
Abstract
DNA and Cell Biology
officially retracts the article entitled, “microRNA-130a Promotes Human Keratinocyte Viability and Migration and Inhibits Apoptosis Through Direct Regulation of STK40-Mediated NF-κB Pathway and Indirect Regulation of SOX9-Meditated JNK/MAPK Pathway: A Potential Role in Psoriasis,” by Ying Xiong, Hongxiao Chen, Liqian Liu, Leihong Lu, Zongshan Wang, Fujun Tian, and Yongliang Zhao (DNA Cell Biol 2017;36(3):219–226; doi: 10.1089/dna.2016.3517).
A serious concern was brought to the attention of Dr. Carol Reiss, the Editor-in-Chief of DNA and Cell Biology, regarding the supporting data underlying the study. Dr. Reiss contacted the authors to request that all original uncropped scans of membranes used for Western blot analysis be supplied for further assessment, however the authors did not respond within the 2-week timeframe supplied. Dr. Reiss contacted the authors again, indicating that their failure to reply was indicative of their acknowledgement of misconduct.
DNA and Cell Biology
and its editorial leadership treat these reports of possible scientific misconduct very seriously and therefore officially retracts this article from the scientific literature.
Introduction
P
microRNAs (miRNAs), an extensive class of small noncoding RNAs (18–25 nt), have been shown to possess prominent effects in regulating gene function by binding to the 3′ untranslational region (3′UTR) of target messenger RNAs (mRNAs) (Rana, 2007; Thai et al., 2007; Boll et al., 2013). There is overwhelming evidence confirming that alterations in miRNA expressions are closely related to plentiful biological processes and tumorigenesis (Wang et al., 2010a; Chen et al., 2012; Tang et al., 2012). Downregulation of miR-146a, miR-19a, miR-223, miR-143, and miR-217 have been reported to be significant systemic biomarkers in psoriasis (Hirao et al., 2013; Lovendorf et al., 2014; Zhang et al., 2014; Zhu et al., 2016). Thereby, focusing on miRNAs appears to provide potential insight into the diagnosis and targeted treatment of psoriasis.
A host of literatures have corroborated that miR-130a has considerable implications for many kinds of diseases. Previous studies found that miR-130a enhanced proliferation and migration, and promoted tumorigenesis in gastric cancer, ovarian cancer, chronic lymphocytic leukemia, and colorectal cancer (Kovaleva et al., 2012; Liu et al., 2013; Zhang et al., 2013; Jiang et al., 2015). However, in breast cancer, non-small cell lung cancer, and arteriosclerosis obliterans (ASO), miR-130a was considered as a tumor suppressor to inhibit cell proliferation, migration, and invasion (Wang et al., 2010b; Li et al., 2011; Pan et al., 2015), which revealed the complexity and diversity of miR-130a in diseases. Nevertheless, the role and mechanism of miR-130a in psoriasis remain unknown.
The aim of this study was to determine the regulating role of miR-130a in keratinocyte viability, apoptosis, and migration along with underlying mechanism, which is expected to lay a theoretical basis for clinical therapy of psoriasis.
Materials and Methods
Patients
All psoriasis specimens were collected from 12 patients (7 male and 5 female, 14–54 years old) with psoriasis during April 2015 to March 2016 in Linyi People's Hospital. The average psoriasis area and severity index (PASI) score was 20.6 ± 7.12. Punch biopsies (4-mm) were obtained by histopathologic biopsy and were immediately stored at −80°C for further studies. The subjects had not received systemic immunosuppressive treatment for at least 1 month or topical therapy for at least 2 weeks before skin biopsy. Additionally, normal skin tissues (4-mm) were obtained from eight healthy controls undergoing plastic surgery at the same time in Linyi People's Hospital. They were 18–40 years old, including two men and six women. This study was approved by the Ethics Committee of Linyi People's Hospital and all patients or guardians (patients who were <18 years old) signed informed consent.
Cell culture
Human keratinocyte cell line HaCaT was purchased from German Cancer Research Center (Eppelheim, Germany). HaCaT cells were maintained in Dulbecco's modified Eagle's medium (DMEM; Lonza) supplemented with 10% fetal bovine serum (FBS; Hyclone Laboratories, Logan, UT), 1.4 mM
miR-130a overexpression and silencing transfection
HaCaT cells were incubated at 1 × 105 cells per well of six-well plates overnight. The miR-130a mimic (50 nM), inhibitor (150 nM), and miR-control (150 nM) (GenePharma Co., Shanghai, China) were transfected into HaCaT cells on the next day, respectively. Cell transfections were carried out using Lipofectamine 3000 reagent (Invitrogen, Carlsbad, CA) based on manufacturer's introductions. Furthermore, the small interfering RNA (siRNA) specific serine/threonine kinase 40 (STK40; 50 nM) or sex-determining region Y chromosome-box 9 (SOX9; 50 nM; GenePharma Co.) was transfected into the cells after aberrant expression of miR-130a.
