
Abstract
Exposure to UVB radiation induces inflammation and free radical-mediated oxidative stress through reactive oxygen species (ROS) that play a crucial role in the induction of skin cancer. Glycolic acid (GA) is frequently used in cosmetics and dermatology. The aim of the study was to analyze the photoprotective mechanisms through which GA retards UVB-induced ROS accumulation and inflammation in normal human epidermal keratinocytes (NHEKs) and mice skin, respectively. NHEK cell line and C57BL/6J mice were treated with GA (0.1 or 5 mM) for 24 h followed by UVB irradiation. ROS accumulation, DNA damage, and expression of inflammasome complexes (NLRP3, NLRC4, ASC, and AIM2) were measured in vitro. Epidermal thickness and inflammasome complex proteins were analyzed in vivo. GA significantly prevented UVB-induced loss of skin cell viability, ROS formation, and DNA damage (single and double strands DNA break). GA suppressed the mRNA expression levels of NLRC4 and AIM2 among the inflammasome complexes. GA also blocked interleukin (IL)-1β by reducing the activity of caspase-1 in the NHEKs. Treatment with GA (2%) inhibited UVB-induced inflammation marker NLRC4 protein levels in mouse dorsal skin. The photoprotective activity of GA was ascribed to the inhibition of ROS formation and DNA damage, as well as a reduction in the activities of inflammasome complexes and IL-1β. We propose that GA has anti-inflammatory and photoprotective effects against UVB irradiation. GA is potentially beneficial to the protection of human skin from UV damage.
Introduction
A
Keratinocytes are the major cell population in the epidermis, constituting approximately 90% of all epidermal cells (Seo et al., 2015). Adverse reactions such as redness, swelling, burning, and pruritus caused by the use of AHA-containing products have been reported (Kurtzweil, 1998). Some investigators demonstrated that topical application of GA (10%) induced photosensitivity by reducing the minimal erythema dose and increasing the number of sunburn cells (Kaidbey et al., 2003; Kornhauser et al., 2009). By contrast, topical GA treatments on human skin providing photoprotective and antioxidative effects have been reported (Perricone and DiNardo, 1996; Morreale and Livrea, 1997). We have shown that AHAs (GA and LA) exhibit antiproliferative effects through the induction of apoptosis and cell cycle arrest in a human keratinocyte cell line (HaCaT cells) (Hong et al., 2001; Ahn et al., 2002; Yang et al., 2004; Hsiao et al., 2009). AHAs either enhance or diminish skin photodamage depending mainly on the concentration used for topical application and the duration of exposure. We previously demonstrated the phototoxic synergistic effects of GA (5 mM) and UVB (50 mJ/cm2) in HaCaT cells. (Lai et al., 2011). In addition, we found that GA at a low dose (0.1 mM) either alone or with UVB irradiation reduces the expression of inflammasome genes (such as NLRC4 and ASC) through epigenetic modification by increasing total DNA methyltransferase (DNMT) activity (Tang et al., 2016). Therefore, we propose that the concentration of GA determines whether it has phototoxic or photoprotective effects on human keratinocytes and that inflammasomes are involved in such processes. However, the anti-inflammatory action of different concentrations of GA in primary normal human epidermal keratinocytes (NHEKs) or mouse skin remains unclear.
Inflammasomes are a crucial component of innate immunity involved in inflammation (Beer et al., 2014; Zhou et al., 2011). An inflammasome complex acts as an intracellular sensor and recruits pro-caspase-1 to the complex (Meylan et al., 2006). Subsequently, caspase-1 is autoproteolytically activated, leading to the conversion of pro-interleukin IL-1β to its biologically active form (Drexler et al., 2012). Repeated UVB irradiation is a complete carcinogen, because it induces reactive oxygen species (ROS) generation and DNA mutations (Feldmeyer et al., 2007; Akhalaya et al., 2014). UVB irradiation induces inflammasome-dependent IL-1β secretion in human skin keratinocytes (Sollberger et al., 2015); changes in IL-1β reflect the UVB-induced inflammatory status of the skin.
In this study, we examined the potential molecular mechanisms underlying the photoprotective and anti-inflammatory effects of GA treatment on UVB irradiation in primary NHEKs and the skin of C57BL/6J mice. We demonstrated that the effects were regulated through anti-inflammatory pathways related to ROS formation, the expression of inflammasome complexes, and the pro-inflammatory cytokine IL-1β. Furthermore, we elucidated on the appropriate doses of GA for phototoxic and photoprotective effects in an animal model.
Methods
Cell culture and drug treatment
Primary NHEKs were obtained from Cell Applications and cultured in a serum-free keratinocyte growth medium (Gibco) containing insulin (10 mg/mL) and human recombinant epidermal growth factor (0.1 ng/mL) in a humidified atmosphere with 5% CO2 at 37°C. The experiments were performed between passages 3 and 5. Cells at 70–80% confluence were stimulated with GA. Aqueous solutions of GA were prepared in water. NHEKs were treated with 0.1 mM (pH 7.4) or 5 mM (pH 7.1) for 24 h followed by UVB exposure.
