
Abstract
The phenotypic modulation of vessel smooth muscle cells (VSMCs) plays a crucial role in the physiological and pathological conditions of vasculature in response to local environmental changes. The phenotypic transition of VSMCs is largely modulated by the serum response factor (SRF). miR-181a and miR-181b are members of the well-studied miR-181 family and both have complementary sequence in the 3′ untranslated region (UTR) of SRF gene. In this article, evidence insinuates that miR-181a/b was involved in VSMCs differentiation through upregulating synthetic marker genes and downregulating contractile ones, respectively. We also confirmed the roles of the miR-181a/b in promoting SMC proliferation, migration, and synthetic phenotype transformation. In addition, miR-181a/b was indicated as directly targeting at 3′ UTR of SRF by dual-luciferase assay and Western blot assay. In a word, miR-181a/b is one of the factors involved in VSMC differentiation toward a synthetic phenotype through targeting at SRF. These findings may provide a potential therapeutic approach that miR-181a/b took part in regulating the vessel disorders.
Introduction
V
Serum response factor (SRF, c-fos serum response element-binding transcription factor) plays a necessary role in the development of vascular smooth muscle cells in various animals, including humans (Miano, 2003). SRF is a key transcription factor that binds to CArG boxes in the promoter regions of SMC contractile genes and, thus, activates gene transcription during SMC differentiation (Miano, 2003). Some genes, including SM22α (Kumar and Owens, 2003), smooth muscle myosin heavy chain (SM-MHC) (Miano et al., 1994), α-smooth muscle actin (α-SMA) (Mack and Owens 1999), calponin (Mack et al., 2001), and smoothelin (van der Loop et al., 1996), are used as the marker of contractile phenotype for SMC. Similarly, some other genes such as cellular retinol binding protein (CRBP)-1 (Sluiter et al., 2013), Moesin (Doevendans and van Eys, 2002), SMemb (Yoshida and Owens, 2005), osteopontin (OPN) (Speer et al., 2002), and matrix Gla protein (MGP) (Lai et al., 2002) are used as a maker of synthetic SMC. Myocardin (MYOCD) is one of the major cofactors acting on the SRF in the vascular development process and also responsible for SMC differentiation and has diverse roles in normal homeostasis and disease (Du et al., 2003; Miano, 2015). However, the regulation mechanism of SRF in VSMC remains complex, and novel evidence was needed to help elaborate the process through which SRF alters VSMC behavior.
MicroRNAs (miRNAs) are recently discovered negative regulators of gene expression and act at the posttranscriptional level (Urbich et al., 2008). MiRNAs downregulate the gene expression by binding to the 3′ untranslated region (UTR) of mRNA transcripts through a target recognition sequence known as a ‘seed sequence’ (Huntzinger and Izaurralde, 2011). The differentiation and dedifferentiation of smooth muscle cells could be regulated by specific miRNAs (Song and Li, 2010). In vascular biology, miRNAs play a major role in the capability of VSMCs to phenotypically switch from a contractile to a synthetic state (Robinson and Baker, 2012). Altering the expression level of miR-21, miR-143, and miR-145 can change the synthetic phenotype of VSMC in vivo (Cordes et al., 2009; Elia et al., 2009; Sarkar et al., 2010).
In bioinformatic analysis, we discovered that the 3′ UTR of SRF gene has complementary sequence of miR-181a and miR-181b, which indicated that SRF was one of the possible targets of miRNA 181a/b (Lin et al., 2016). MiR-181a and miR-181b are well-studied members of the miR-181 family, which play diverse roles in regulating key aspects of cellular growth, development, and activation (Sun et al., 2014; Li et al., 2015). Therefore, we proposed that miR-181a/b might regulate the phenotype modulation of VSMCs depending on SRF. Our result showed that the contractile phenotype marker gene expression was downregulated and synthetic marker gene expression upregulated by miR-181a/b. Further research showed that miR-181a/b could target on 3′ UTR of SRF and simultaneously regulate the mRNA and protein level of SRF. In this study, we provide evidence that miR-181a/b is involved in the phenotypic modulation of VSMC through targeting SRF.
Materials and Methods
Cell culture, treatment, and transfection
Human aortic smooth muscle cells (HAoSMCs) were cultured in a VascuLife SMC medium (Lifeline Cell Technology). PDGF (10 ng/mL) was used to treat HAoSMCs. HAoSMCs were transfected with miR-181a/b mimics/inhibitors (Table 1) or negative control scramble sequences (GenePharma, Shanghai, China) using Lipofectamine RNAiMAX (Invitrogen, CA) according to the manufacturer's instructions. The cells were transfected with vector SRF-pGL3 or pGL3 using a Lipofectamine 2000 reagent (Invitrogen).
