
Abstract
Sex-determining region Y-box 2 (SOX2) is an oncogene known to be amplified and overexpressed in various human malignancies, including lung squamous cell carcinoma (SCC). However, the role played by SOX2 in lung SCC development remains to be elucidated. We measured the levels of SOX2 and cyclin D1 mRNA and protein expression in lung SCC tissues and a lung SCC cell line, and found that both levels were dramatically upregulated in specimens of lung SCC tissue when compared with their expression levels in samples of adjacent nonneoplastic tissue. The lung SCC cell line also showed higher levels of SOX2 and cyclin D1 expression than a normal human bronchial epithelium cell line. After using RNA interference to knock down SOX2 expression in NCI-H520 lung SCC cells, their proliferation was reduced. Furthermore, overexpression of SOX2 promoted the proliferation of normal human bronchial epithelium cells. To further determine whether cyclin D1 was downstream target gene of SOX2, we measured the levels of cyclin D1 expression that occurred when SOX2 was knocked down or overexpressed. SOX2 knockdown significantly decreased the levels of cyclin D1 mRNA and protein expression, while SOX2 overexpression upregulated the levels of cyclin D1. We used bioinformatics data to identify potential cyclin D1 promoter binding sites for SOX2. Results of luciferase reporter assays, electrophoretic mobility shift assays, and chromatin immunoprecipitation assays confirmed that cyclin D1 was a direct target of transcription factor SOX2 in human lung SCC cells.
Introduction
L
The embryonic stem cell transcription factors SOX2 and cyclin D1 have recently received increased attention by researchers. The SOX2 gene is located on chromosome 3q26.3–q27, and encodes a transcription factor with three main domains: the N-terminal domain, HMG domain, and the transactivation domain (Boyer et al., 2005; Bass et al., 2009). These studies on SOX2 have primarily focused on its crucial roles in regulation of pluripotency in embryonic stem cells, lineage fate determinant, maintenance of homoeostasis in tracheobronchial epithelium, and reprogramming somatic cells back toward pluripotency (Adameyko et al., 2012). SOX2 is amplified and overexpressed in several human cancers, including lung cancer, glioma, breast cancer, pancreatic carcinoma, esophageal carcinoma, and ovarian carcinoma (Bass et al., 2009; Herreros-Villanueva et al., 2013; Zhang et al., 2013; Tian et al., 2014). Furthermore, SOX2 is more highly expressed in lung cancer tissue than in normal lung tissue and other tumor. In addition, the SOX2 gene is amplified to a greater extent in lung SCC than in lung adenocarcinoma tissue (Lu et al., 2010; Yuan et al., 2010). Accumulated evidence indicates that in cancer cells, SOX2 is involved in cellular proliferation, invasion, migration, self-renewal, and the maintenance of “steminess.” Although SOX2 is an oncogenic transcription factor, the downstream target genes of SOX2 in lung SCC cells remain to be identified.
In this study, we measured the levels of SOX2 expression in lung SCC tissue and an SCC cell line, and found that both SOX2 mRNA and protein levels were dramatically higher in SCC tissue when compared with their levels in adjacent nonneoplastic tissue. Furthermore, the lung SCC cell line had higher levels of SOX2 and cyclin D1 than a normal human bronchial epithelium cell line. We also investigated the role of SOX2 expression in the proliferation of SCC cells and normal human bronchial epithelial cells. The SOX2 knockdown inhibited the proliferation of lung SCC cells, while the SOX2 overexpression promoted the proliferation of normal human bronchial epithelial cells.
Cyclin D1 is a key regulator of the G0–G1 transition that occurs during cycling of both normal and cancer cells, and is thought to be a proto-oncogene involved in the initiation and progression of carcinoma (Quelle et al., 1993; Yu et al., 2001). In this study, we found that the levels of cyclin D1 mRNA and protein were both dramatically higher in samples of lung SCC tissue when compared with samples of adjacent nonneoplastic tissue. We also confirmed that SOX2 is involved in the proliferation of lung SCC cells by regulating the cell cycle. In addition, we investigated whether the cyclin D1 gene might be located downstream of SOX3 and performed a bioinformatics search to identify potential cyclin D1 promoter binding sites for SOX2. Finally, we confirmed that cyclin D1 was a direct target of transcription factor SOX2 in human lung SCC cells.