In vivo experiments
C57BL/6J mice were purchased from the National Laboratory Animal Center and housed in the Laboratory Animal Center, Tzu-Chi University, until they were 8–9 weeks old. The mice were maintained in clean rooms with a controlled temperature and humidity and 12-h light/12-h darkness cycles. The mice were anesthetized through an intraperitoneal injection of ketamine:xylazine (80 and 10 mg/kg body weight, respectively); the volume is 0.1 mL/each animal mouse before being subjected to procedures. Nine groups (n = 4) of mice were used. The hair on the backs of the mice was removed using commercially available hair removal creams containing thioglycolate trihydrate (approximately 250 μL/mouse) 2–3 days before the experiments. We determined the effects of the topical application of GA solutions (1%, 1.25%, 1.5%, 1.75%, 2%, 2.25%, 2.5%, 2.75%, and 3%). Aqueous solutions of GA (Sigma Chemical Co.) were prepared; the back of each animal was marked with Indian ink to divide it into five squares (each 0.6 × 0.6 cm2), four of which were treated with 10 μL of each concentration of peeling agents, while one square was left untreated. The mice were pretreated by applying GA once a day on day 1, after which they were exposed to UVB (50 and 100 mJ/cm2) on day 2. After 24 h, they were sacrificed on day 3. They were irradiated using KLBiotech STS-1 sunlamps (KLBiotech). UVB is 290–315 nm although a boundary of 320 nm is also used, and the irradiance of UVB was 1.5 mW/cm2, determined using a Dermaray UV Meter and Detector (Gigahertz-Optik, Pochheim). Exposure of the skin of the back of C57BL/6J mice to UVB (320 nm peak) irradiation at doses of 60–75 mJ/cm2 and 24–48 h later results in an erythema. Our animal research methods were approved by the Animal Care, and they comply with the commonly accepted three R's of Tzu-Chi University (Approval ID 103-29-1). Samples of mouse skin were obtained, immediately immersed in buffered 10% formalin, and processed for immunohistochemical (IHC) staining as previously described (Feldmeyer et al., 2007; Tang et al., 2013). Serial sections of 4-μm thickness were mounted on glass slides and stained using hematoxylin–eosin staining, according to previously reported methods (Kim et al., 1998; Wu et al., 2015a). The viable epidermal thickness, excluding the stratum corneum, of the stained sections was histometrically measured using digital photographs. The epidermal thickness was measured at a minimum of four points (a, b, c, and d) on multiple sections per treatment site.
UVB irradiation in vitro experiments
After pretreatment with GA for 24 h, the growth medium of the NHEK monolayer was replaced with phosphate-buffered saline (PBS, pH 7.3) 2 mL. Cells were irradiated using UVB (50 mJ/cm2) radiation in the presence of PBS and without a plastic lid.
Viability assay with MTS
The MTS assay was performed using CellTiter 96 AQueous One Solution (MTS solution) according to the manufacturer's instructions (Promega). Diluted MTS solution 1:20 (v/v) was added to the NHEK cells with a complete change of the culture medium at 37°C for 2 h. The absorbance of the formazan products at 490 nm was measured using a Multiskan spectrum microplate spectrophotometer (Thermo Labsystems). The cells incubated with control medium were considered 100% viable.
Real-time polymerase chain reaction
Total RNA derived from cells was isolated using the REzolTM C&T method (Protech Technology). The derived total RNA was reverse-transcribed using the GoScript™ Reverse Transcriptase Kit (Promega), and real-time PCR was performed using a Roche LightCycler 480 system and the IQ2 SYBR Green Fast qPCR Mix (Biogenesis), according to the manufacturer's protocol. The primers used for qPCR analysis are listed in Table 1.
Western blot assay
Cells were extracted with RIPA buffer (Millipore Corp.) to yield cell lysates. To determine the protein expression of components of inflammasome complex (NLRC4, NLRP3, ASC, and AIM2) and caspase-1, cell lysates were resolved using 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis, followed by Western blot analysis with appropriate primary antibodies overnight at 4°C. Blots were washed with tris-buffered saline and Tween 20 for 1 h. Monoclonal Mouse anti-CIAS/NLRP3 (Abcam), rabbit anti-caspase-1α (p20) antibody, rabbit anti-NLRC4 antibody, rabbit anti-ASC1 antibody, rabbit anti-IL1-β (Cell Signaling Technology), mouse anti-AIM2, and mouse anti-β-actin monoclonal antibody (Sigma-Aldrich) were used as primary antibodies. HRP-conjugated rabbit anti-goat IgG antibody (GeneTex), HRP-linked anti-mouse, and anti-rabbit IgG antibody (Cell Signaling) were used as secondary antibodies. The immune reactions were visualized by an ECL and UVP BioSpectrum Imaging System (Upland). The immunoreactive bands were quantified using ImageJ software.
ELISA and cytokine array
To determine the activity of inflammasomes in the NHEK cells, we used UVB irradiation, which is known to induce IL-1β and IL-18 secretion. We also measured other cytokines (including IL-6 and tumor necrosis factor [TNF]-α). All of the secreted cytokine levels were determined using commercial ELISA kits (R&D Systems). Culture supernatants were assayed according to the manufacturer's protocol. The standard curve revealed a direct relationship between the optical density and secreted cytokine levels. Colorimetric analysis was performed using a plate reader (MRX II, DYNEX Technologies) at 450 nm. Data were expressed as mean ± SD pg/mL of retrieved culture supernatant. The human cytokine array Panel A (R&D Systems) was used to evaluate cytokine secretion profiles in the supernatants of the NHEK cells, according to the manufacturer's instructions.
ROS assay
Changes in the levels of ROS in the NHEK cells were detected using flow cytometry. The NHEK cells were treated with GA (0.1 and 5 mM) and then exposed to UVB. The cells were harvested and washed twice, resuspended in 10 mM 2,7-dichlorodihydrofluorescein diacetate (Sigma-Aldrich), and incubated at 37°C for 30 min. The ROS levels were analyzed through flow cytometry (Becton Dickinson). We calculated the percentage of ROS using CellQuest software (Becton Dickinson).
DNA damage detection by immunofluorescence γ-H2AX expression and comet assay
The NHEK cells were washed twice in PBS and fixed in 95% ethanol containing 5% acetic acid for 5 min. The cells were then suspended in 0.2% Triton X-100 (Sigma-Aldrich) in a 1% (w/v) solution of bovine serum albumin (BSA; Sigma-Aldrich) in PBS for 30 min. After washing the cells with PBS, we incubated them overnight with 1% BSA containing 1:100 diluted anti-phospho-histone H2AX (Ser 139) antibodies (Sigma-Aldrich) at 4°C, followed by incubating them with FITC-conjugated secondary antibody for 1.5 h at room temperature. The nuclei were counterstained with 4,6-diamidino-2-phenylindole (DAPI; Sigma-Aldrich). The cells were mounted on microscope slides and observed using a fluorescence microscope. The experiments were repeated thrice, and independent results for each sample (clonogenic fraction and number of cells lacking foci) were plotted. For detecting DNA single-strand breaks and alkali-labile sites, we used the comet assay, as previously described (Tang et al., 2010; Osipov et al., 2014; Won et al., 2014). Recorded images were analyzed using CometScore software available as freeware (TriTek.).