RNA extraction and real-time PCR
Total RNA was extracted from HAoSMCs using a miRcute miRNA isolation kit (Tiangen, Beijing, China) according to the manufacturer's protocol. Reversed transcription was conducted using a FastQuant RT kit (Tiangen) for mRNA and a miRcute miRNA first-strand cDNA synthesis kit (Tiangen) for miRNA. Real-time quantitative PCR (qPCR) was performed using a miRcute miRNA qPCR detection kit (Tiangen) with U6 used as an endogenous control for determination of miR-181a/b expressions. SuperReal PreMix (SYBR Green) detection kit (Tiangen) was used for examination of SMC marker gene expression with GAPDH as an internal control. Primers were synthesized from Sangon Biotech (Sangon, Shanghai, China), and the sequences of primers are listed in Table 2. Specific primers for human miR-21, miR-143, miR-145, and internal control U6 snRNA were purchased from GenePharma Co., Ltd (GenePharma).
Western blotting
Proteins were extracted using a radioimmunoprecipitation assay buffer (50 mM Tris-HCl [pH 7.4], 150 mM NaCl, 1% NP-40, 0.1% sodium dodecyl sulfate [SDS]) with a proteinase inhibitor (Huaxingbio Biotechnology, Beijing, China). Protein samples were loaded and separated in 12% SDS polyacrylamide gel electrophoresis (SDS-PAGE; Huaxingbio Biotechnology). Protein bands were transferred to polyvinylidene difluoride membranes (Millipore, Bedford, NY) and blocked with 5% nonfat milk for 1 h at room temperature. The membranes were incubated with anti-SRF rabbit monoclonal primary antibodies (CST, Boston) and a horseradish peroxidase-conjugated goat-anti-rabbit secondary antibody (Sigma-Aldrich, MO), followed by visualization using enhanced chemiluminescence.
miRNA target prediction
A perl script was downloaded from TargetScan to predict potential binding sites of miR-181a/b (
Dual-luciferase assays
The 3′ UTR of SRF was cloned into pGL3 luciferase reporter vector (Qinglan Biotech, Suzhou, China). The reporter vectors and miRNA mimics were cotransfected into HAoSMCs. At 24 h after transfection, luciferase activity was measured using a Dual-Luciferase® Reporter Assay System (Promega BioSciences, CA).
Cell proliferation and migration assay
About 2000/well HAoSMCs were seeded into 96-well plates. After transfection with miR-181a/b, miR-181a/b inhibitor, and negative control, cell proliferation activity was determined by the Cell Counting Kit-8 (CCK-8, No. C0042 from Beyotime Institute of Biotechnology, Jiangsu, China) according to the manufacturer's instructions.
Migration assays were performed with a 24-well Transwell system (Corning, NY) containing an 8-μm polycarbonate membrane. After 36–48 h transfection with miRNA mimics or inhibitors, nearly 15,000 HAoSMCs were seeded on a Transwell inner chamber, induced by a cell medium containing 20% FBS in an outer chamber at 37°C for 12 h. The cells that migrated onto the outer side of the membrane were observed by an inverted microscope with crystal violet staining.
Statistical analysis
Student's t test was performed for statistical analysis. *p < 0.05 or **p < 0.01 was considered significant or very significant. Data are expressed as mean ± standard deviation.
Results
Expression level of miR-181a/b is upregulated in the proliferated HAoSMCs
PDGF has been proved to promote the shift of SMCs to synthetic phenotype by suppressing the expressions of contractile phenotype marker genes and also to promote the proliferation and migration of SMCs (Ha et al., 2015). Cultured HAoSMCs were treated with mock or PDGF in culture medium for 48 h, then the cell morphology was observed by invert microscope. It was observed that the proliferation ability of HAoSMCs was significantly enhanced by PDGF (Fig. 1A).
