
Abstract
Exosomes are nano-sized biological membrane-enclosed vesicles that contain a cell-specific cargo of proteins, lipids, and nucleic acids that are released and taken up by most cell types, thereby inducing expression and functional changes via horizontal transfer of cargos between cells. Thus, exosomes present a largely unknown “cell-to-cell” communication system, which is now increasingly being investigated for diagnostic and therapeutic use in cardiovascular disease (CVD). The purpose of this review is to summarize recent findings on the properties and roles of exosomes in a variety of physiological and pathological settings related to CVD. We focus on available information on exosome-mediated intercellular communication relevant to myocardial injury, repair, and regeneration. Finally, we address the promise of exosomes as valuable diagnostic and prognostic biomarkers, and their potential use as therapeutic tools in CVD. Exosomes remain largely unexplored for therapeutic use in the field of cardiovascular diagnosis and medicine. A more detailed characterization of cardiac exosomes shed by different components of the heart will be of fundamental importance to address specific changes in the profile of exosomal microRNAs and proteins, which will enable the clinical use of exosomes as minimally invasive diagnostic tools and vehicles for delivery of targeted therapies for CVD.
Introduction
C
Exosomes are lipid bilayer vesicles of endosomal origin that range from 40 to 100 nm in diameter, which arise on fusion of multivesicular bodies (MVBs) with the plasma membrane (Huber and Holvoet, 2015). On fusion with the plasma membrane, MVBs release their contents into the extracellular space. These released vesicles are termed exosomes (Emanueli et al., 2015). When first identified in mature sheep reticulocytes in the late 1980s, exosomes were initially ignored as cell debris derivatives with no significant impact on neighboring cells (Johnstone et al., 1987). However, recent evidence indicates that these extracellular vesicles are not only for “rubbish collection” but also functional vehicles that carry a complex cargo of proteins (Choi et al., 2015; Abramowicz et al., 2016), lipids (Record et al., 2014), and nucleic aids (Gezer et al., 2014; Ahadi et al., 2016; Ohno and Kuroda, 2016) that are capable of delivering these cargos to the target cells they encounter, which may ultimately alter their function and behavior over long distances. Thus, exosomes represent a novel mode of intercellular communication, which may play a major role in many cellular processes and pathways, such as immune responses, intercellular communication, and antigen presentation (Thery et al., 2009; Huang-Doran et al., 2016).
Since exosomes were first isolated from cultures of cardiomyocytes grown in vitro in 2007 (Gupta and Knowlton, 2007), the potential of exosomes as diagnostic biomarkers or therapeutic agents for CVD has attracted significant attention. Several lines of evidence confirmed that exosomes are involved in mediating intercellular communication between both proximal and distant cells under not only physiological conditions, such as cardiac growth, development, and angiogenesis, but also diverse pathological states, including atherosclerosis, IR injury, and cardiac remodeling. In this review, we update recent progress made in exosome research and highlight the roles of exosomes in cardiovascular pathology.
Exosomes in CVD
Exosomes in atherosclerosis
Exosomes derived from different cell types exhibit pleiotropic biological activities, either protective or detrimental, which are primarily dependent on the cellular origin and current status (Keerthikumar et al., 2016). It has been shown that exosome-mediated cellular communication plays an important role in each phase of atherosclerosis. Generally, atherosclerosis development is initiated by endothelial dysfunction, which is primarily due to local disturbances in blood flow along the endothelium. The transcription factor Krüppel-like factor 2 (KLF2) is a critical regulator of endothelial gene expression patterns that are activated by atheroprotective flow in endothelial cells (ECs) (Sun and Feinberg, 2015). Hergenreider et al. (2012) demonstrated that KLF2-expressing human umbilical vein ECs can prevent de-differentiation of smooth muscle cells and, consequently, reduce atherosclerotic lesion formation in an exosome-dependent manner through exosome-mediated transfer of atheroprotective miR-143/145, providing a promising strategy to combat atherosclerosis. Meanwhile, endothelial dysfunction can also be activated by elevation in serum levels of oxidized LDL (ox-LDL) and homocysteine (Hcy), the most important risk factors for atherosclerosis and cardiovascular morbidity (Tsuda, 2015). It has been shown that ox-LDL and Hcy can induce the release of HSP70-containing exosomes from cultured aortic ECs, which selectively activate monocytes via proinflammatory differentiation to macrophages in a toll-like receptor 4 (TLR4)-dependent manner, resulting in adhesion of monocytes to ECs so as to play a pro-inflammatory and proatherogenic role in the early steps of atherosclerosis (Zhan et al., 2009).