Cell viability assay
The HaCaT cell viability was determined using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) colorimetric assay (Xu et al., 2015). After transfection, HaCaT cells were cultured in DMEM (with 10% FBS) containing 0.5 mg/mL MTT (Sigma) on days 1, 2, 3, and 4 and 100 μL dimethylsulfoxide (DMSO; Lonza) was added to dissolve the blue formazan (Sigma) product for 1 h. Then, cell viability was measured by absorbance at a wavelength of 490 nm.
Detection of apoptosis
Apoptotic cells were identified and quantified by flow cytometry with Annexin V-FITC/Propidium iodide (PI) apoptosis detection kit (Beijing Biosea Biotechnology, Beijing, China). After transfection, HaCaT cells (1 × 105 cells/well) were seeded in six-well culture plate respectively and were washed twice with cold phosphate buffer saline. Then, they were co-incubated with serum-free culture medium containing 10 μM dichlorofluorescein diacetate (20 min, 37°C, in dark). Subsequently, samples were collected, centrifuged, resuspended in 100 μL annexin-binding buffer, and then were measured with flow cytometer according to the manufacturer's protocol.
Migration assay
For the migration assay, transwell assay with a pore size of 8 μm and 6.5 mm polycarbonate transwell filters (Corning, Lowell, MA) was determined (Poincloux et al., 2011). Complete medium (600 μL) was added to the bottom compartment. HaCaT cells were suspended in 200 μL of serum-free DMEM medium (Sigma) in 5% CO2 at 37°C for 24 h. Then, they were incubated on the top chamber of 24-well. After 12 h, HaCaT cells that attached to the lower surface of the membranes were fixed with 4% methanol (Sigma) for 30 min and stained with 0.1 g/mL crystal violet (Beyotime Institute of Biotechnology, Haimen, China). The number of HaCaT cells on the lower surface of the filter was counted under a light microscope (Biotech).
Target prediction and luciferase reporter assay
Target genes of miR-130a were predicted by bioinformatics analysis using TargetScan 6.2 (
Quantitative reverse transcription PCR
Total RNA of HaCaT cells in each treatment were isolated, respectively, using TRIzol reagent (Invitrogen) and treated with DNaseI (Promega, Madison, WI). A total of 2 μg RNA was used to synthesize poly-oligo (dT) primed complementary DNA (cDNA) with the RevertAid H Minus First strand cDNA Synthesis Kit (Thermo Fischer Scientific, Inc., Waltham, MA). Primers for miR-130a and U6 were produced by the miScript Primer Assay kit (Qiagen, Dusseldorf, Germany): miR-130a, forward 5′GGAATTGCAATGCTGAGGAG′3, reverse 5′GCCGTTTTCTTTGAGGACTG′3; U6, forward 5′CGCTTCGGCAGCACATATACTA′3, reverse 5′CGCTTCACGA ATTTGCGTGTCA′3. The relative expression of miR-130a was normalized to the internal control (U6) using the 2−ΔΔCt method.
Quantitative reverse transcription (qRT)-PCR reactions for c-Jun, c-Jun N-terminal kinase (JNK), and p38 mitogen-activated protein kinase (MAPK) were performed using RiboMAX Large Scale RNA Production System T7 (Promega, Karlsruhe, Germany). The specific primer sequences were as follows: STK40, forward 5′CATCCTTGGTCCCCGTCTGG′3; reverse 5′CCTCTCCTCCAGGGTCAGGA′3; c-Jun, forward 5′GGAGGGAGGTTTGTGAGAGC′3, reverse 5′ACAAACAACAC TGGGCAGGA′3; JNK, forward 5′AACTGTTCCCCGATGTGCT′3, reverse 5′TCTCTTGCCTGACTGGCTTT′3; p38MAPK, forward 5′ACTGCCAAGGAGCA TCTA′3, reverse 5′GAAGAGCCTGACCTACAGT′3; glyceraldehyde-3-phosphate dehydrogenase (GAPDH; Abcam, Cambridge, United Kingdom), forward 5′ATCTGGCACCACACCTTCTACA′3, reverse 5′GTTTGGTGGATGCCACAGGA CT′3.