Statistical analysis
All measurements were performed in triplicate, and data of at least three independent experiments were statistically evaluated. Results were expressed as the mean ± SD. Data are expressed as mean ± SD. Mean was compared using nonparametric test, and p-value less than 0.05 was considered statistically significant. SPSS 18.0 software (SPSS, Inc.) was used to perform statistical analysis.
Results
GA reduced UVB-induced death in NHEKs
To assess the effects of GA on cell viability following UVB irradiation, NHEK cells were treated with 0.1 mM GA for 24 h, followed by exposure to 50 mJ/cm2 of UVB. Cell viability was measured using the MTS assay. The measured viability of the UVB-irradiated cells significantly decreased compared with control group (p < 0.01). A large proportion of cells were necrotic, and some exhibited shrinkage and indistinct nuclei, suggesting apoptosis (arrowheads). However, the group treated with GA before UVB irradiation exhibited no significant change in cell morphology, compared with the control (Fig. 1). NHEKs that were treated with GA before exposure to UVB were significantly protected from UVB-induced loss of cell viability and cell death. We thus used GA treatment at 0.1 mM and UVB irradiation at 50 mJ/cm2 in the subsequent experiments.
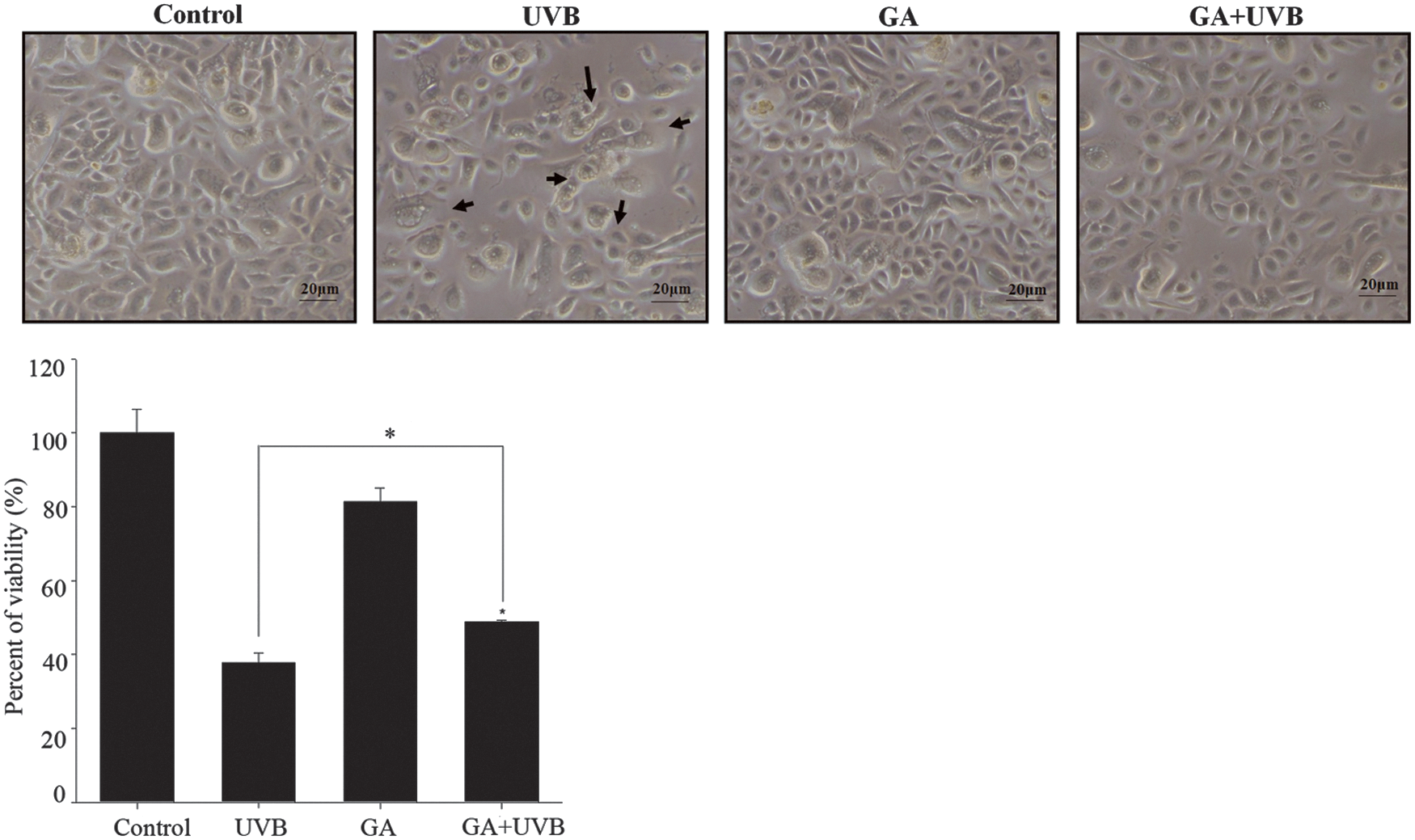
NHEK cell morphologies following treatment with GA and UVB irradiation. NHEK cells pretreated with GA (0.1 mM) with or without UVB irradiation (50 mJ/cm2) were observed (phase-contrast microscopy). Cells were necrotic; shrinkage of cells with indistinct nuclei, suggesting apoptosis (arrowheads). Cell culture morphologies (bar = 20 μm). Cell viability was evaluated using the MTS assay. *Significantly different between groups compared (p < 0.01). GA, glycolic acid; NHEK, normal human epidermal keratinocyte.