Impairment of the endothelium is mostly accompanied with increased endothelial permeability to lipoproteins, macrophage recruitment, foam cell formation, and homing of T lymphocytes and platelets. During these processes, diminished scavenger receptors, such as the type II scavenger receptor CD36, seem to play a crucial role in the protective effects of exosomes in atherosclerosis. Evidence has confirmed that activated platelet-derived exosomes can rapidly reduce CD36, a type II scavenger receptor in platelets, through enhanced ubiquitination and proteasome degradation of CD36, resulting in attenuation of platelet aggregation and adhesion to a collagen matrix ex vivo, and suppression of occlusive thrombosis in an FeCl3-induced arterial injury model (Srikanthan et al., 2014). Meanwhile, CD36 also allows macrophages to bind and internalize ox-LDL to supply lipids to promote foam cell formation (Rahaman et al., 2006; Shantsila et al., 2010). Consequently, platelet exosomes inhibited ox-LDL binding and cholesterol loading into macrophages (Srikanthan et al., 2014). As platelet-derived exosomes can simultaneously interfere with two essential elements of athero-thrombosis, foam cell formation and platelet responsiveness to damaged arteries, they may serve as a potential new therapeutic targets of the disease process. However, exosomes released from activated T lymphocytes enriched with cholesterol and exposing phosphatidylserine (PS) at the outer membrane leaflet can induce cholesterol accumulation in parallel with enhanced production of the proinflammatory cytokine tumor necrosis factor (TNF)-α in human monocytes by exosome internalization via endogenous PS receptors. This underlies that T cell exosomes may promote atherogenesis within the atherosclerotic plaque (Zakharova et al., 2007).
Furthermore, the phenotypic changes that transform vascular smooth muscle cells (VSMCs) from a quiescent contractile state to a synthetic active state also play a role in the pathogenesis of atherosclerosis. In a recent study, Kapustin et al. (2015) demonstrated that proliferative VSMCs can actively release exosomes enriched with calcification inhibitors, such a fetuin-A, and other cargoes to facilitate vascular repair processes, whereas prolonged osteogenic stimuli (e.g., elevated calcium, cytokines, and growth factors) and a mineral imbalance may enhance exosome secretion through upregulation of sphingomyelin phosphodiesterase 3 and shift of VSMCs toward a procalcific state, implying modulation of the exosome release pathway as a novel therapeutic target for vascular calcification, a common complication in patients with atherosclerosis. Atherosclerosis is a chronic inflammation disease with dramatically increased numbers of dendritic cells (DCs), which participate in almost all stages of atherosclerosis and contribute to ECs activation, cytokine and chemokine production, and even fatty steaks formation. Interestingly, a recent study revealed that DCs can release exosomes to crosstalk with ECs, resulting in accelerated endothelial inflammation and progression of atherosclerosis through the exosomal TNF-α-mediated NF-κB pathway, underlying that DC-exos represent a novel mechanism by which DCs promote activation of ECs, as well as progression of atherosclerosis (Gao et al., 2016) (Fig. 1).

The role of exosomes in atherosclerosis. An outline of the known cellular communication pathways that exosomes are known to participate in with regard to the development of atherosclerosis. For a detailed explanation of the pathways, see text on exosomes in atherosclerosis section. DCs, dendritic cells; ECs, endothelial cells; ox-LDL, oxidized LDL; miR, microRNA; PS, phosphatidylserine; VSMCs, vascular smooth muscle cells.
Exosome in myocardial ischemic injury
After atherosclerotic plaque formation via the gradual buildup of cholesterol and fibrous tissue in the arterial wall, the lumen becomes narrowed after decades of blood stream irregularities. Plaques can become unstable, rupture, and additionally promote blood clot formation, which results in occlusion of the artery and eventual myocardial ischemic injury. A rupture of an atherosclerotic plaque, severe enough to completely block a coronary artery, may lead to acute myocardial infarction (AMI) and subsequent necrosis of the downstream myocardium. Although subject to debate, a recent study established that the majority of miRs in the blood plasma are enclosed in exosomes. The remarkable alterations in miRs in circulation have been reported to accurately reflect myocardial injury in vivo, thereby laying a foundation for the idea that exosomal miR-mediated communication plays a significant role in the occurrence, development, and even cardiac repair in AMI.