Western blot analysis
After transfection, the proteins were extracted using radio-immunoprecipitation assay (RIPA) lysis buffer (Beyotime Biotechnology, Shanghai, China) with protease inhibitors (Applygen Technologies, Inc., Beijing, China). The protein was quantified by Bicinchoninic Acid (BCA) Protein Assay Kit (Pierce, Appleton, WI), subjected to 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and blotted on nitrocellulose membrane (Millipore). GAPDH was regarded as an internal control.
Primary antibodies SOX9 (ab26414), c-Jun (ab32137), p-c-Jun (ab32385), JNK (ab76125), p-JNK (ab4821), p38 MAPK (ab197348), p-p38 MAPK (ab47363), and STK40 (ab77653) were obtained from Abcam. After incubation with the membrane at 4°C overnight, secondary antibodies were marked by horseradish peroxidase for 1 h at room temperature. Then, images were developed and photographed using Image Lab Software (Bio-Rad, Shanghai, China).
Statistical analysis
Each experiment was carried out in triplicate with three independent replicates. All results were presented as mean ± standard deviation. Values were performed by one-way analysis of variance with SPSS 19.0 software (SPSS, Chicago, IL). Statistical significance was defined as p < 0.05.
Results
miR-130a expressions in psoriasis tissues
To investigate the functional role of miR-130a in psoriasis, we enrolled a total of 12 patients with psoriasis and 8 healthy controls, and assessed the expression patterns of miR-130 by qRT-PCR. As shown in Figure 1, the relative expression levels of miR-130a in psoriasis tissues were significantly higher than normal skin tissues (p < 0.01). It revealed that miR-130a might play a significant role in the development of psoriasis.
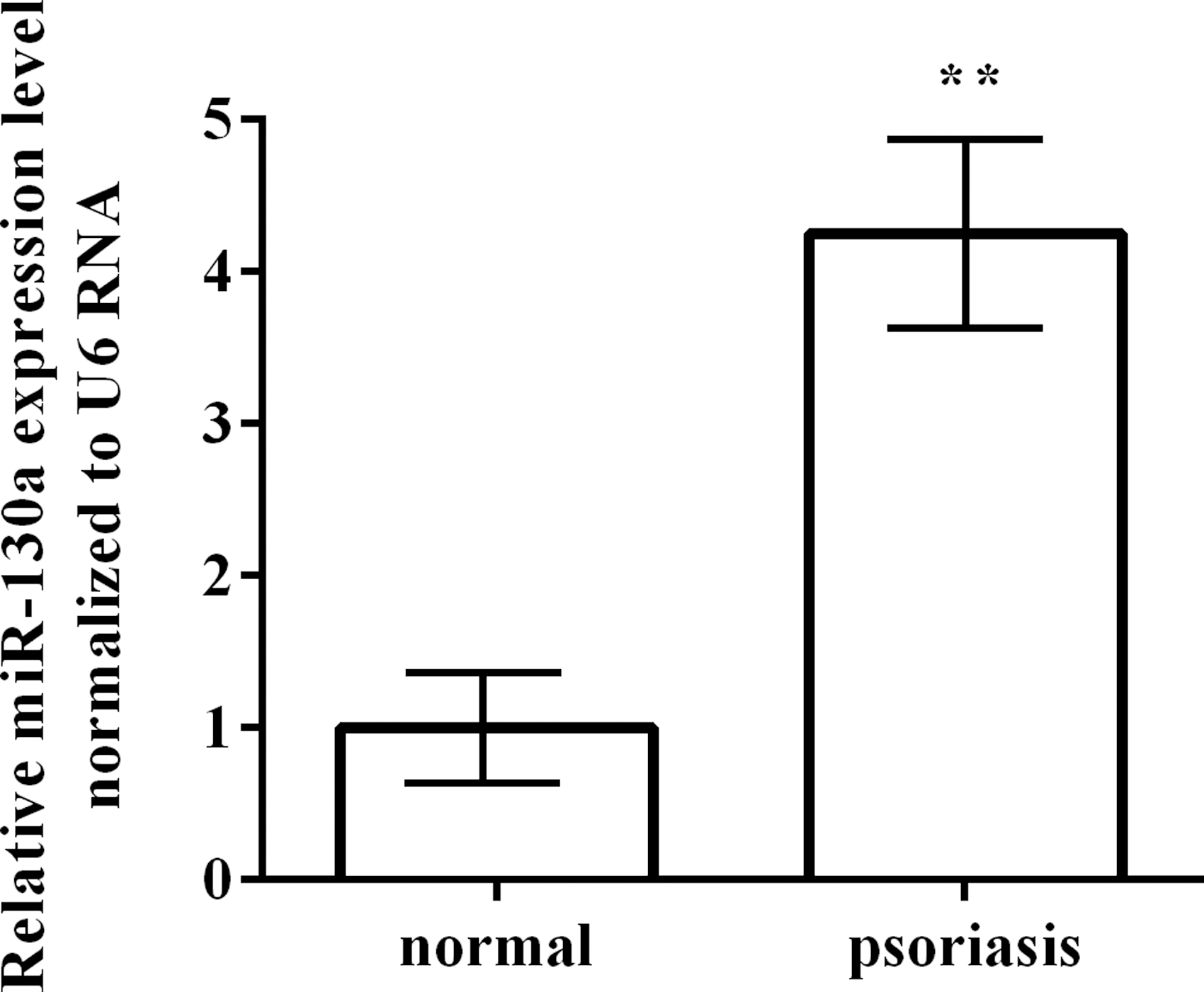
Expression levels of miR-130a in patients with psoriasis and normal skins. **p < 0.01. Each assay was performed in triplicate with three independent replicates.