GA reduced ROS levels in UVB-irradiated NHEKs
ROS formation increases with UVB irradiation. To determine whether GA pretreatment impairs ROS responses in NHEK cells of UVB-induced inflammation, the cells were UVB irradiated at 50 mJ/cm2 (blank bar, p < 0.01). GA (0.1 and 5 mM) significantly reduced the ROS levels in the NHEK cells after UVB irradiation (50 mJ/cm2) (p < 0.01) (Fig. 2), thereby reducing photodamage in the NHEK cells.
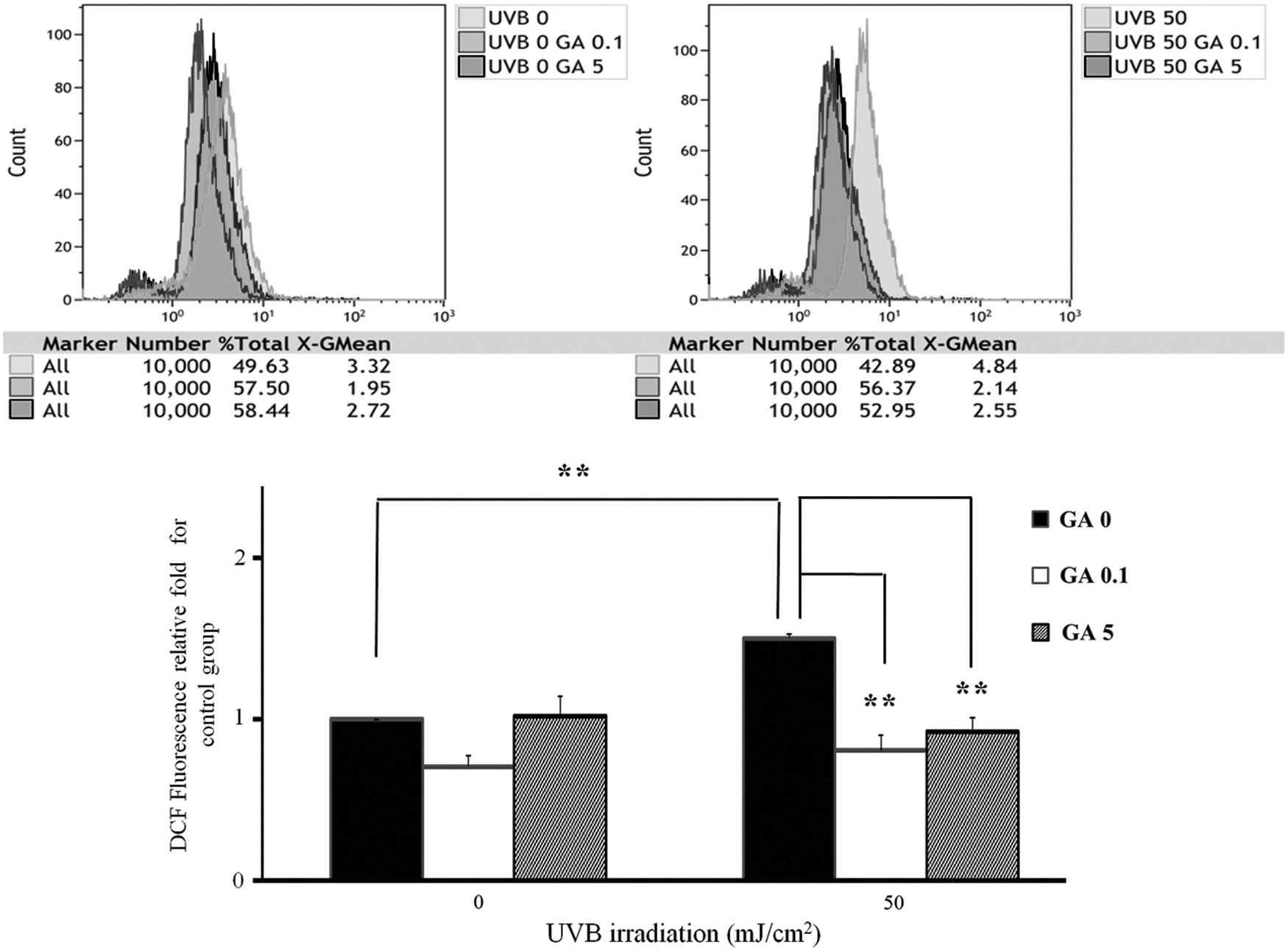
Effect of GA (0.1 and 5 mM) and UVB irradiation (50 mJ/cm2) on ROS of NHEK cells. The intracellular ROS levels were measured through fluorescence microscopy using the oxidant sensitive probe H2DCFDA. DCF fluorescence in the cells was quantified with a flow cytometry. The results represent the mean ± SD; n = 3. **p < 0.01. Values obtained from three separate experiments. ROS, reactive oxygen species.
GA reduced levels of γ-H2AX expression and DNA fragments in NHEK cells treated with UVB irradiation
Immunostaining of DNA fragments and phosphorylated H2AX histone protein, indicating single- and double-strand DNA breaks, was performed. As a predictive test for micronucleus formation, we examined whether GA pretreatment can also promote cell recovery from UVB-induced DNA breakage using a γ-H2AX assay and comet assay. The NHEK cells were irradiated with UVB, and immunofluorescence staining was performed directly thereafter. The UVB-irradiated cells showed greater γ-H2AX fluorescence than nonirradiated cells. GA (0.1 mM) significantly reduced DNA damage, indicated by γ- H2AX fluorescence, in the UVB-irradiated NHEK cells (Fig. 3A). A measurement of the γ- H2AX foci number indicated that GA (0.1 mM) reduced UVB-induced foci numbers (Fig. 3B). We also observed a reduction in the intensity of comet tails with GA treatment in the UVB-treated NHEK cells (Fig. 3C, D). All of these data suggested that GA (0.1 mM) showed a photoprotective effect by reducing the DNA damage markers, namely γ-H2AX expressions and DNA break tail. Therefore, GA (0.1 mM) has a protective effect against DNA damage in UVB-treated NHEK cells (Fig. 3).