Materials and Methods
Cells and tissue samples
Human lung SCC cell line NCI-H520 and normal human bronchial epithelium cell line BEAS-B2 were purchased from the American Type Culture Collection (ATCC, Manassas, VA) and cultured in the Roswell Park Memorial Institute (RPMI) medium supplemented with 10% fetal bovine serum. Ten samples of lung SCC tissue and adjacent nonneoplastic tissue were collected from 10 consecutive lung cancer patients at the First People's Hospital in Chenzhou City, China. The study protocol was approved by the Ethics Committee of Chenzhou First Peoples Hospital, and each patient signed a written Informed Consent document indicating their willingness to participate in the study. All of the enrolled patients were undergoing curative surgical resection without receiving chemotherapy or radiation therapy. The samples of bronchial epithelium collected from adjacent nonneoplastic lung tissue were used as control tissues.
CCK-8 assays
A commercial cell counting kit (CCK-8) (Dojindo; Kumamoto, Japan) was used to evaluate cell viability.
BrdU cell proliferation assay
Cells were seeded on cover slips (Fisher Scientific International, Hampton, NH), and then incubated with bromodeoxyuridine (BrdU) for 1 h, after which, they were stained with anti-BrdU antibody (Upstate) (Merck KGaA, Darmstadt, Germany) as described in the manufacturer's instructions. Hoechst staining solution was used to stain cell nuclei. Images were acquired with a confocal microscope (Olympus FV1000; Olympus, Tokyo, Japan).
Cell cycle analysis
Briefly, cells were trypsinized, harvested, washed with cold phosphate-buffered saline, and then fixed in 70% solution overnight at 4°C. On the following day, the cells were stained with RNase A (10 μg/mL) and propidium iodide (20 mg/mL) (Sigma, St Louis, MO) in the dark for 30 min. The cells were then analyzed for their DNA content with an Aria Calibur flow cytometer (BD Biosciences; Franklin Lakes, NJ) and Flowjo 7.6 software (BD Biosciences).
Western blot studies
Cells and tissues were lysed in the lysis buffer, and aliquots of their total proteins were separated on polyacrylamide gels containing 4–12% SDS. Next, the separated protein bands were transferred onto polyvinylidene difluoride membranes (Millipore, Billerica, MA). The membranes were then blocked for 1 h in TBST containing 2% bovine serum albumin; after which, they were incubated with antibodies against SOX2 [1:1000, Cell Signaling Technology (CST); Danvers, MA], cyclin D1 (1:1000, CST), GAPDH (1:1000, CST), and HRP-conjugated goat anti-rabbit secondary antibodies (1:1000; Promab, Richmond, CA).
RNA extraction and quantitative reverse transcription PCR
Total RNA was extracted from cells and tissues using Trizol Reagent (Life Technologies, Carlsbad, CA) following the manufacturer's protocols. RT-qPCR was performed using SYBR green PCR Mix (iTAP; Bio-Rad, Hercules, CA) in conjunction with an Applied Biosystems (ABI) step-one plus sequence detection system (Applied Biosystems, Foster City, CA). Fold changes were calculated using ΔΔCT values and the formula 2−ΔΔCT.
Transient transfection and small interfering RNA
Small interfering RNA (siRNA) with the oligonucleotides 5′-GGT TGA CAC CGT TGG TAAT-3′ and targeting human SOX2 was purchased from RiboBio (Guangzhou, China). Nonspecific oligonucleotides were used as negative controls (NCs). Lipofectamine 2000 (Life Technologies) was used to transfect siRNA into the targeted cells. Following transfection, the cells were harvested at the specified time point and used for further assays.