In infarcted heart tissue, elevated serum levels of miR-1 and miR-133a can be detected early after the onset of chest pain without any elevation of serum creatine phosphokinase or cardiac troponin T. These elevated miR-133a and miR-1 have been shown to localize in exosomes primarily originating from the injured myocardium and can be transferred to recipient cells (Kuwabara et al., 2011). The cardioprotective effects of miR-133a and miR-1 by inhibiting heart hypertrophy and their capture by adjacent surviving myocardium via exosomes may be utilized to protect cardiomyocytes from hypertrophy, indicating that circulating miRs released from injured myocardium after AMI can be transferred through the circulation to distant organs by exosomes and functionally impact the biological activities of recipient cells. In support of this concept, Cheng et al. (2012) detected higher levels of heart-enriched-specific miR-1 and miR-208 in urine exosomes isolated from rats after AMI. Injection of PKH67-labeled exosomes into the circulating blood has been found to enter the kidney tissues and cells, which suggests the capacity of exosome released from the heart to enter the urine by transrenal release. Therefore, urine exosomal miRs, such as miR-1, present a novel group of potential urine biomarkers for AMI with possible high sensitivity and specificity (Cheng et al., 2012). Surprisingly, another recent report indicated that plasma exosomes isolated from AMI patients are also enriched with markedly elevated miR-1, which was found to suppress CXCR4 expression in bone marrow (BM)-derived mononuclear cells. This may contribute to the increased mobilization of BM cells elicited by ischemic injury, suggesting exosome-mediated functional crosstalk between the ischemic heart and BM cells (Cheng et al., 2014). Furthermore, significant elevation of circulating miR-214 has been detected in multiple disease models of hypertrophy and heart failure, which exerts a cardioprotective effect against IR injury by rescuing the heart from further cell death and loss of contractility through repression of the mRNA encoding sodium/calcium exchanger 1, a key regulator of Ca2+ influx, and downstream effectors of Ca2+ signaling that mediate cell death (Aurora et al., 2012). miR-214 is also known to be released via exosomes from ECs (van Balkom et al., 2013), suggesting an additional therapeutic benefit of miR-214-containing exosomes for attenuation of IR injury. Recently, Yang et al. (2016) observed high enrichment of miR-30 in exosomes isolated from serum of AMI patients or culture medium of hypoxic cardiomyocytes, which are capable of regulating autophagy through miR-30a-mediated downregulation of the core autophagy regulators beclin-1, Atg12, and LC3II/LC3I after hypoxia, indicating a promising strategy to treat ischemic heart disease.
Remarkably, several reports suggest a pivotal role of circulating miR-mediated intercellular communication via exosomes in remote ischemic preconditioning (RIPC)-induced myocardial protection, an applicable therapeutic strategy to effectively protect the heart against IR injury. Giricz et al. (2014) first confirmed that exosomes released from the heart after preconditioning are necessary for cardioprotection by RIPC, indicating the importance of vesicular transfer mechanisms in remote conditioning signals. Using a Langendorff isolated heart model of global IR injury, Li et al. (2014) showed that RIPC increased miR-144 precursor levels in circulating exosomes, resulting in increased serum concentrations in mice and humans, and subsequent increases in phospho-Akt, phospho-Glycogen Synthase Kinase 3 Beta, and P-p44/42 mitogen-activated protein kinase (MAPK), and a decrease in phospho-mammalian target of rapamycin, and upregulation of autophagy signaling. Apparently, the endogenous release of miR-144 by exosomes plays a pivotal role in RIPC-induced cardioprotection by improving functional recovery and reducing infarct size, and thus can be considered a biomarker of the effectiveness of RIPC-induced limb ischemia and a potential therapeutic target to reduce clinical IR injury. Furthermore, Yamaguchi et al. (2015) demonstrated an increase in recruitment of miR-29a and insulin-like growth factor 1 receptor (IGF-1R) into exosomes by RIPC treatment. In addition to the known protective roles of miR-29 and IGF-1R in fibrosis and the prevention of cardiac remodeling, there were also found high expression levels of both molecules in the remote noninfarcted myocardium of the RIC group, indicating that transportation to the noninfarcted myocardium via plasma exosomes may have contributed to reduced LV remodeling and oxidative stress, further identifying beneficial therapeutic effects of exosome-mediated intercellular communication in repeated RIPC on cardiac dysfunction after AMI (Yamaguchi et al., 2015). Recently, Vicencio et al. (2015) reported that plasma exosomes of rats and human can protect the myocardium from IR injury via HSP70 on exosomes, which, in turn, activates a pathway downstream of TLR4 that involves extracellular signal-regulated protein kinases 1 and 2 and p38 MAPK, leading to phosphorylation of HSP27, indicating that the HSP70/TLR4 communication axis is a critical component in exosome-mediated cardioprotection.