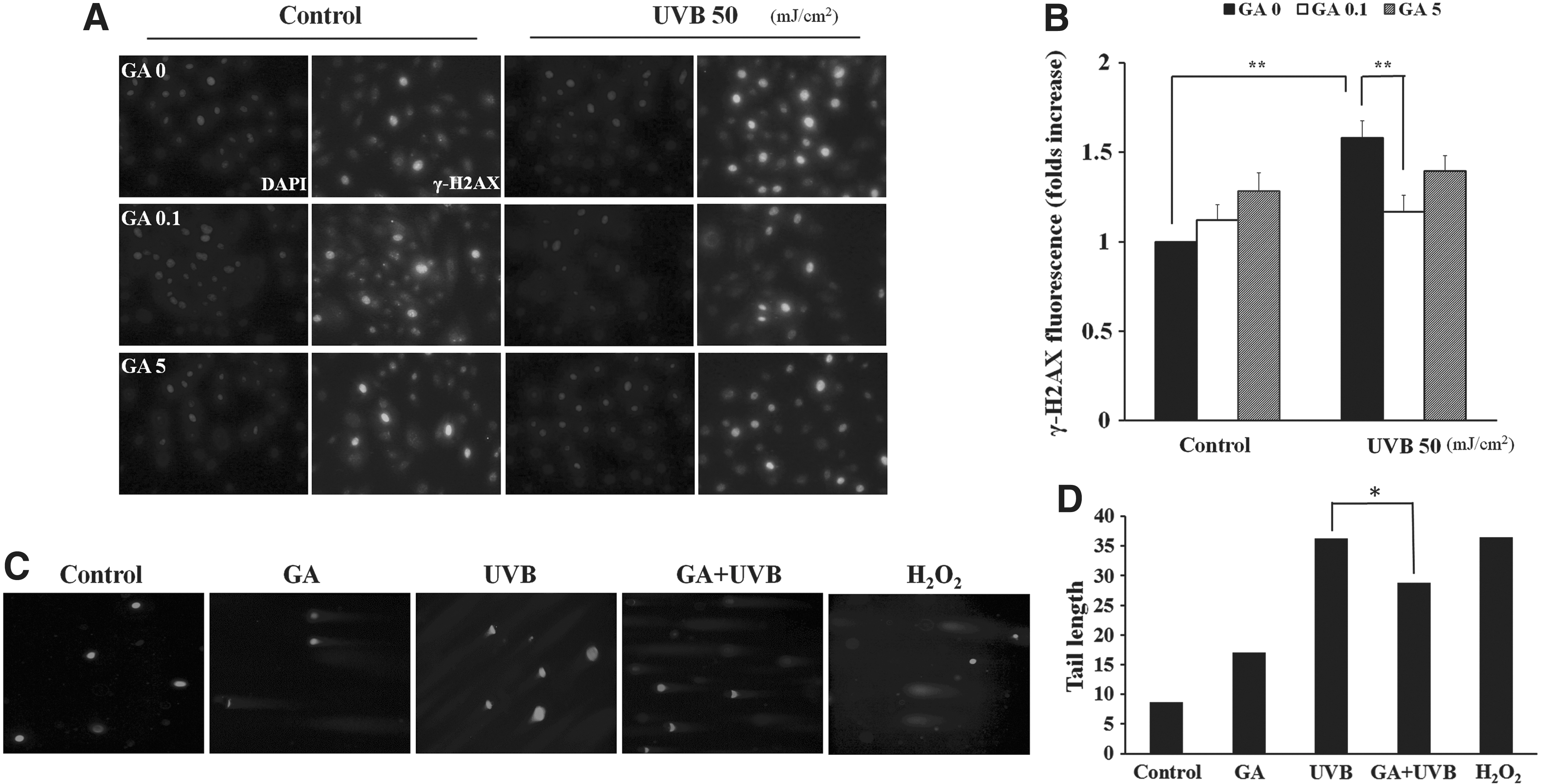
Immunostaining for γ-H2AX and DNA tail in NHEK cells exposed to UVB (50 mJ/cm2) irradiation.
Effects of GA on the expression of inflammasome complexes and caspase-1 in NHEKs
According to the identity of the sensor mRNA involved, the following four types of inflammasomes have been described: NLRP3, NLRC4, ASC, and AIM2. Q-PCR examination demonstrated that UVB irradiation alone in the NHEK cells increased the expressions of the inflammasome complexes. Treatment with GA (0.1 mM) before UVB exposure significantly reduced the expression of NLRC4 and AIM2 in the NHEK cells (Fig. 4A). Western blot analysis showed a reduction in the levels of the proteins NLRC4 and AIM2 in the cells treated with GA before UVB irradiation (Fig. 4B). We also elucidate that GA reduced the gene and protein expressions of caspase-1 (Fig. 4C). These data show that GA is potentially photoprotective due to GA reducing the expression of the inflammasome complexes NLRC4 and AIM2, as well as caspase-1, thereby diminishing inflammatory reactions.

GA abolished the UVB-mediated inflammasome complex (NLRC4, ASC, and caspase-1) expression in NHEK cells.
Inhibition of UVB-induced production of IL-1β by GA in NHEKs
To determine whether UVB irradiation of NHEK cells induces the secretion of inflammatory cytokines, cytokine array and ELISA were conducted to compare the levels of inflammatory cytokines in the NHEK cells with or without GA pretreatment. Thirty-five cytokines were detected on an inflammation panel array. The levels of the cytokines CXCL1, CXCL2, IL-1α, IL-1β, IL-8, and DER6 significantly decreased in the GA-pretreated UVB-irradiated cells, compared with the untreated UVB-irradiated cells (Fig. 5A). We also detected other inflammation cytokines using ELISA kits. The results revealed that IL-1β, IL-18, and TNF-α levels in culture media significantly increased after UVB irradiation. GA (0.1 mM) was found to reduce IL-1β and TNF-α levels in UVB-irradiated keratinocytes (p < 0.05) (Fig. 5B).