Luciferase assays
Plasmid pCDNA3.1-SOX2 expressing SOX2 was constructed. The reporter construct was generated by ligating the putative cyclin D1 promoter region and mutations to pGL3-basic (pGL3-cyclinD1-WT and pGL3-cyclinD1-mut reporters). The region containing the putative cyclin D1 promoter mutations (pGL3-cyclinD1-mut reporter) was amplified using the following cloning primers: 5′-TTCTTGGAAATGCGCAATCCAACCCGGCTT-GGATATG-3′ and 5′-CATATCCAAGCCGGGTTGGATTGCGCATTTCCAAGAA-CTCTCA-3′. Plasmids pcDNA3.1 and pGL3-basic were used as controls. All vectors were sequenced to verify the presence of the correct insert. NCI-H520 cells cultured in 12-well plates were transfected using Lipofectamine 3000 (Life Technologies). After 48 h of transfection, the cells were lysed and assayed for the luciferase activity. Transfections performed for luciferase assays were carried at least four times, and the results were measured in duplicate. All measurements of luciferase activity were performed using a Promega Turner Biosystems Modulus Multimode Reader luminometer. The resultant data were analyzed using the two-way ANOVA test.
Immunofluorescence staining
NCI-H520 and BEAS-B2 cells were grown on round cover slips and processed for immunofluorescence staining as previously described (Khromov et al., 2011). SOX2 and cyclin D1 were detected by using a polyclonal antibody against SOX2 and cyclin D1 (CST). The secondary antibodies were conjugated with Alexa Fluorescence 568 and DAPI (1:1000; Sigma). The stained cells were visualized using an Olympus BX60 fluorescent microscope.
Electrophoretic mobility shift assays
Electrophoretic mobility shift assays (EMSAs) were performed using 5′-Cy5-labeled cognate double-stranded DNA elements containing the proper motif sequence. The nuclear extracts were prepared and EMSAs were performed as previously described (Li et al., 2013; Mistri et al., 2015). The following probes were used in the assays: (1) probe containing the Sox2 binding site sequence: 5′-TCGGGCAGCCATTGTGATGCATAT-3′ and (2) mutant probe sequence: 5′-TCGGGCAGC
Chromatin immunoprecipitation
Chromatin immunoprecipitation (ChIP) assays were performed as previously described (Turner et al., 2006; Carey et al., 2009) using SOX2 (CST) and a matched IgG antibody (CST). The immunoprecipitated samples were analyzed by RT-qPCR, and the results were normalized using the percent input method. The Ct value of each chromatin immunoprecipitated DNA fraction was normalized to the Ct value of the input DNA fraction. The percent input (% input) was calculated for each ChIP fraction. The cyclin D1 primers used were as follows: forward, 5′-TGCCGGGCTTTGATCTTT-3′; reverse, 5′-CGGTCGTTGAGGAGGTTGG-3′.
Statistical analysis
All results are presented as the mean ± standard error of the mean (SEM) of at least three independent experiments. Student's t-test was used to assess differences between two groups. A p value <0.05 was considered statistically significant.
Results
SOX2 and cyclin D1 were overexpressed in lung SCC tissues and cell lines
We measured the levels of SOX2 and cyclin D1 expression in samples of lung SCC tissue and adjacent nonneoplastic tissue. As shown in Figure 1A and B, the levels of SOX2 and cyclin D1 mRNA were dramatically higher in lung SCC tissue when compared with their levels in adjacent nonneoplastic tissue. In addition, Western blot results showed that both SOX2 and cyclin D1 proteins were overexpressed in lung SCC tissues (Fig. 1C, E). We also examined SOX2 and cyclin D1 expression in a lung SCC cell line (NCI-H520) and a normal human bronchial epithelium cell line (BEAS-2B), and found that the lung SCC cell line showed higher levels of SOX2 and cyclin D1 expression than the normal human bronchial epithelium cell line (Fig. 1F, G). Moreover, amplification of SOX2 and cyclin D1 in the lung SCC cell line (NCI-H520) was further confirmed by immunofluorescence (Fig. 1H, I).