Exosomes in heart failure
Although continued advancements in AMI therapy have significantly improved survival, the sequelae of IR injury frequently include heart failure. In response, the heart may undergo a series of stresses during the process of blood reconstruction, including certain metabolic and structural adaptations for maintenance of adequate cardiac output. Once these adaptive measures become insufficient, a state of maladaptation may occur, such as hypertrophy (pathological growth of cardiomyocytes and increased wall thickness), fibrosis (loss of contractile myocardium), and aberrant angiogenesis (growth of new vessels) (Waldenstrom and Ronquist, 2014). Interestingly, recent studies revealed that these adaptions are orchestrated by chemokines, growth factors, and miRs, most of which are actively transported by exosomes.
Pathological cardiac hypertrophy is primarily induced by chronic activation of the myocardial renin angiotensin system (RAS), resulting in elevated levels of angiotensin II (Ang II). In a recent study, Lyu et al. (2015) showed a critical role of exosomal intercellular communication between cardiac fibroblasts (CFs) and cardiomyocytes in cardiac pathological hypertrophy and heart failure. They found that Ang II treatment significantly increased exosome production from CFs, which, in turn, upregulated RAS in cardiomyocytes via exosome-mediated upregulation of renin, angiotensinogen, Ang II receptor types 1 and 2 (AT2R), downregulation of angiotensin-converting enzyme 2, and ultimately enhanced Ang II production in cultured cardiomyocytes, resulting in intensification of Ang II-induced pathological cardiac hypertrophy (Lyu et al., 2015). In a pioneering work, Bang et al. (2014) detected exosome-mediated communication between CFs and cardiomyocytes. CFs were shown to secrete exosomes enriched with miR-21*, so-called “star” miRNA, which is usually degraded during miR biogenesis. Importantly, fibroblast-derived exosomal miR-21* can be taken up by cardiomyocytes in a temperature- and actin-dependent manner, which further alters the expression of the miR-21* targets SORBS2 (sarcoplasmic protein sorbin and SH3 domain-containing protein2) and PDLIM5 (PDZ and LIM domain 5), leading to cellular hypertrophy. Meanwhile, antagonism of miR-21* attenuated the development of cardiac hypertrophy in mice infused with Ang II, thus confirming exosomal miR-21* as a crucial paracrine signaling mediator of cardiomyocyte hypertrophy and a potential potent therapeutic target in cardiac failure. Moreover, van Balkom et al. (2013) identified EC-derived exosomes enriched with miR-214 (an miR that controls EC function and angiogenesis) as a crucial paracrine signaling mediator in crosstalk between ECs in the suppression of senescence and stimulation of blood vessel formation in an miR-214-dependent manner. Other researchers also found that miRs such as miR-21, miR-125a, miR-135b, and miR-145-5p can be transferred by exosomes and promoted angiogenesis through up-/downregulation of its target genes in ECs (Umezu et al., 2014; Liang et al., 2016; Liu et al., 2016; Thuringer et al., 2016; Wang et al., 2016).
Not only signaling between ECs, but also intracellular communication between cardiac progenitor cells (CPCs) via an exosome-mediated mechanism is crucial for the establishment and maintenance of vascular integrity and to regenerate scarred or poorly perfused myocardium. CPCs are a small population of stem-like cells existing in the heart with particularly specialized function appropriated to cardiac pathologies. Several studies have reported the therapeutic potential of CPC exosomes. In one, Vrijsen demonstrated that CPC exosomes enhanced endothelial migration via an extracellular matrix metalloproteinase inducer (EMMPRIN)-mediated mechanism, which resulted in increased capillary density, indicating angiogenic effects (Vrijsen et al., 2010). In a parallel study, CPC exosomes were shown to protect myoblasts from oxidative-induced apoptosis via inhibition of caspase 3/7 activation in vitro and to induce a significant reduction in myocyte cell death in an animal model of AMI (Chen et al., 2013). Recently, Gray et al. (2015) identified a diminished reparative capacity of exosomes generated by CPCs under normoxic conditions compared with those from hypoxic cells, as the hypoxic exosomes enhanced tube formation of ECs and decreased profibrotic gene expression in transforming growth factor-β-stimulated fibroblasts with covarying miR clusters. A further injection of exosomes improved cardiac function and reduced fibrosis in a rat model of IR injury, implying hypoxic CPC-derived exosomes as a naturally derived therapeutic strategy to promote recovery from heart damage (Gray et al., 2015).