The concentrations of the inflammation cytokines secreted from the GA pretreated with or without UVB irradiation in NHEKs. Inflammatory cytokines in culture supernatant were examined by cytokine array and ELISA kit.
Effects of GA on epidermal thickness and inflammasome complex proteins in C57BL/6J mice
Epidermal thickness was used as a measure of epidermal and dermal vascular permeability, as well as edema, which are the hallmarks of a cutaneous inflammatory response. GA (1.25–3%) was topically applied on hairless mice before UVB irradiation. UVB (100 mJ/cm2) irradiation significantly damaged and injured the skin tissue, including the epidermis and dermis of the mice. Topical skin treatment with GA concentrations higher than 2.5% resulted in a significant removal of stratum corneum, increased epidermal thickness, and discernible inflammatory cell (neutrophil) infiltration (white arrows). In contrast, at concentrations less than 2.0%, GA did not cause any change in epidermal thickness and inflammatory response (Fig. 6A). Among the inflammasome proteins, the increase in NLRC4 protein expression (black arrow) depended on UVB irradiation dose. Topical treatment with 3% of GA also increased the NLRC4 protein expression in IHC, but treatment with a low dose of GA (less than 2%) followed by UVB irradiation significantly reduced the NLRC4 protein expression (Fig. 6B). We also observed a reduction in the mRNA levels of NLRC4 with GA pretreatment in the UVB-irradiated mice dorsal skin cells (Fig. 6C). All of these data indicated that the topical application of GA at optimal concentrations (1.5% or 2%) on hairless mouse skin resulted in an anti-inflammatory effect.
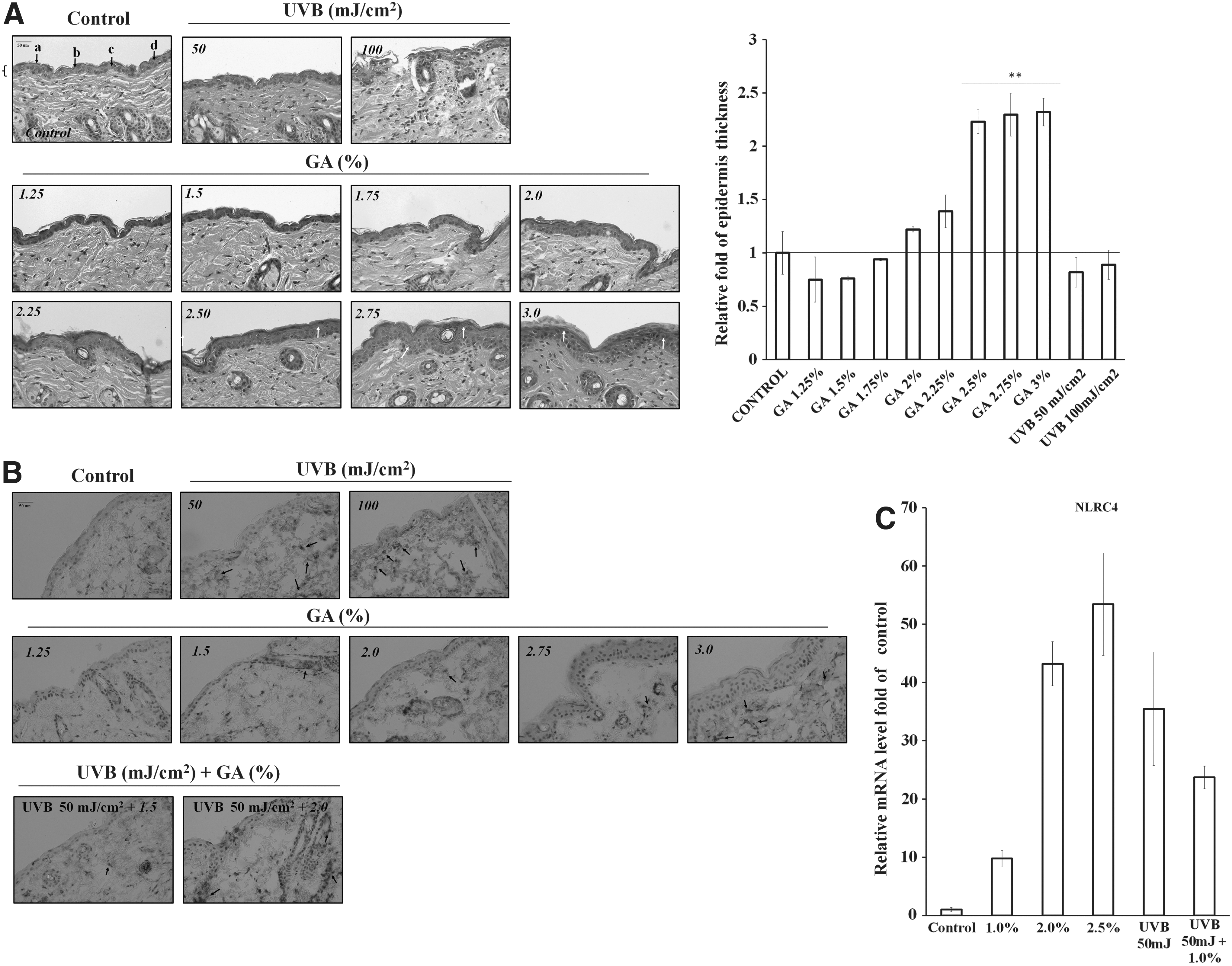
Effect of topical application of GA on UVB-induced skin damage in mice.