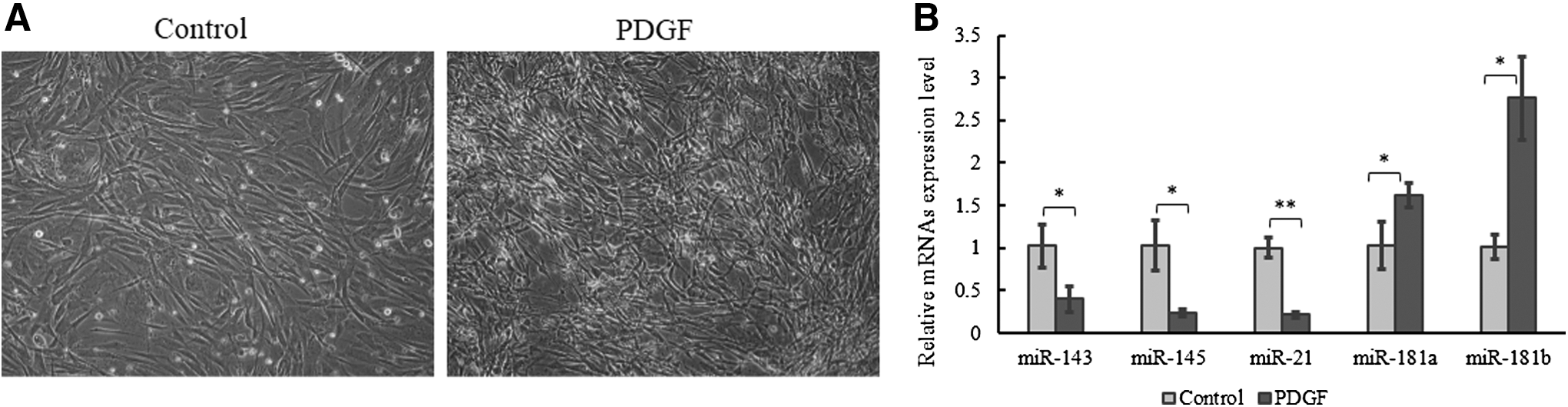
MiR-181a/b expression level increased in PDGF-treated (10 ng/mL) HAoSMCs for 48 h.
The expression level of miR-181a/b in HAoSMCs was measured by quantitative real-time PCR (qRT-PCR), together with biomarkers for differentiated SMCs, miR-143/145, and miR-21 (Elia et al., 2009; Sarkar et al., 2010). The results showed that the expression levels of miR-143/145 and miR-21 were decreased in PDGF-induced HAoSMCs (Fig. 1B). Meanwhile, the expression levels of miR-181a and miR-181b were obviously increased in proliferated HAoSMCs (Fig. 1B). The expression patterns of miR-181c and miR-181d were also detected under PDGF treatment. Similar with the results in carotid artery cells (Li et al., 2015), their expression levels were unchangeable.
miR-181a/b regulates the expression of phenotypic marker genes in HAoSMCs
The modulation between contractile phenotype and synthetic phenotype of smooth muscle cells is associated with changes in proliferation ability, migration capability, and expression level of SMC marker genes (Rensen et al., 2007). To reveal the role of miR-181a/b in expression regulation of SMC marker genes, we transiently transfected HAoSMCs with miR-181a/b mimics or inhibitors for 72 h (sequences listed in Table 1). The expression levels of marker genes in SMC were examined by qRT-PCR with GAPDH as the internal