Transfection efficiency of miR-130a in HaCaT cells
The miR-130a mimic or anti-miR-130a was transferred to HaCaT cells, respectively, to examine the effect of miR-130a on keratinocytes. After transfection with the miR-130a mimic, the expression of miR-130a was remarkably higher than that in the miR-control (Fig. 2, p < 0.01). Meanwhile the anti-miR-130a significantly reduced the expression of miR-130a compared with the miR-control (p < 0.01). The results indicated that transfection efficiency was high and could be used for further study.
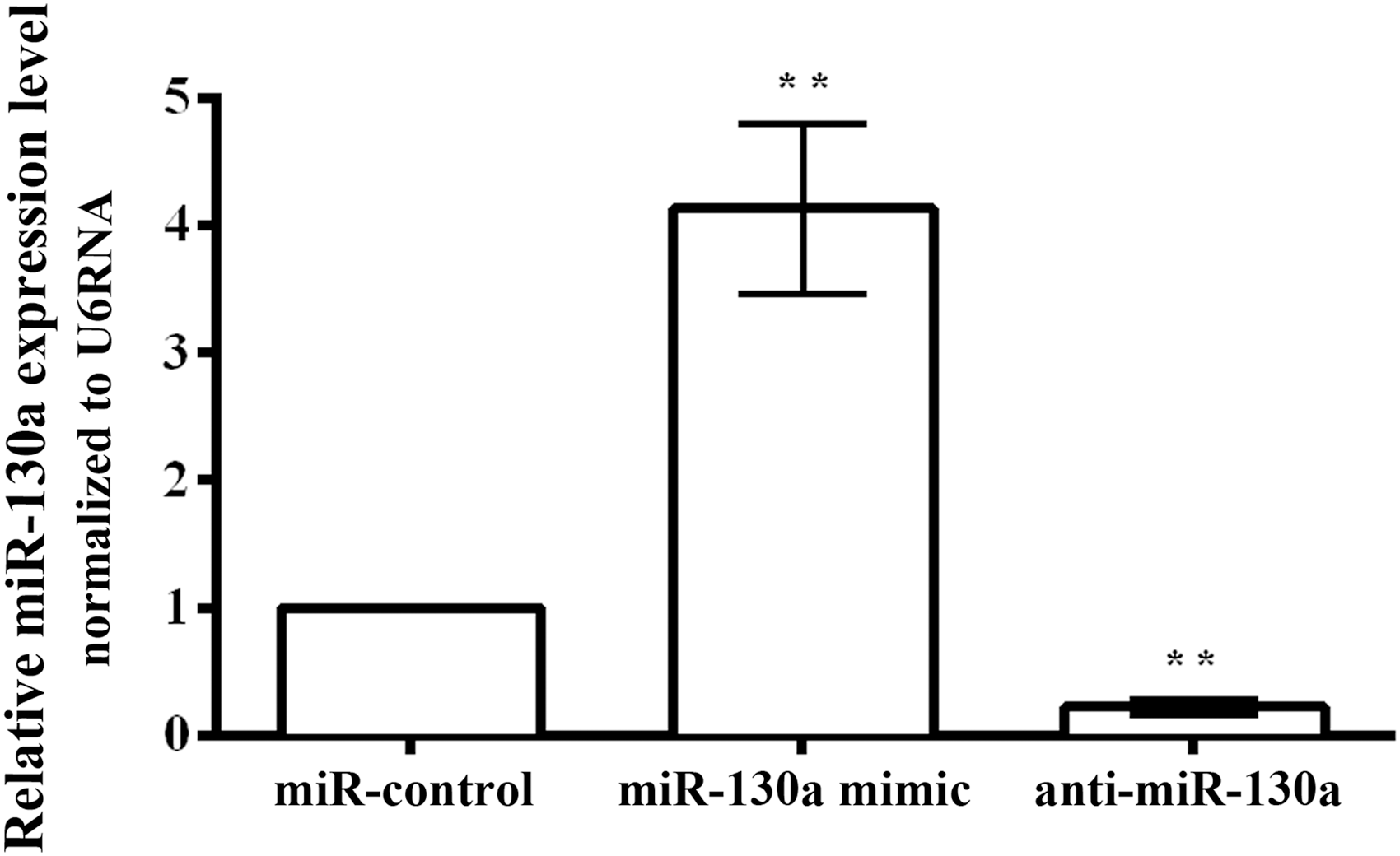
Transfection efficiency of miR-130a in HaCaT cells. **p < 0.01. Each assay was performed in triplicate with three independent replicates.