Discussion
UVB irradiation causes inflammation and DNA damage, and a detailed analysis of how the inflammatory genes are affected in response to GA is crucial for investigating the anti-inflammatory effect on NHEKs. In this study, we investigated the protective effect of GA treatment on NHEK cells before UVB radiation, determining a significant reduction of UVB-induced ROS formation and accumulation; single- and double-strand DNA break markers (γ-H2AX and DNA tailing); expression of genes and proteins of the inflammasome complex (NLRC4 and AIM2); caspase-1 expression; and pro-inflammatory cytokine IL-1β secretion in the NHEK cells. We also elucidate that the topical application of GA at optimal concentrations (1.5% or 2%) on hairless mouse skin resulted in an anti-inflammatory effect. Our data demonstrate the molecular mechanisms through which the optimal dose of GA (0.1 mM) exerts its anti-inflammatory and photoprotective activities on UVB-irradiated NHEKs.
IL-1β mediates acute inflammatory responses and provides a link between innate and adaptive immunity (Watanabe et al., 2007). IL-1β secretion follows two sequential events as follows: (i) upregulation of pro-IL-1β through NF-kB-mediated transcriptional activation (Martinon and Tschopp, 2005) and (ii) inflammasome/caspase-1-mediated processing (Keller et al., 2008). Recent studies have shown that the maturation of IL-1β is inflammasome dependent on UVB irradiation (Drexler et al., 2012; Sollberger et al., 2015). In our data, the UVB-irradiated NHEK cells pretreated with GA showed significantly inhibited maturation and secretion of IL-1β (Fig. 5B). Caspase-1 is a major UVB-dependent activator of IL-1β in epidermal keratinocytes (Feldmeyer et al., 2007; Hiramoto et al., 2012; Hattinger and Wolf, 2015; Sollberger et al., 2015); this is consistent with our finding that GA reduced the caspase-1 mRNA and protein expression in the UVB-irradiated NHEK cells (Fig. 4C). GA significantly reduced NLRC4 and AIM2 expressions in UVB-treated NHEKs (Fig. 4), indicating that GA inhibits UVB-induced oxidative damage and inflammation through inflammasome/caspase-1-mediated signaling pathways in NHEKs.
The conditioned medium was analyzed using an inflammation-related cytokine array to detect other secreted proteins from the GA-treated NHEK cells with or without UVB irradiation (Fig. 5A). As a result, increased expression levels of IL-1(α, β) and other chemokines (including CXCL1, IL-8, and DER6) were significantly reduced in the GA-pretreated UVB-irradiated cells. We also examined the levels of other pro-inflammatory cytokines, such as IL-6, IL-18, and TNF-α, through ELISA kits (Fig. 5B). TNF-α significantly decreased in the GA-pretreated UVB-irradiated group. Therefore, we considered that GA may also be involved in other cellular regulated pathways such as COX-2, p38, and JNK MAPK signal pathways (Zhai et al., 2015); PKC associated pathway (Grandjean-Laquerriere et al., 2005); or Nrf2/HO-1 inflammation pathway (Wu et al., 2015b). Previous studies have indicated that UVB activated autophagy in JB6 epidermal cells and autophagy appeared to be a protective response to UVB-induced damage (Yang et al., 2012; Strozyk and Kulms, 2013). In this study, we demonstrate that GA had anti-inflammatory and photoprotective effects against UVB irradiation; however, the autophagy responsible for the beneficial effects on GA has not been thoroughly investigated in skin cells. Different concentrations of GA produced either phototoxic or photoprotective effects in skin cells. Our previous study demonstrated a phototoxic synergistic increase of IL-1β in HaCaT cells treated with UVB and GA (5 mM) (Lai et al., 2011). However, we did not find a synergistic increase of IL-1β in similarly treated NHEKs in the current study. Although HaCaT and NHEKs are both human keratinocytes, NHEK has a more sensitive response in expression of IL-1β and provides a more appropriate cell model for investigating the effects and photoprotective molecular mechanisms of GA in human skin keratinocytes.
GA has been established as a nontoxic fruit substance. Previous studies have attributed the photoprotective and anti-inflammatory effects of topically applied GA (8%, pH 3.25) to a transient enediol-like structure and hypothesized that the resonating double bonds exert the antioxidant activity of ascorbic acid (Perricone and DiNardo, 1996). However, topical GA (10%) has been shown to increase the sensitivity of human skin in vivo to solar simulated radiation in other studies (Tsai et al., 2000; Kaidbey et al., 2003; Kornhauser et al., 2009). In our previous studies, we have found that the exacerbation or attenuation of inflammatory reaction in human keratinocytes by GA depended on its dose. AHAs at higher concentrations (e.g., GA at 5 mM or lactic acid at 17.5 mM) induced caspase-3 activation and resulted in increased apoptosis in the HaCaT cell line (Hsiao et al., 2009; Lai et al., 2011). Recently, we found that GA silences the inflammasome complex genes NLRC4 and ASC by inducing DNA methylation in HaCaT cells (Tang et al., 2016), which indicates that GA at 0.1 mM activated DNA methyl-transferase (DNMT), particularly DNMT-3b, and is involved in the epigenetic methylation mechanism. A lower dose of GA (e.g., 0.1 mM) reduced ROS formation, levels of DNA damage markers of γ-H2AX expressions and DNA fragments, and secretion of the inflammatory mediator IL-1β in this study. We thus conclude that GA at an optimal low dose (e.g., 0.1 mM) reduces ROS generation following UVB irradiation. Recently, we demonstrated that low dose of GA on UVB-treated NHEKs resulted in decrease of apoptosis. The photoprotection effect of GA was supported by suppressing expressions of p-Akt, p-ERK1/2, p-p-38 MAPK, p-c-Jun, and NF-κB (p65), indicating that GA inhibited PI3K/Akt and MAPK signaling pathways (data not show). In this study, we provided more evidence regarding the role of GA in the regulation of inflammasome complex activation. GA exerts either anti-inflammatory and photoprotective effects or harmful effects on UV-irradiated human keratinocytes, depending on its concentration (Ditre et al., 1996; Park et al., 2002; Monheit, 2004; Wheeland and Dhawan, 2011). The potential of pharmacokinetics and bioavailability of GA in clinical trials depends on its concentration in the adjuvant (cream or lotion). The concentrations of GA used in our study were 0.1 and 5 mM, which are approximately 0.00094% and 0.047% (in w/v). These concentrations of GA are remarkably lower than those used in cosmetic (less than 5%) or clinical-peeling formulations (10–70%). Topical application of 3% GA, the threshold concentration, resulted in epidermal thickness, stratum corneum loss, and dermal inflammatory cell infiltration in the mouse
Conclusions
We summarized our findings as shown in Figure 7. We demonstrate that GA (0.1 mM) exerts anti-inflammatory and photoprotective effects through the suppression of ROS formation and reduction of DNA damage, γ-H2AX expressions, and DNA break tail. We also demonstrate that topical application of GA (1.5–2%) can protect mouse skin from UVB-induced inflammation (e.g., inhibition of NLRC4 expression). Taken together, GA (0.1 mM) exhibits a protective effect against DNA damage in UVB-treated NHEKs through the attenuation of DNA damage, reduction of the expression of inflammasome genes NLRC4 and AIM2, and reduction of caspase-1 and IL-1β secretion in UVB-irradiated NHEKs.