reference. The results showed that miR-181a/b downregulated the expressions of contractile phenotype marker genes (Fig. 2A) while it upregulated those of synthetic phenotype marker genes (Fig. 2B). In contrast, the inhibition of miR-181a/b upregulated the expressions of contractile phenotype marker genes (Fig. 2C) but downregulated those of synthetic phenotype marker genes (Fig. 2D).
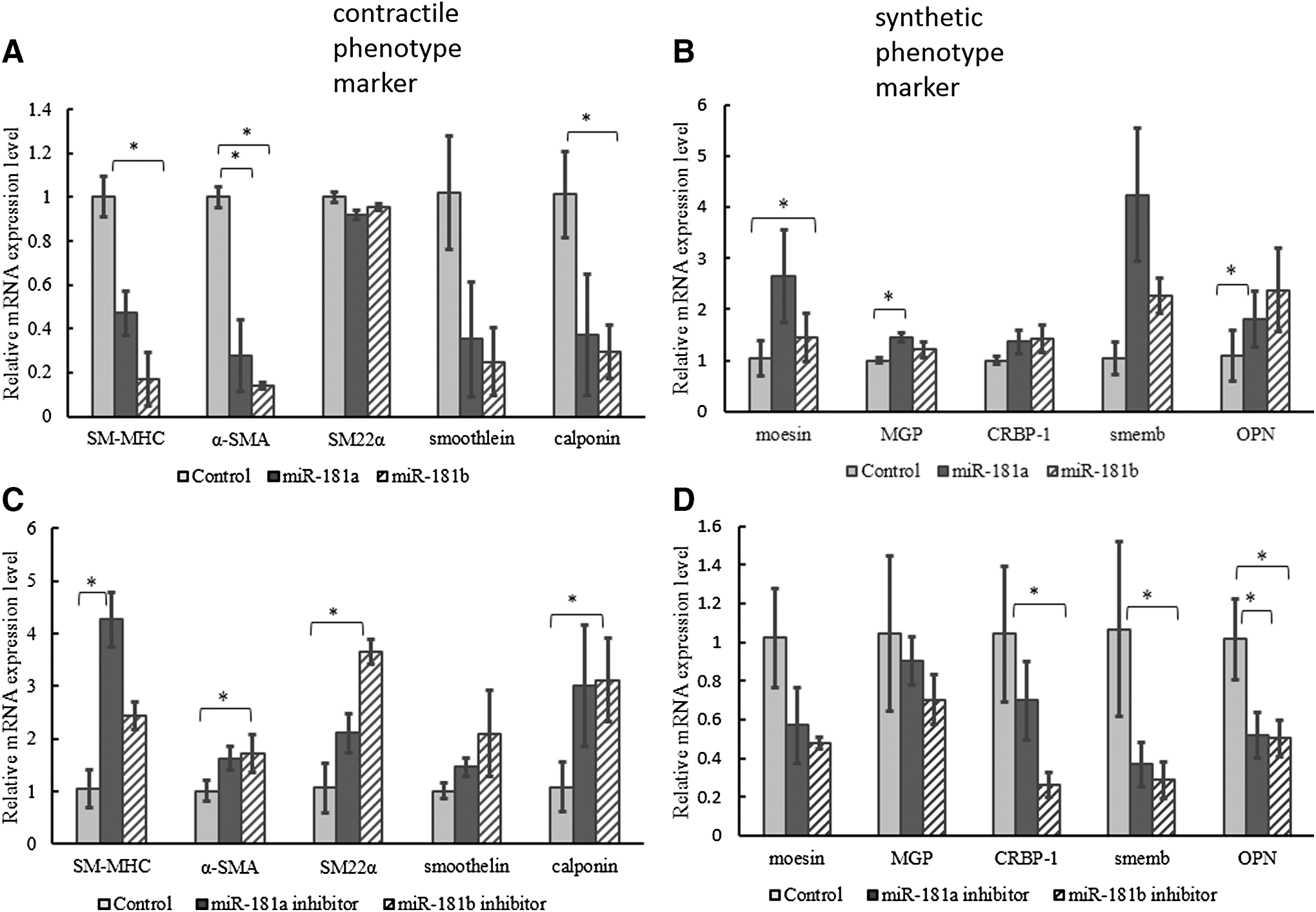
miR-181a/b regulates expressions of HAoSMCs phenotype marker genes. Overexpression of miR-181a/b downregulated expressions of contractile phenotype marker genes (SM-MHC, α-SMA, SM22α, smoothelin, calponin)
miR-181a/b promotes proliferation and migration of HAoSMCs
To determine the effect of miR-181a/b on the proliferation and migration in VSMCs, we transfected HAoSMCs with miR-181a/b mimics or inhibitors for 72 h. The proliferation and migration abilities of HAoSMCs were measured by CCK-8 assay and Transwell assay, respectively. The result of CCK-8 assay showed that the proliferation of HAoSMCs was promoted by miR-181a/b (Fig. 3A) compared to the negative control. In contrast, the proliferation of HAoSMCs was inhibited by miR-181a/b inhibitors (Fig. 3B). The Transwell assay showed that the migration of HAoSMCs was significantly promoted by overexpression of miR-181a/b and suppressed by the inhibition of miR-181a/b (Fig. 3C).