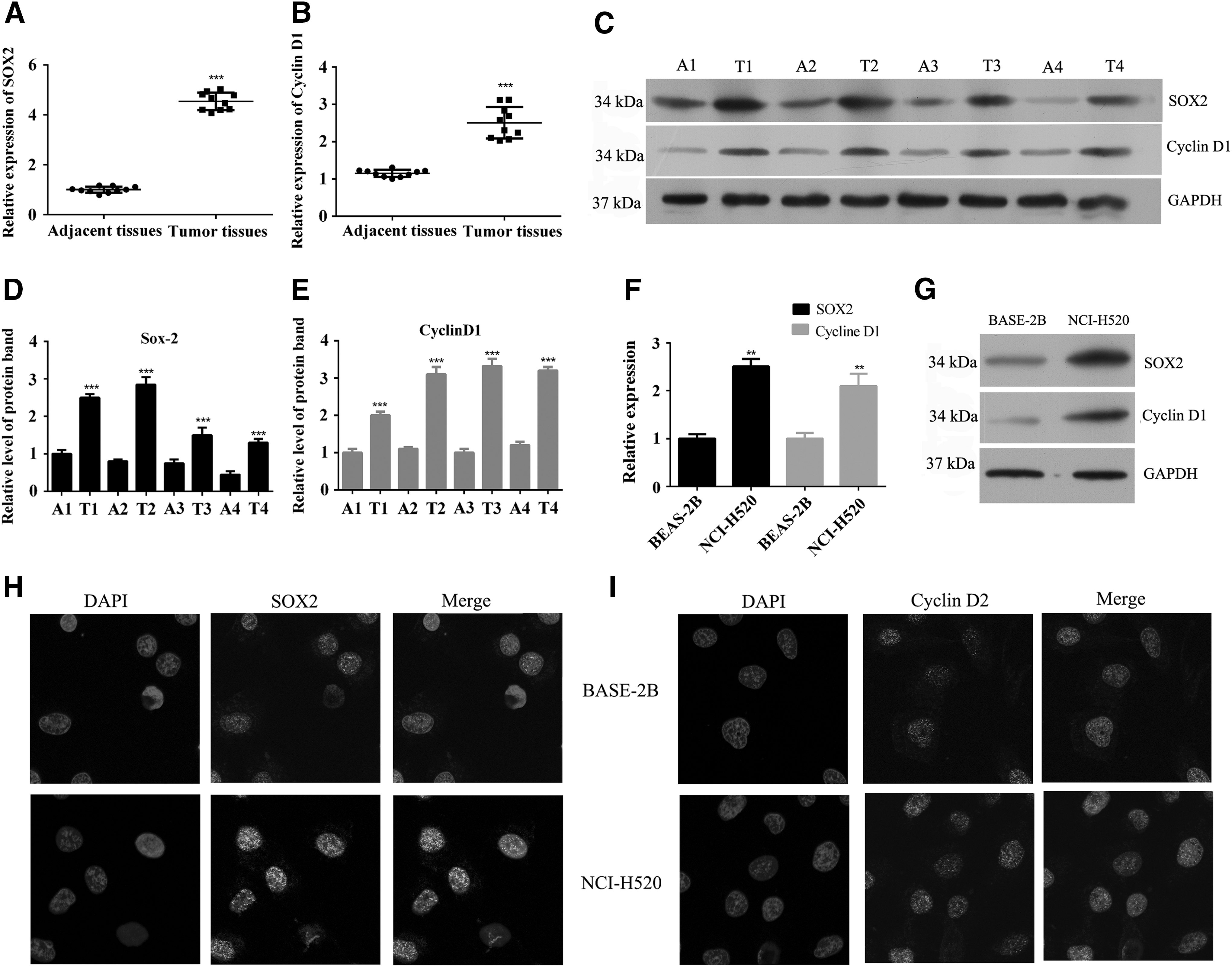
SOX2 and cyclin D1 were overexpressed in lung SCC tissues and cell lines.