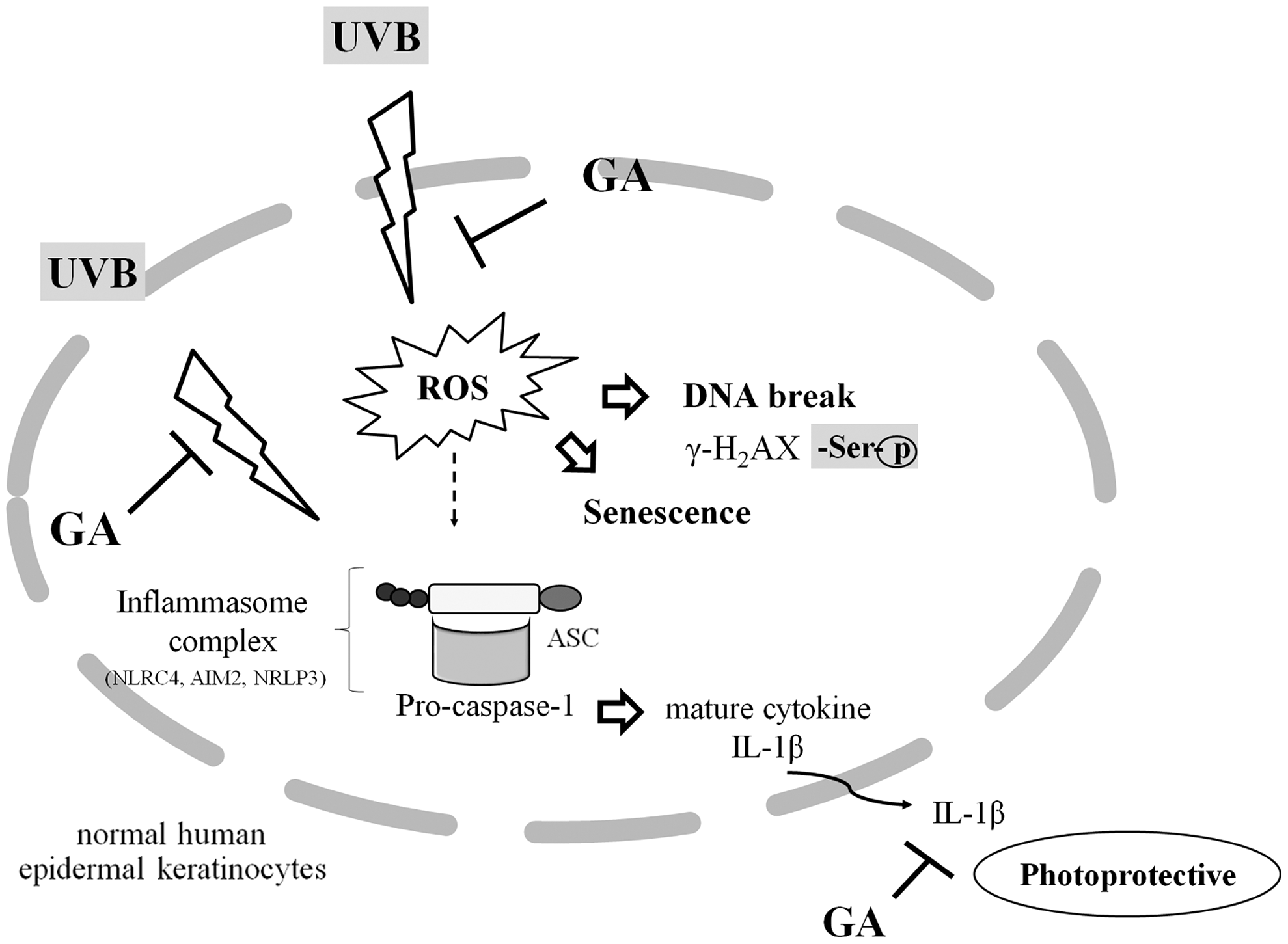
Graphical Summary. The schema proposes possible anti-inflammatory and photoprotective mechanisms of GA to reduce ROS levels and DNA damage in NHEK cells. GA had anti-inflammatory and photoprotective effects against UVB irradiation through the inflammasome complex (NLRC4 and ASC) and subsequently reduce the expressions of caspase-1 and pro-inflammatory cytokine IL-1β in NHEK cells.
Footnotes
Acknowledgments
The authors thank Dr. Fu-Tong Liu at Academia Sinica Institute of Biomedical Science, Taiwan, for providing constructive discussion and recommendations. The authors thank Miss Ko-Chen Chen. This study was supported by the Buddhist Tzu Chi Hospital, Hualien, R.O.C. (TCRD103-03 and TCRD104-44).
Disclosure Statement
No competing financial interests exist.