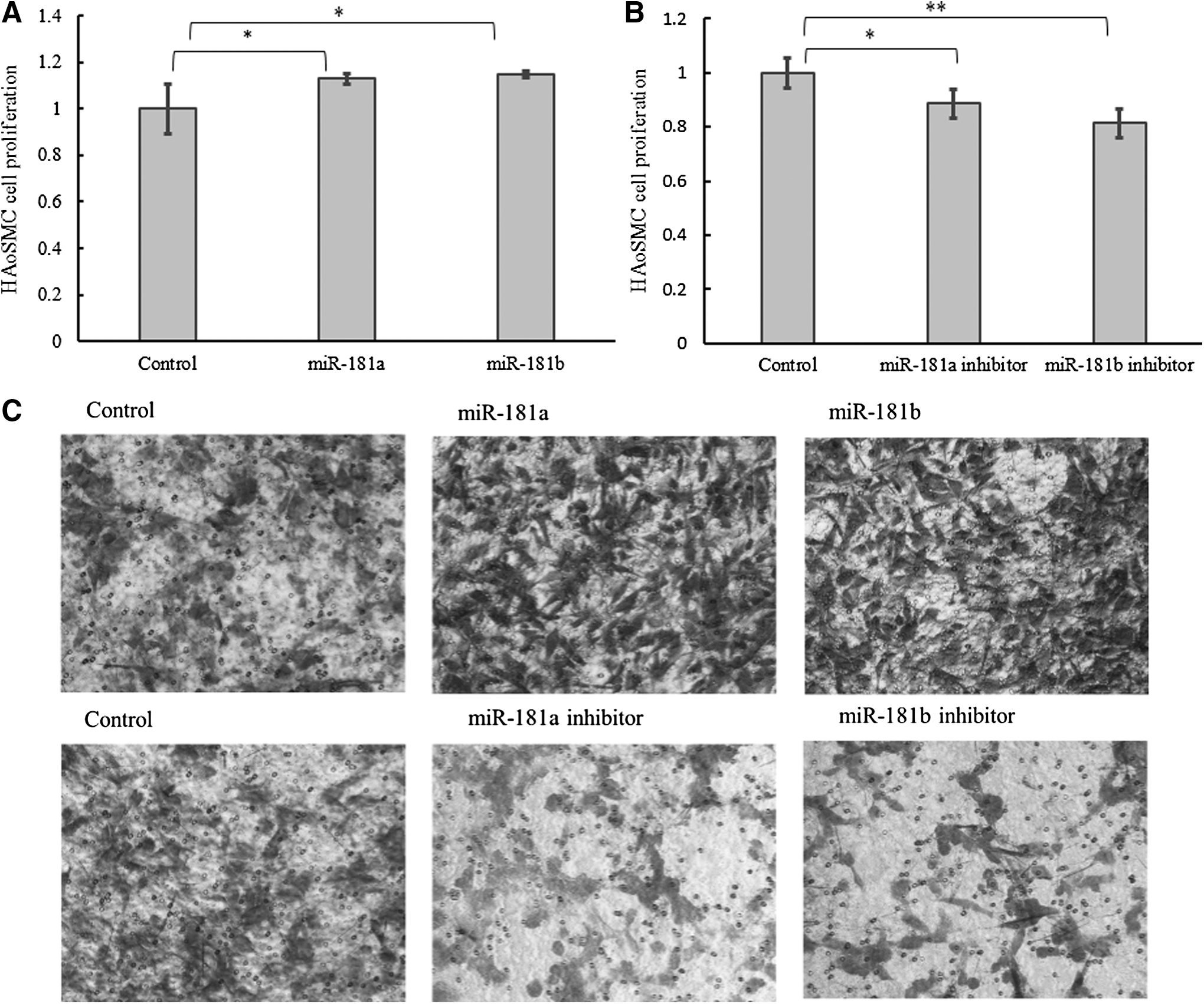
Role of miR-181a/b in HAoSMCs proliferation and migration. After transfection with miR-181a/b mimics or inhibitor for 72 h, HAoSMCs proliferation was detected with a CCK-8 kit. HAoSMCs proliferation was increased by miR-181a/b
miR-181a/b directly targets at 3′ UTR of SRF in HAoSMCs
Through miRNA target prediction, we found that the 3′ UTR of SRF gene has a complementary sequence of miR-181a/b, which indicated that SRF is the possible target of miRNA-181a/b. To investigate whether miR-181a/b could directly target at 3′ UTR of SRF, the dual-luciferase reporter assay was performed by cotransfecting miR-181a or miR-181b with SRF-pGL3 into HAoSMCs for 24 h. The results showed that miR-181a/b clearly suppressed the luciferase activity of the SRF-pGL3 vector (Fig. 4A, B), suggesting that miR-181a/b could directly target on the 3′ UTR of SRF.

miR-181a/b directly binds to and downregulates SRF level. Luciferase assay in HAoSMCs transfected with miR-181a/b mimics and SRF-pGL3 vector
To further confirm whether miR-181a/b could regulate SRF expression, we transfected the HAoSMCs with miR-181a/b mimics or inhibitors for 72 h. The mRNA and protein levels of SRF were measured by qRT-PCR and Western blot, respectively. The qRT-PCR results showed that the mRNA concentration of SRF was reduced in the miR-181a/b overexpression group and increased in the miR-181b inhibition group (Fig. 4C, D). In Western blot experiment, results showed that the protein level of SRF was suppressed by miR-181a/b mimics (Fig. 4E) and enhanced by miR-181a/b inhibitors (Fig. 4F).