Knockdown of SOX2 inhibited the proliferation of lung SCC cells
To further study the role of SOX2 in lung SCC cell proliferation, we performed SOX2-silencing transfection studies using NCI-H520 cells. After transfection for 48 h, the levels of SOX2 mRNA and protein were detected to determine transfection efficiency (Fig. 2A–C). Cell viability was assessed after transfection, and found to be decreased by SOX2 knockdown in both NCI-H520 and BEAS-2B cells (Fig. 2D). However, the SOX2 knockdown produced no effect on the rates of apoptosis in NCI-H520 and BEAS-2B cells (Supplementary Fig. S1A; Supplementary Data are available online at

Knockdown of SOX2 inhibited the proliferation of lung SCC cells.
SOX2 overexpression promoted the proliferation of normal human bronchial epithelial cells
As SOX2 expression in BEAS-2B cells was lower than that in NCI-H520 cells, we chose BEAS-2B cells for use in performing further studies on cell proliferation. SOX2 was highly expressed in BEAS-2B cells after they had been transfected with pcDNA3.1-SOX2 plasmid for 48 h (Fig. 3A). As shown in Figure 3B and C, the SOX2 protein was also more highly expressed in BEAS-2B cells transfected with pcDNA3.1-SOX2 plasmid than in cells transfected with the empty vector. The BrdU proliferation assay revealed that overexpression of SOX2 significantly increased the numbers of BrdU-positive cells, suggesting that SOX2 overexpression had promoted BEAS-2B cell proliferation (Fig. 3D). In addition, we