Overexpressing miR-130a promoted cell viability in HaCaT cells
The cell viability was explored by MTT assay in HaCaT cells (Fig. 3). Overexpression of miR-130a increased cell viability in HaCaT cells at 2 and 3 days compared with the miR-control, however, there was no significant difference with the miR-control (p > 0.05). Overexpression of miR-130a statistically upregulated cell viability at 4 days (p < 0.05). Additionally, the silence of miR-130a did not observably reduced cell viability until the 4 days compared with the miR-control (p < 0.05). In consequence, overexpressing miR-130a promoted cell viability in HaCaT cells.
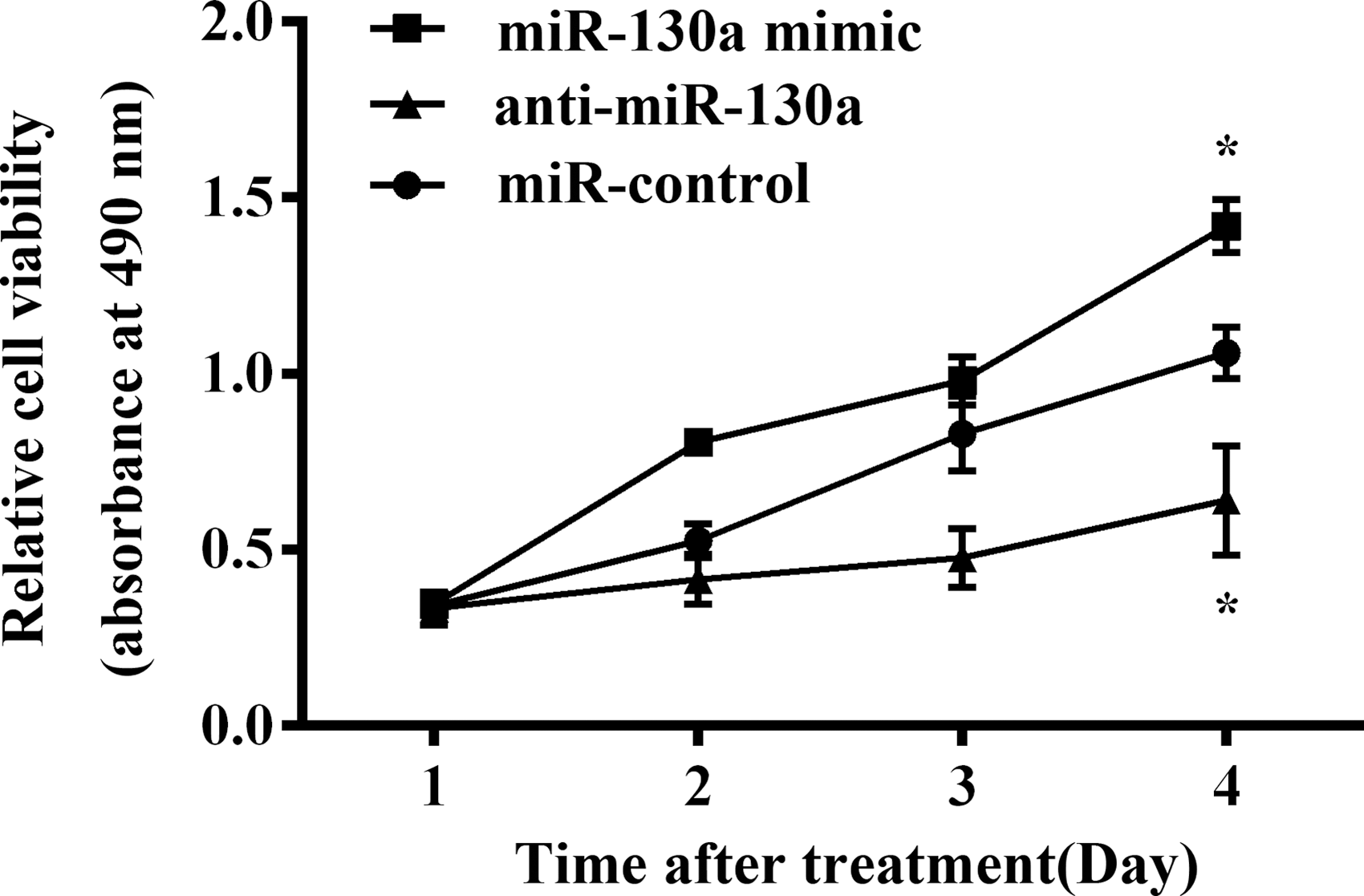
Overexpressing miR-130a promoted cell viability in HaCaT cells. *p < 0.05. Each assay was performed in triplicate with three independent replicates.