SRF regulates the expression of marker genes in HAoSMCs
To examine the effect of SRF on the marker genes in VSMCs during phenotypic transformation, siSRF was designed (F: 5′-GCAAGGCACUGAUUCAGACTT-3′; R: 5′-GUCUGAAUCAGUGCCUUGCTT-3′) and transfected into HAoSMCs. The results indicated that SRF was knocked down with siSRF significantly in HAoSMCs (Fig. 5A, B), and the expression levels of SMC marker genes were tested by qRT-PCR. The expression levels of contractile phenotype genes were decreased (Fig. 5C), while those of synthetic phenotype markers were increased by the weakening of SRF in HAoSMCs (Fig. 5D). These results are similar to overexpression of miR-181a/b (Fig. 2A, B).
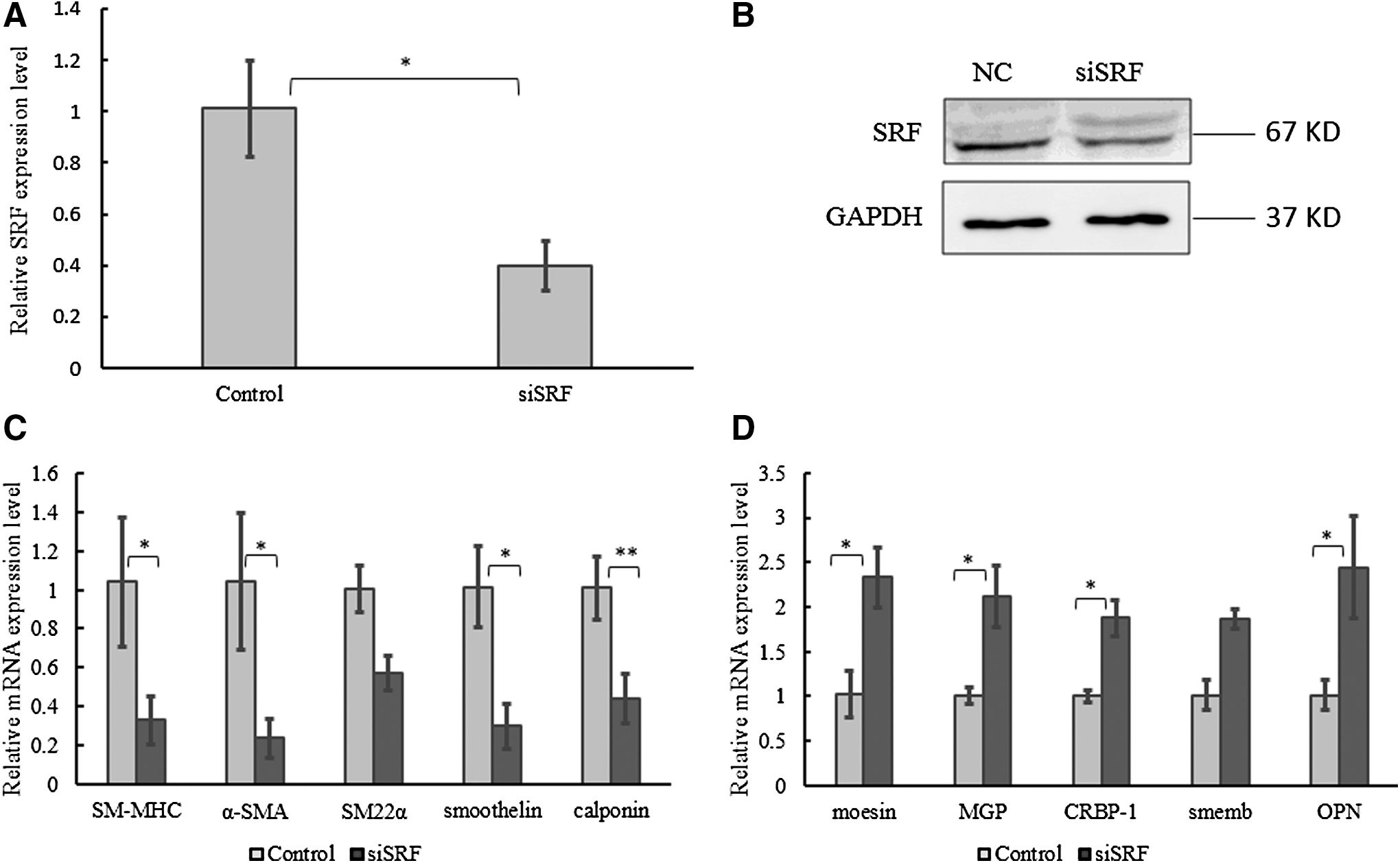
Knockdown of SRF regulates expressions of HAoSMCs phenotype marker genes. The mRNA
Discussion
SMCs possess remarkable phenotypic plasticity that allows rapid adaptation to vascular injury or the development and progression of cardiovascular diseases, including atherosclerosis and restenosis (Orr et al., 2010; Alexander and Owens, 2012). The expression of SMC contractile genes depends on a cis-acting DNA sequence called CArG box (CC(A/T)6GG), which is the binding site for SRF (Owens, 2007). SRF is an essential transcription factor that can toggle between programs of SMC differentiation and growth depending on local environmental cues. Thus, the expression level of SRF has an important influence on SMC differentiation (Miano, 2003). In this study, we found that miR-181a/b could directly target at SRF and be involved in the regulation of the phenotypic transformation in VSMCs (Fig. 6).