Exosomes in cardiac regeneration
With the emergence of stem cells as potential therapeutic agents, attempts to use stem cells to reduce infarct size and to enhance cardiac function in animal models and patients have increased exponentially. Interestingly, careful rodent experimentation to attempt stem cell transplantation to sites of IR injury after AMI demonstrated that many of the transplanted cells did not integrate into the damaged tissue, whereas almost <6% actually persisted in the heart for 2 weeks after transplant (Weil et al., 2011; Krasnodembskaya et al., 2012). Moreover, stem cell transplantation in most animal models of AMI generally resulted in reduced infarct size, improved left ventricular ejection fraction, increased vascular density, and improved myocardial perfusion, which raises the hypothesis that transplanted stem cells might work in a paracrine manner. Surprisingly, recently, studies described that the trophic effects of paracrine secretion by mesenchymal stem cells (MSCs) that ameliorate the deleterious consequences of myocardial ischemia were mostly mediated by exosomes, which rapidly initiate cellular repair through intracellular delivery of functional proteins and possibly RNA (Lai et al., 2010a, 2010b). This group further explored the functional consequences and cardioprotective mechanisms of exosomes, and they found that the well-documented ATP deficit and increased oxidative stress and apoptosis were central derangements in IR injury (Ovize et al., 2010) that could be underpinned by significant proteomic changes in enzymes that are critical for fatty acid oxidation, glycolysis, and tricarboxylic acid cycle, as well as for generating a surplus of proapoptotic proteins. However, this deficit was complemented by an abundance of metabolic-related proteins in MSC exosomes, leading to increased levels of ATP and NADH, decreased oxidative stress, increased phosphorylated-Akt and -GSK-3β, and reduced phosphorylated-c-JNK in IR hearts (Arslan et al., 2013). Finally, the study highlighted the proteomic potential of MSC exosomes in correcting fundamental biochemical aberrations in reperfused myocardium via complementation of biochemically active enzymes. These critical proteins were, no doubt, transferred from MSC exosomes across the plasma membrane into reperfused cardiomyocytes, thereby providing a safer and cost-free “off-the-shelf” therapeutic alternative to cell-based therapy for the treatment of AMI.
Although autologous, novel stem cell therapies continue to emerge as strategies for treating various cardiovascular disorders. The major benefit of using CD34+ stem cells for the patient is that the cell source is autologous, which eliminates inflammatory responses and toxicity elicited by nonautologous cells. However, challenges remain for the natural time-dependent decrease in viability and angiogenic quality of CD34+ cells along with advanced age and diminished cardiovascular health (Taguchi et al., 2004; Valgimigli et al., 2004); thus, some measures need to be taken to improve the therapeutic efficacy and normalize the inherent reparative abilities of CD34+ cells across all patients. To solve these serious and complex problems, Mackie et al. (2012) modified CD34+ cells to express an established angiogenic protein, sonic hedgehog (Shh), to avoid age- and health-related declines in CD34+ cell function. They reported that treatment with CD34Shh induced a robust increase in capillary density of the infarct border zone, leading to reduced infarct size and enhanced myocardial retention. As exosomes derived from CD34+ stem cells have been recently characterized and were shown to replicate the angiogenic activity of their mother cells, the authors isolated exosomes from CD34Shh cells and surprisingly detected that the overwhelming majority of secreted Shh existed in exosomes. These Shh-containing exosomes effectively transferred the Shh protein cargo to neighboring cells by activating Shh signaling pathways, which illustrates a higher therapeutic efficacy of exosomes than modified cells in delivering a treatment to preserve or improve cardiac function (Mackie et al., 2012).
Despite the increasing evidence of the therapeutic efficacy of stem cell-derived exosomes, research is still in its infancy and much remains unclear. Undoubtedly, specific delivery of exosomes to target cells or organs is crucial, as selective overexpression of specific molecules in parent MSCs using genetic modification may enhance the therapeutic efficiency of exosomes, thereby advancing the feasibility of their clinical application.
Exosomes in other cardiomyopathy
Aside from traditional CVD, exosome-mediated cellular communication has also been observed in other cardiomyopathies, such as diabetes-related ischemic CVD, sepsis-induced myocardial dysfunction, and peripartum cardiomyopathy (PPCM). In diabetes mellitus, Wang et al. (2014) have shown that exosomes derived from cardiomyocytes of type 2 diabetic Goto-Kakizaki rats exert an anti-angiogenic function through transfer of miR-320 into ECs, leading to inhibited EC proliferation, migration, and tube-like formation via downregulation of miR-320 target angiogenesis-related genes, such as IGF-1, Hsp20, and Ets2.