found that SOX2 overexpression decreased the proportion of cells in G1 phase and increased the proportion of cells in S phase (Fig. 3E). Our studies also showed that knockdown of SOX2 inhibited the migration/invasion capabilities of NCI-H520 cells, while SOX2 overexpression increased those capabilities (Supplementary Fig. 1C, D).
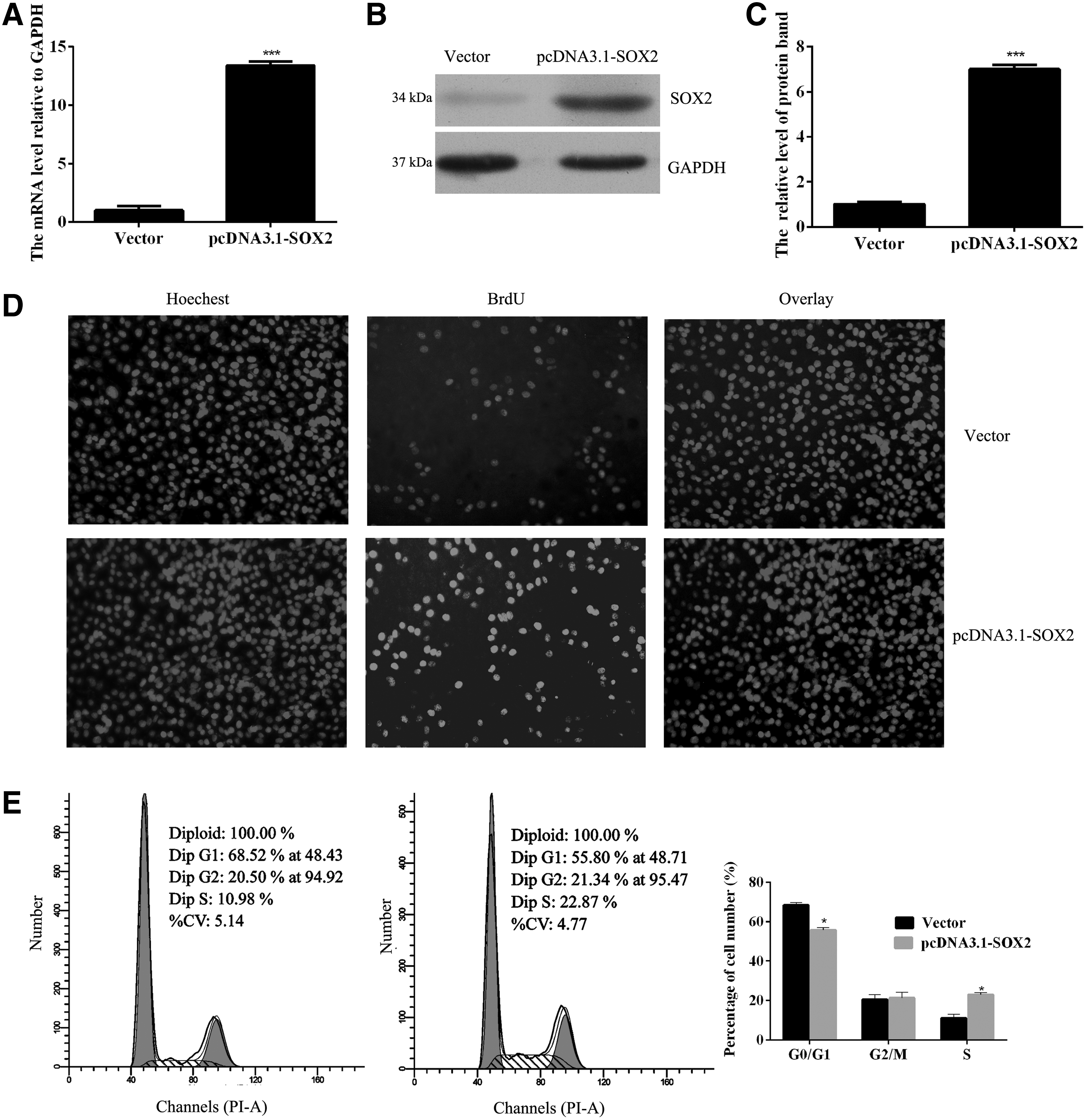
SOX2 overexpression promoted the proliferation of lung SCC cells.
Cyclin D1 was a direct target of transcription factor SOX2 in human lung SCC cells
We confirmed that SOX2 was involved in the proliferation of lung SCC cells through its ability to regulate the cell cycle. As a key regulator of the G1 progression step within the cell cycle, cyclin D1 plays a pivotal role in the lung SCC cell proliferation process. To further examine whether cyclin D1 might be a downstream target gene for SOX2, transfections using siRNA-SOX2 and pcDNA3.1-SOX2 plasmids were performed. The SOX2 knockdown significantly decreased the levels of cyclin D1 mRNA, while SOX2 overexpression upregulated the cyclin D1 mRNA levels (Fig. 4A, B). Moreover, similar results were found when investigating the corresponding changes in cyclin D1 protein expression, Figure 4C–E. We used the Jaspar database (
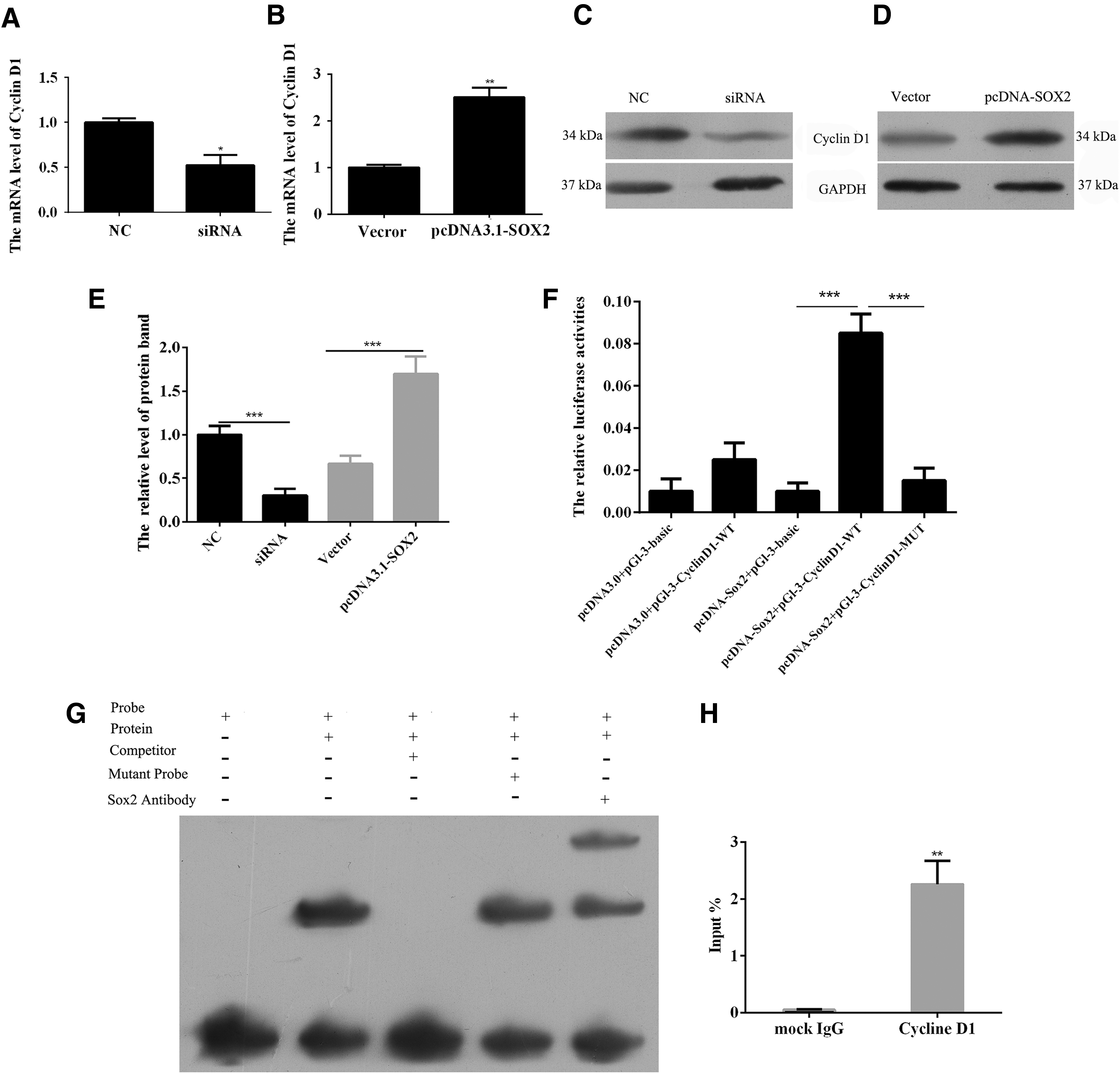
Cyclin D1 is a downstream target gene for transcription factor SOX2.
Discussion