Schematic diagram of miR-181a/b regulates SMC phenotype through SRF. miR-181a/b facilitates the conversion of cell to synthetic phenotype through the downregulation of SRF. miR-181a and miR-181b activate the marker genes of synthetic phenotype (Moesin, MGP, CRBP-1, SMemb, OPN), while inhibit the marker genes of contractile phenotype (SM-MHC, α-SMA, SM22α, smoothelin, calponin). SRF is an important factor that regulates SMC from synthetic to contractile phenotype. Myocardin is one of the coactivators of SRF, which is to activate SRF. PDGF activates miR-181a/b at the time of the inhibition of miR-21, miR-143, and miR-145. The full line represents the current results, and the dotted line indicates data reported by other research.
miR-181a and miR-181b are members of the miR-181 family (miR-181a, miR-181b, miR-181c, and miR-181d) and cluster together on the same genomic locations. Members of miR-181 family have distinct gene targets and pathways (Sun et al., 2014), although their seed sequences are similar. miR-181 family is involved in the oncogenic process of acute myeloid leukemia (Su et al., 2015), metabolic regulation in the immune system (Williams et al., 2013), vascular inflammation (Sun et al., 2014), and lymphocyte development and homeostasis (Henao-Mejia et al., 2013). miR-181a plays a significant role in regulation of endothelial cell proliferation by targeting at the gene of nicotinamide adenine dinucleotide phosphate oxidase 4 (NOX4) (Staszel et al., 2011). miR-181a also participates in atherosclerosis through modulation of osteopontin (OPN, an SMC-restricted protein found in abundance in atherosclerotic plaques) (Remus et al., 2013). miR-181b serves as a potent vascular inflammation regulator of NF-κB signaling by targeting at importin-α3 (Sun et al., 2012). miR-181c is a negative regulator that modulates the activation of CD4 (+) T cells (Xue et al., 2011), and miR-181d regulates the acute stress response in thymocytes (Belkaya et al., 2011). However, the effect of miR-181a/b on the phenotypic regulation of SMC was not clear (Li et al., 2015; Lin et al., 2016). In this study, expression level of miR-181a/b is obviously increased in PDGF-induced HAoSMCs (Fig. 1B), suggesting that miR-181a/b might participate in SMCs proliferation and be involved in the regulation of downstream reactions.
To evaluate whether miR-181a/b plays a role in phenotypic transformation of HAoSMCs, we detected the expression levels of SMC marker genes in miR-181a/b-overexpressed or miR-181a/b-inhibited HAoSMCs, respectively. The results showed that most of the expression levels of contractile phenotype marker genes (SM-MHC, α-SMA, SM22α, smoothelin, calponin) were downregulated and that of synthetic marker genes (Moesin, MGP, CRBP-1, SMemb, OPN) upregulated by miR-181a/b. Inhibition of miR-181a/b regulated the expressions of marker genes in the opposite way. Contractile gene SM22α is the only exception in Figure 2A. No obvious reduction of its mRNA was observed when miR-181a/b was overexpressed. Possible reason was that the SRF was not the only activator for SM22α. There are other factors in VSMC that influence the expression of SM22α. These data indicated that miR-181a/b could promote phenotypic transformation to a synthetic phenotype in VSMCs.
During vessel remodeling in physiological conditions, SMCs increase proliferation and migration (Rensen et al., 2007). Then, the role of miR-181a/b in SMC proliferation and migration was clarified by CCK-8 assay and Transwell assay, respectively (Fig. 3). Thus, miR-181a/b appears to be involved in the regulation of the proliferation and migration of HAoSMCs.