In sepsis-induced myocardial depression, Gambim et al. (2007) described a redox-reactive role of platelet-derived exosomes in vascular signaling, which can induce EC caspase-3 activation followed by apoptosis via generation of the superoxide, nitric oxide (NO), and peroxynitrite, suggesting the potent proapoptotic effect of exosomes in ECs from septic shock patients. Meanwhile, in a parallel study, Azevedo et al. (2007) detected a significant reduction of myocardial contractility induced by platelet-derived exosomes from septic patients in isolated heart and papillary muscle preparations. Besides, the increase in myocardial nitrate content together with an intrinsic production of NO by exosomes from septic patients indicated that exosomal NO and NO derivatives interchange may, in fact, represent a novel mechanism of myocardial dysfunction in sepsis (Azevedo et al., 2007). However, the sepsis-trigged myocardial depression has been surprisingly identified by a recent study to be attenuated by the MSCs-derived exosomal transfer of miR-223 to cardiomyocytes, resulting in downregulation of Sema3A and Stat3, which play a crucial role in promotion of macrophage inflammation and cardiomyocyte death in sepsis. Therefore, this study demonstrated miR-223-bearing exosomes as a great therapeutic strategy to promote cardiac recovery from sepsis (Wang et al., 2015).
PPCM is a life-threatening heart disease manifested as a sudden onset of heart failure toward the end of pregnancy or in the months postpartum. Enhanced accumulation of the 16-kDa N-terminal prolactin fragment (16K PRL) in the peripartum heart has been proposed as a cause of PPCM (Hilfiker-Kleiner et al., 2007). Halkein et al. (2013) detected that 16K PRL exerted its angiostatic effect mostly through induction of miR-146-enriched exosomes from ECs. These exosomes were shown to be absorbed and taken up by cardiomyocytes in a paracrine manner, which eventually contributed to a subsequent decrease in cardiomyocyte metabolic activities and impaired contractile function. The study further suggested that exosomal miR 146-based intercellular communication is perhaps not merely limited to ECs and cardiomyocytes but also exists in other heart cells with protective, or alternatively, detrimental roles in disease course. (Halkein et al., 2013)
Exosomes as functional CVD biomarkers
To date, the search for new biomarkers of CVDs remains a large and growing enterprise. New surrogate markers for CVD should be accurate, sensitive, specific, quickly detectable, and noninvasively collected from readily accessible body fluids (Hoefer et al., 2015). Exosomes are nanovesicles that can be isolated from almost all body fluids, including blood, urine, saliva, breast milk (Naslund et al., 2014), cerebrospinal fluid (Akers et al., 2015), semen (Madison et al., 2015), amniotic fluid (Foster et al., 2016), and ascites (Cho, 2013). Those released from a damaged or diseased heart may exhibit changes in the profile of exosomal miRs or proteins that can mirror the cellular origin and physiological state, as a “fingerprint” or a “signature” of the donor cell. These characteristics indicate that exosomes better reflect the cellular processes associated with the pathogenesis of CVD. Therefore, exosomes may represent novel minimally invasive (using blood samples) or noninvasive diagnostic markers (based on analysis of urine and saliva) to overcome the current setbacks of traditional needle or excision biopsies that reduce patient pain and inconvenience, and are more cost-effective and efficient.
The detection of significant elevated levels of exosomal miR-133a in the serum of patients with CVDs emphasizes the potential utility of exosomes and their contents as biomarkers for AMI, as these exosomes were found to originate primarily from the injured myocardium, before any elevation of serum creatine phosphokinase or cardiac troponin (Kuwabara et al., 2011). In the sera of post-AMI patients who developed heart failure within one year of AMI onset, coordinately elevated levels of three exosomal p53-responsive miRs (miR-192, -194, and -34a) may serve as predictors for future development of heart failure (Matsumoto et al., 2013). Moreover, a pivotal cardioprotective role of miR-144 associated with RIPC has been established, which raises the possibility of exosomal miR-144 as a potential biomarker to assess the degree of cardioprotection (Li et al., 2014). Notably, compared with traditional serological biomarkers, a recent study by Cheng et al. reported that some heart-specific miRs entered the urine by transrenal release from exosome packages. They detected significant elevated levels of urine exosomal miR-1 in patients with AMI, which peaked at 24 h after AMI, and declined to the base level after 7 days. A positive correlation was demonstrated between urine miR-1 and serum TnI levels, as well as myocardial infarct sizes, which suggests a novel group of urine biomarkers for AMI (Cheng et al., 2012).
However, despite the abundance of exosomes in biological fluids, the actual use of exosomal contents (proteins or miRs) as biomarkers has not yet been put into effect in clinical practice, although several companies are currently intensively exploring the development of exosome-based diagnostics. Undoubtedly, the emergence of exosome-based molecular diagnostic tests in the context of CVD will herald a new chapter in clinical diagnosis.