Overexpressing miR-130a inhibited cell apoptosis in HaCaT cells
Apoptotic HaCaT cells were identified and quantified by flow cytometry. Figure 4A and B showed that apoptotic cells in miR-130a mimic group (1.3%) were significantly lower than that in miR-control (3.1%; p < 0.05). Anti-miR-130a (13.4%) statistically generated more apoptotic cells compared with the miR-control (p < 0.01). As a result, overexpressing miR-130a inhibited cell apoptosis in HaCaT cells.

Overexpressing miR-130a inhibited cell apoptosis in HaCaT cells.
Overexpressing miR-130a promoted cell migration in HaCaT cells
The migrating capacity of HaCaT cells were identified by transwell assay (Fig. 5). Overexpressing miR-130a strikingly enhanced cell migration in HaCaT cells compared with the miR-control (p < 0.01). Anti-miR-130a statistically weakened cell migration in HaCaT cells compared with the miR-control (p < 0.05). Thus, overexpressing miR-130a promoted cell migration in HaCaT cells.

Overexpressing miR-130a promoted cell migration in HaCaT cells. *p < 0.05; **p < 0.01. Each assay was performed in triplicate with three independent replicates.
STK40 was predicted as a targeting gene of miR-130a and was involved in miR-130a-induced cell functions
Two open access programs (TargetScan 6.2 and

STK40 was predicted as a targeting gene of miR-130a and was involved in miR-130a-induced cell functions.
We further explored whether STK40 was involved in miR-130a-induced cell functions. Both the expression of STK40 and miR-130a were downregulated and then the effects on cell viability, apoptosis, and cell migration were analyzed again. As shown in Figure 6E–G, the results revealed that silencing of STK40 significantly increased the cell viability at 4 days (p < 0.05), decreased the cell apoptosis (p < 0.01), and elevated the migration ability (p < 0.01) compared to the control group. However, the above effects were reversed by simultaneous transfection with anti-miR-130a. These results suggested that STK40 was predicted as a targeted gene of miR-130a and that STK40 was involved in miR-130a-induced cell functions.
miR-130a regulated STK40-mediated NF-kB pathway and SOX9-mediated downstream JNK/MAPK signaling pathway
To further understand the mechanism of miR-130a in keratinocytes, we evaluated the impact of miR-130a aberrant expression on STK40-mediated NF-κB pathway key protein NF-κB p65. Previous studies pointed out that STK40 was a negative regulator of NF-κB (Huang et al., 2003). As shown in Figure 7A, we found that the expression levels of NF-κB p-p65 was markedly increased by miR-130a mimic but was decreased by anti-miR-130a compared to the control group. However, simultaneous silencing of miR-130a and STK40 had no obvious changes on the levels of p-p65. The results indicated that miR-130a activated the STK40-mediated NF-κB pathway. In addition, it has been reported that Sox9 was highly expressed in psoriasis (Shi et al., 2013). Therefore, we assumed that there might be an underlying regulatory relationship among miR-130, STK40, and SOX9.