As Sox2 is highly expressed in conjunction with cyclin D1 in most human lung SCC cells, the downstream targets of SOX2 have been widely investigated. As a transcription factor, SOX2 exerts a tumorigenic effect by regulating several downstream genes in different types of cancers. SOX2 amplifications and overexpression are generally associated with tumor progression in the esophagus, lungs, retina, skin, and pituitary. Tian et al. (2014) showed that SOX2 activated AKT signaling and thereby promoted gastric cancer progression. It was reported that the tumorigenic effect of SOX2 in lung SCC cells is partially mediated by suppression of CDKN1A (Fukazawa et al., 2016). However, the role of Sox2 in gastric cancer is still controversial. SOX2 was progressively downregulated during gastric cancer development and inhibited cell proliferation and metastasis by regulating PTEN (Wang et al., 2015). Otsubo et al. (2008) reported that SOX2 played important roles in growth inhibition through cell cycle arrest and apoptosis in gastric epithelial cells. Recently, Sox2 was reported as a tumor suppressor by modulating Wnt-related and intestinal genes (Sarkar et al., 2016). Our data, as well as data from other investigators, clearly indicate that inhibition of SOX2 suppresses lung SCC cell growth in vitro. Even though SOX2 is recognized as a lineage-specific oncogenic transcription factor in lung cells, the downstream genes that SOX2 targets in lung SCC cells have not been identified. These results indicate that Sox2's effect on cell proliferation and tumorigenesis is highly context dependent.