Exosomes and its role in CVD gene therapy
Given the exosomal nanoscale size, the potential ability to target specific cells or tissues, and even transfer of genetic material, recent interest in exosome research directed to the use of exosomes as attractive biological vehicles for drug or gene delivery has emerged. These nanovesicles have excellent host biodistribution and biocompatibility, and the issue of immunogenicity can be circumvented owing to generation from the patient's own body fluids or cell culture (Kooijmans et al., 2012). As a result, in addition to being used as therapeutic entities themselves, exosomes could be regarded as excellent vesicles for the efficient delivery of biological therapeutics across different biological barriers to target cells. A variety of therapeutic material can be encapsulated for exosome-mediated delivery, such as short interfering-RNA (siRNA), antagomirs, recombinant proteins, and anti-inflammatory drugs, which are prone to RNase degradation and incapable of traversing biological membranes to reach target specific cells, if released free in the circulation. Excitingly, Wood et al. (2011). demonstrated the first proof and concept that exosomes can be exploited as vehicles for delivery of exogenous siRNA across the blood–brain barrier and confer a gene silencing effect on target cells via knock-down of target-specific transcription and translation. Furthermore, Momen-Heravi et al. (2014) showed that B cell-derived exosomes can be harnessed to deliver exogenous miRNA-155 inhibitors to macrophages and functionally decrease TNF-α production. They further showed that exosome-mediated delivery of miRNA-155 mimicked the liver in vivo with minimal cytotoxicity (Momen-Heravi et al., 2014). Similarly, exosomes that are artificially loaded with curcumin, an anti-inflammatory, antioxidant reagent, were shown to not only enhance delivery of curcumin to target monocytes but also improved solubility, stability, and bioavailability of curcumin (Sun et al., 2010). Considering the mechanistic role of monocytes and macrophages in CVD, these studies cooperatively demonstrated that exosomes could be used as more efficient delivery vehicles not only to improve bioavailability of drugs but also to direct specific targeting of new therapeutics with no immunogenicity or adverse effects.
Despite these exciting possibilities for cardiovascular exosome treatment, some major challenges remain before thorough clinical application. One issue is the lack of both cost- and time-effective nanotechnologies for large-scale production of exosomes. Besides, it is necessary to identify the most appropriate human cell type for exosome derivation without any influence on potency or efficacy for optimal gene therapy. As exosomes are heterogeneous, the exact mechanism behind protein and nucleic acid sorting within exosomes remains to be elucidated, which renders the outcome of potential therapeutic strategies unpredictable. Thus, a more detailed understanding of the complex structure of natural exosomes will serve to optimize the development of artificial exosome-like delivery vehicles. Exosomes represent novel frontiers in the field of nanomedicine (Fig. 2).
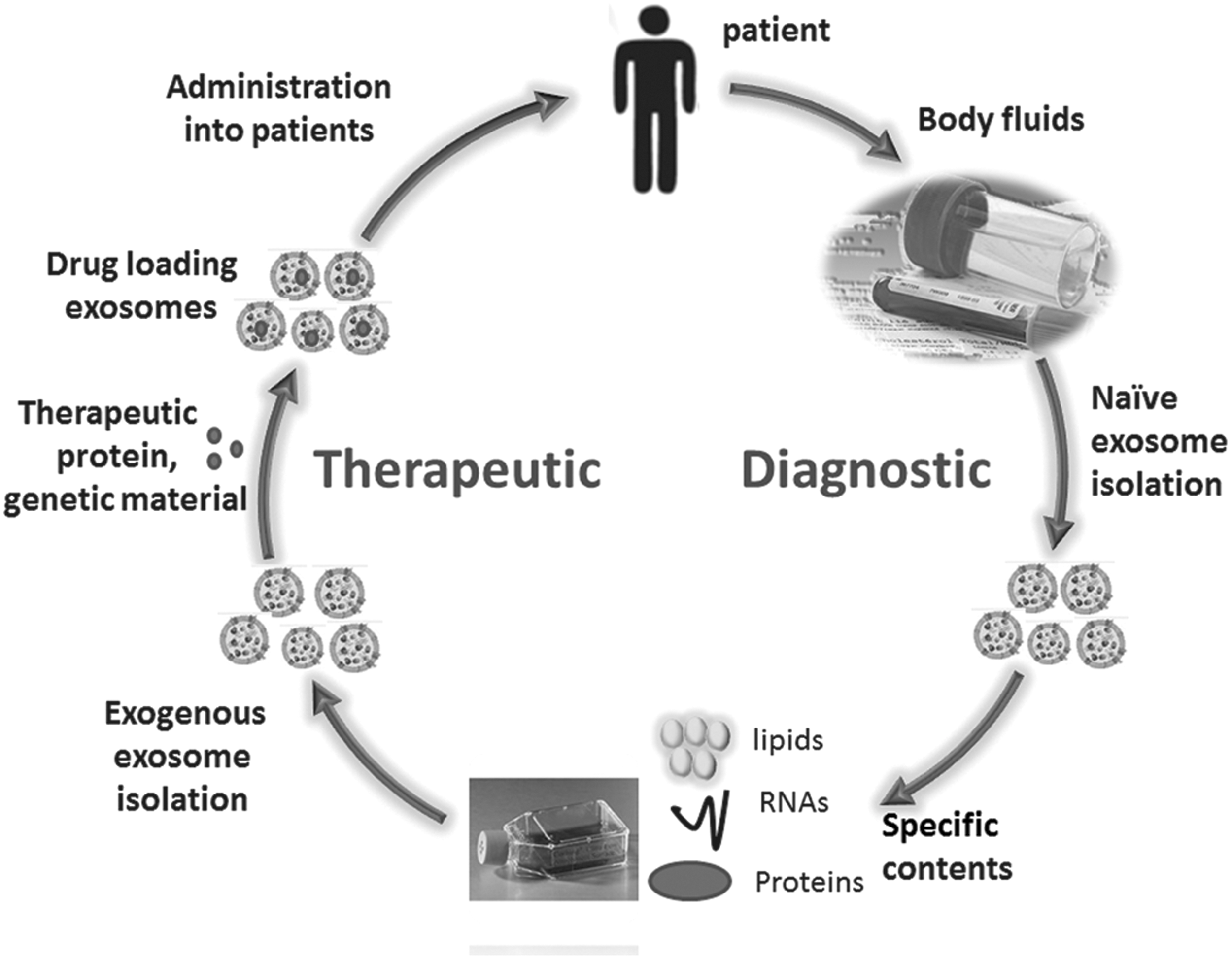