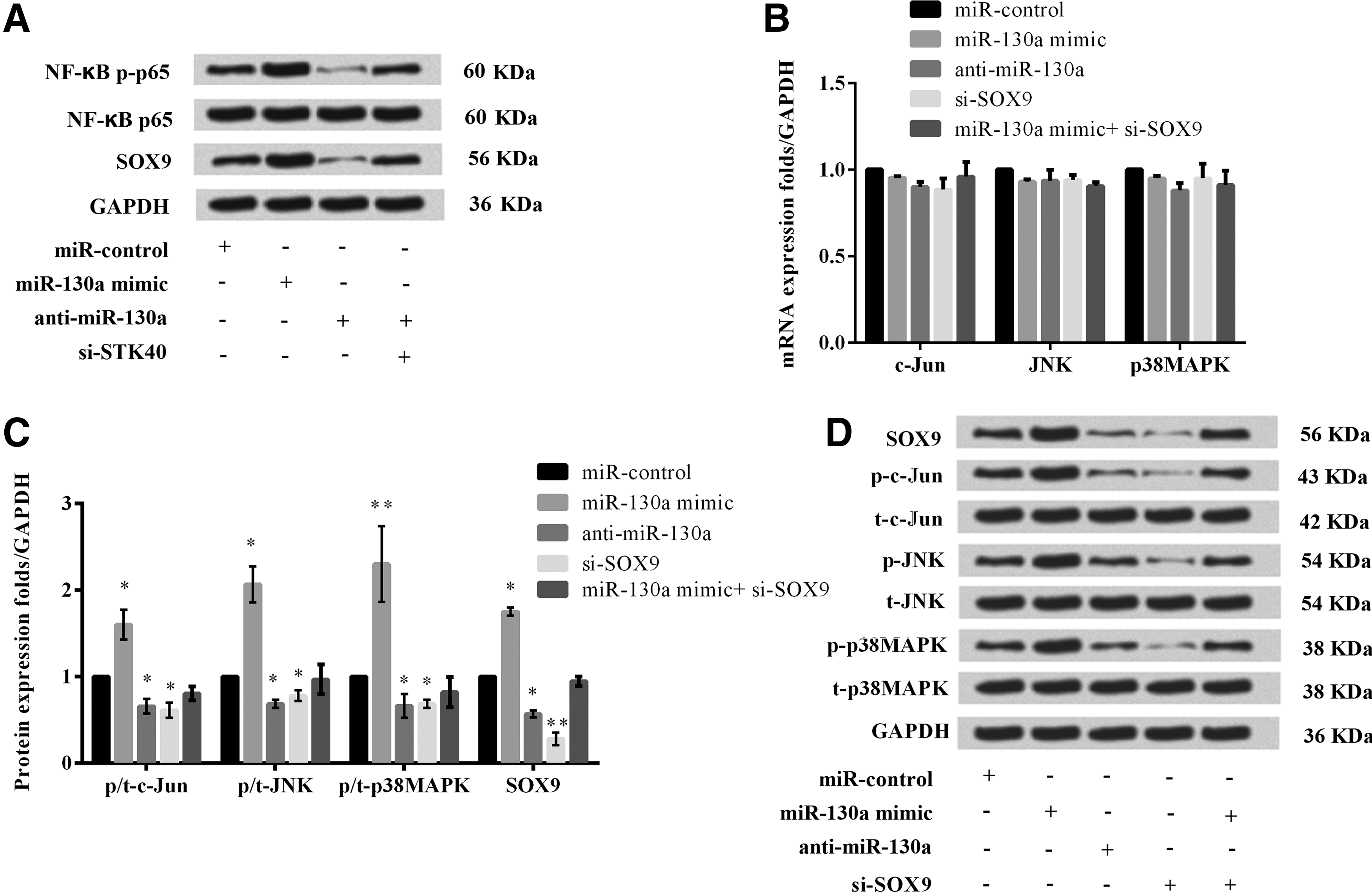
MiR-130a regulated STK40-mediated NF-κB pathway and SOX9-mediated downstream JNK/MAPK signaling pathway.
Interestingly, we found that the levels of SOX9 were obviously increased by miR-130a overexpression but decreased by miR-130a downregulation. However, simultaneous silencing of STK40 alleviated the effects of miR-130a downregulation on expression of SOX9. The results implied that miR-130a directly regulated the expression of STK40 but indirectly regulated the level of SOX9.
We further investigated the effects of miR-130a aberrant expression on SOX9-mediated downstream JNK/MAPK signaling pathway key proteins expression. The results showed that the mRNA expression of c-Jun, JNK, and p38MAPK had not been significantly changed by aberrant expression of miR-130a or SOX9 (Fig. 7B). However, Figure 7C and D showed that overexpression of miR-130a observably raised the protein expressions of p-c-Jun, p-JNK, p-p38MAPK, and SOX9 (p < 0.05 or p < 0.01). Anti-miR-130a or silencing of SOX9 significantly reduced the expressions of them (p < 0.05 or p < 0.01). There was no difference in the protein expressions of p-c-Jun, p-JNK, and p-p38MAPK between miR-130a mimic+si-SOX9 and the miR-control. These results indicated that miR-130a activated JNK/MAPK signaling pathway by upregulating SOX9.