Previous studies reported that upregulated SOX2 expression is associated with a poor clinical outcome among lung SCC patients (Sholl et al., 2010). In this study, we found dramatically upregulated levels of SOX2 and cyclin D1 mRNA and protein expression in lung SCC tissue when compared with those levels in samples of adjacent nonneoplastic tissue. Moreover, similar results were found when examining SOX2 and cyclin D1 expression in lung SCC and normal human bronchial epithelium cell lines.
To understand the role of SOX2 in lung SCC and normal human bronchial epithelium cells, we knocked down SOX2 expression in lung SCC cells and induced its overexpression in normal human bronchial epithelium cells. The knockdown of SOX2 inhibited the proliferation of lung SCC cells, while SOX2 overexpression promoted the proliferation of normal human bronchial epithelium cells. Downregulation of SOX2 was previously shown to inhibit proliferation and induce apoptosis in two nonsmall cell lung cancer cell lines (A549 and H460) (Chen et al., 2012). Other studies have focused on the downstream pathways of SOX2 in lung cancer. For example, SOX2 knockdown was shown to induce apoptosis in NSCLC cells by activating the MAP4K4 pathway (Chen et al., 2014). Another study reported that the knockdown of SOX2 inhibited the viability and colony formation of lung SCC cell lines H520 and H226. SOX2 was also reported to sustain the proliferation of lung SCC cells by suppressing of CDKN1A (Fukazawa et al., 2016). Our results are consistent with those in previous studies. SOX2 is highly expressed in isolated cancer stem-like cells and helps to maintain their “stemness” by regulating complex transcriptional networks. Numerous studies have revealed that SOX2 regulates the transcriptional networks of various oncogenes, including WNT1, WNT2, NOTCH1, and c-MYC, and promotes tumor formation by human lung cancer cells (Chen et al., 2012).
Cyclin D1 is an important regulator of the cell cycle and is overexpressed in a large number of human cancers, including lung cancer. Moreover, cyclin D1 is suspected to play a pivotal role in processes involved in carcinogenesis and cancer progression. Cyclin D1 is amplified in both NSCLC and preinvasive bronchial lesions; generally by one parental allele. Furthermore, cyclin D1 expression is highly associated with the clinical outcomes of lung cancer patients (Tsuruta et al., 1993; Zhang et al., 1993; Shinozaki et al., 1996).
We hypothesized that SOX2 might promote the proliferation of lung SCC cells and normal human bronchial epithelial cells by activating a downstream target. In our study, the knockdown of SOX2 significantly decreased cyclin D1 expression, while SOX2 overexpression upregulated cyclin D1 levels. We searched the Jaspar database to identify potential binding sites for SOX2 in the cyclin D1 promoter region, and then performed luciferase reporter assays and EMSA combined with ChIP assays to confirm that cyclin D1 was a direct target for transcription factor SOX2 in human lung SCC cells. Previous studies showed that cyclin D1 was downregulated at the transcription level in a SOX2-dependent manner in human glioma stomal cells (Xu et al., 2016). Sox2 was also shown to enhance the proliferation of cervical squamous cancer cells by upregulating cyclin D1 expression (Ji et al., 2014).
In summary, we confirmed that both SOX2 and cyclin D1 are highly expressed in lung SCC tissue and lung SCC cells. In addition, we showed that SOX2 promotes the proliferation of lung SCC and normal human bronchial epithelium cells. Finally, we provided evidence that cyclin D1 is a direct target of transcription factor SOX2 in human lung SCC cells.
Footnotes
Acknowledgment
This work was supported by the Chenzhou First People's Hospital.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.