The potential use of exosomes in the cardiovascular field. Exosomes isolated from almost all body fluids carry a specific pattern of miRs, proteins, and lipids, which present potential sources of diagnostic biomarkers. Alternatively, exosomes isolated from autologous body fluids or exogenous cell culture medium can be artificially manipulated through encapsulation of therapeutic material, such as small interfering RNA (siRNA), antagomirs, recombinant proteins, and anti-inflammatory drugs, to exploit as drug delivery vehicles for clinical use.
Conclusions
Exosome research in the field of CVD is just in its infancy and relatively few studies have been conducted on such issues to date, which may be ascribed to either a lack of knowledge or technical difficulties working with exosomes. More in-depth research is warranted for fully understanding the biological aspects of loading, targeting, and delivery of exosomes, and for identifying the endogenous content of exosomes. Several unanswered questions remain, such as (1) Do cardiac exosomes released from different cell types differ from each other in terms of composition and content? (2) How do cell type-specific exosomes exert either a protective or detrimental role at the time of myocardium injury? (3) What molecules participate in the intricate intercellular communication during the development of CVD? (4) Are there qualitative and quantitative differences among exosomes from various regions of the myocardium (e.g., the infarct/border zone)? (5) Can conclusions be drawn from altered compositions and contents of lipids, proteins, and nuclear acids in exosomes for CVD early diagnosis, prognostication, and curative effect evaluation? (6) How do cardiac exosomes impact the immune responses after endothelium injury? (7) Can cardiac exosomes be harnessed for the treatment of CVD? (8) Is it possible to artificially modify exosomes to amplify their beneficial effects and/or alleviate their detrimental effects? Of course, not all components of exosomes are required to maintain biological function; thus, an alternative strategy of engineering exosome-mimic liposomes composed of only the factors required for success as nanocarriers would be more attractive (Sato et al., 2016). It is conceivable that further investigations of cardiac exosomes, as well as exosome-mediated intercellular communication in the heart will potentially aid in the clinical application of exosomes in biomarker discovery and delivery of biological therapeutics in CVD.
Footnotes
Acknowledgments
The authors hope for the understanding of all authors whose important work in the field could not be cited due to word count limitations. They would like to acknowledge their indebtedness to men of important achievements in this field, from whose lectures they benefited greatly.
This study was supported by the National Natural Sciences Foundation of China (grant numbers 81301489, 81472009, and 81572051).
Authors' Contributions
Each author substantially contributed to the review. Y.Z.: conception and design, drafting the article; Y.-W.H.: revising it critically for important intellectual content; L.Z. and Q.W.: conception and design, drafting the article or revising it critically for important intellectual content and final approval of the version to be published.
Disclosure Statement
No competing financial interests exist.