Discussion
In the present study, we found that the miR-130a levels were significantly higher in patients with psoriasis. Overexpressing miR-130a predominantly promoted cell viability and migration and inhibited apoptosis in HaCaT cells, and silencing miR-130a showed contrary results. In addition, we confirmed that STK40, a negative regulator of the NF-κB pathway, was a direct targeted gene of miR-130a, and it was involved in miR-130a-induced cell functions. Further mechanism study identified that miR-130a observably upregulated NF-κB p65, SOX9 and key proteins in the downstream JNK/MAPK signaling pathway. After silencing SOX9, JNK/MAPK pathway could not be activated, which hinted the mediated effect of SOX9. Moreover, silencing STK40 alleviated the effects of anti-miR-130a on SOX9 expression.
The role of miR-130a is different in distinct types of cells. In the present study, overexpressing miR-130a promoted cell viability and migration in human keratinocytes, which was in line with the roles in multiple cancers, such as gastric cancer, ovarian cancer, chronic lymphocytic leukemia, and colorectal cancer (Kovaleva et al., 2012; Liu et al., 2013; Zhang et al., 2013; Jiang et al., 2015). Psoriasis is an inflammatory skin disease resulting from hyperproliferation of epidermal keratinocytes (Mease and Armstrong, 2014). We first enrolled patients with psoriasis, with healthy specimens as controls. The results demonstrated a higher expression of miR-130a in patients with psoriasis than healthy controls. Thus, it is indicative of the potential pathogenic role of miR-130a in psoriasis.
In this study, we demonstrated that miR-130a directly targeted STK40 in keratinocytes. STK40 is a putative serine/threonine kinase, which was previously assumed to inhibit tumor necrosis factor (TNF)-induced NF-κB activation (Huang et al., 2003). Interestingly, a previous study suggested that STK40 expression was regulated by miR-31 and contributed to the inflammatory response in keratinocytes (Xu et al., 2013). In our study, STK40 showed a reciprocal expression pattern with miR-130a. Suppression of STK40 reversed the effects of anti-miR-130a on cell viability, apoptosis, and migration in keratinocytes. Activation of NF-κB is well known to produce inflammatory cytokines (Cho et al., 2007) and has also been reported to be involved in the apoptotic-resistant phenotype of keratinocytes (Qin et al., 1999).
STK40 silencing rescued the effects of activation of NF-κB induced by miR-130a. Our results at least partly explained the antiapoptosis effects of miR-130a on keratinocytes. It has been previously shown that SOX9 was closely associated with skin diseases. SOX9, one of sex-determining region Y (Sry) box related gene family, is a transcription factor that fulfills a number of developmental processes. SOX9-dependent mesenchymal signals mainly mediate morphogenesis of epithelium (Singh et al., 2010). Besides, SOX9 activation was considered to be a general hallmark of basal cell carcinoma (BCC) and adnexal skin neoplasms, which was indicative of a contribution of SOX9 to the pathogenesis of these tumors (Vidal et al., 2008).
Shi et al. (2013) proposed that SOX9 was significantly upregulated in several skin diseases including psoriasis, BCC, and squamous cell carcinoma as a vital regulator of epidermal keratinocytes with pro-proliferation and pro-survival functions. In this study, we observed that SOX9 was positively regulated by miR-130a. However, STK40 silencing also rescued the effects of miR-130a on expression of SOX9, and indicated that the effect of miR-130a on keratinocytes was achieved by directly regulation of STK40 but indirectly regulation of SOX9. Moreover, our data revealed that overexpressing miR-130a also upregulated the expressions of p-c-Jun, p-JNK, and p-p38MAPK proteins in HaCaT cells and anti-miR-130a downregulated them. But the effects of si-SOX9 and overexpression of miR-130a were neutralized on these key proteins, supporting that SOX9 inactivation might be a promising approach for the therapy of psoriasis. MAPKs, comprising JNK, extracellular regulated protein kinase (ERK), ERK5 and p38 pathways, have been implicated in numerous aspects of key cellular processes (Wang et al., 2016). Accumulating studies have shown that JNK mediated the induction of autophagic cell death of cancer cells (Park et al., 2009; Shimizu et al., 2010; Zhang et al., 2016) and c-Jun and JNK phosphorylation involves activation of apoptotic signaling (Hong and Kim, 2007; Dhanasekaran and Reddy, 2008). The seemingly paradoxical results with regard to the regulatory role of miR-130a in apoptosis might be explained by activation of NF-κB pathway induced by miR-130a mimic.
In summary, miR-130a regulates human keratinocyte HaCaT viability, migration and apoptosis might be by direct regulation of STK40-mediated NF-κB pathway and by indirect regulation of SOX9-mediated downstream JNK/MAPK signaling pathway. It is concluded that miR-130a may be a potential targeted treatment of psoriasis.
Footnotes
Disclosure Statement
No competing financial interests exist.