Extensive studies have established the crucial role of SRF in controlling SMC-specific gene expression (Miano, 2003). One of the coactivators of SRF is myocardin, which contacts SRF through direct protein–protein interaction (Wang and Olson, 2004). Interestingly, myocardin was found to be activated by NOX4 in mouse embryonic stem cells (Xiao et al., 2009). Regarding that NOX4 is one of the targets of miR-181a in endothelial cell (Staszel et al., 2011), the level of myocardin is supposed to decrease in SMCs when miR-181a is overexpressed; meanwhile, the cellular phenotype switches from contractile to synthetic. In this study, SRF is the target of miR-181a/b predicted by Targetscan software. Moreover, the relationship between miR-181a/b and SRF is conserved in mammals (Fig. 7). The function of SRF in expressions of SMC phenotype genes was confirmed by the knockdown of SRF. Weakened SRF indeed increased the expressions of synthetic marker genes and decreased the expressions of contractile marker genes (Fig. 5C, D). In smooth muscle cells, the knockdown effect of SRF on the expression of phenotypic marker genes is the opposite to that of miR-181a/b. To further identify whether miR-181a/b could regulate SRF, we found that miR-181a/b could target on the 3′ UTR of SRF and regulate its mRNA and protein expression through a series of experiments. In conclusion, our study presented the evidence that miR-181a/b regulated vascular smooth muscle cells from contractile phenotype to synthetic phenotype through targeting on the 3′ UTR of SRF.
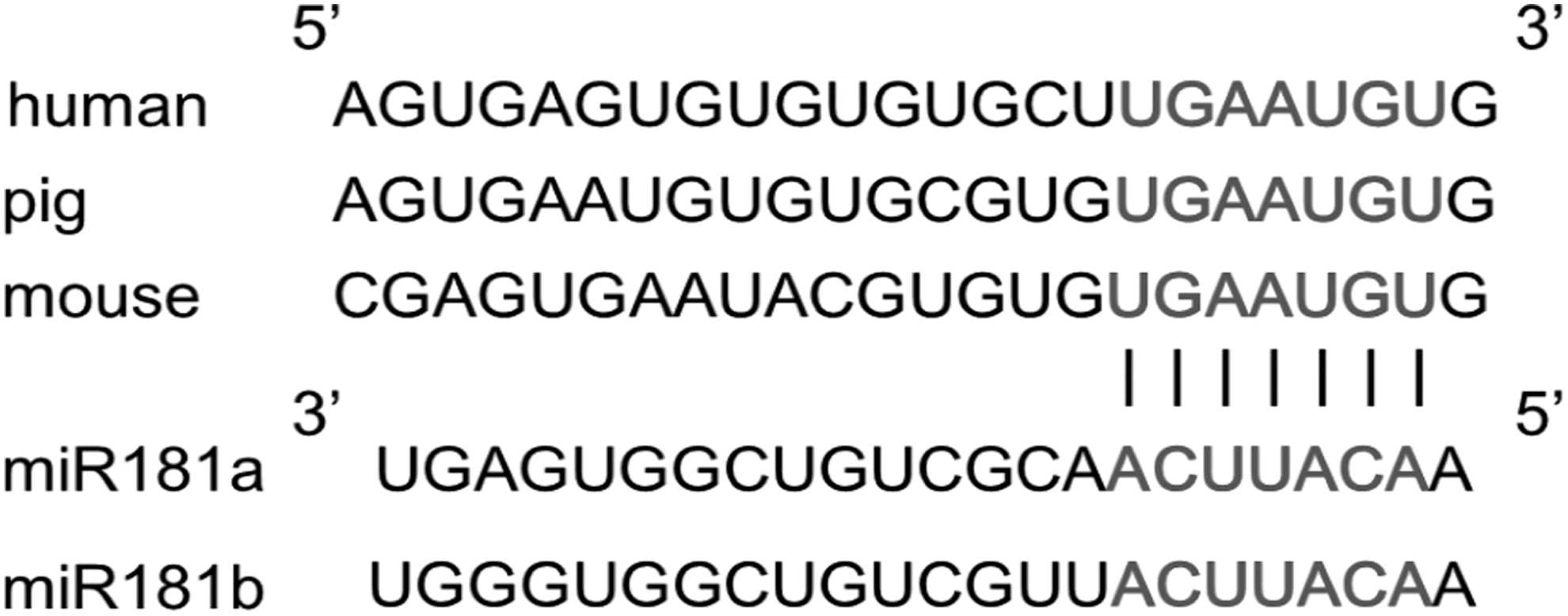
The target sites of miR-181a/b and SRF are conserved in mammals.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China [31260015], the Natural Science Foundation of Qinghai Province [2016-ZJ-746], and the State Key Laboratory for Agrobiotechnology of China Agricultural University [2012SKLAB06-5, 2014SKLAB06-10].
Disclosure Statement
No competing financial interests exist.