formation. Conversely, EWSAT1 silence led to the suppression of these phenotypes. In addition, for the first time, we found a novel role of EWSAT1 on OS metastasis. EWSAT1 overexpression was positively correlated with cell migration and invasion in OS cell in vitro cultures. Our findings suggested that EWSAT1 is one of the critical lncRNAs that contributed to the OS growth and metastasis.
MEG3 is an imprinted gene that belongs to the imprinted DLK1–MEG3 locus located at chromosome 14q32.3 in humans. MEG3 has been proved to be a tumor suppressor gene. MEG3 expression is frequently downregulated in many kinds of human cancers, such as cervical cancer, liver cancer, and gastric cancer (Marques Howarth et al., 2014; Zhu et al., 2015; Peng et al., 2015). Genetic and epigenetic mechanisms contribute to the silence of MEG3 expression in cancers (Modali et al., 2015). Restoring MEG3 expression inhibits cancer cell growth and metastasis (Wang et al., 2015a; Zhou et al., 2015). In the present study, for the first time, we found that lncRNA MEG3 was also regulated by another lncRNA, EWSAT1. EWSAT1 regulated MEG3 expression through association with HNRNPK in a transcriptional level. EWSAT1 promoted malignant phenotypes in a MEG3-dependent manner. However, the exact underlying mechanisms by which the EWSAT1-HNRNPK complex regulated MEG3 expression remain unclear. Understanding the exact molecular mechanism will be helpful for OS treatment by targeting the EWSAT1-MEG3 axis.
In summary, we found that EWSAT1 plays a critical role in cell growth and metastasis of OS cells through regulation of MEG3 expression. EWSAT1-MEG3 may also be a potential therapeutic target for OS.
Footnotes
Disclosure Statement
No competing financial interests exist.