
Abstract
Treatment options for chronic hepatitis B (CHB) infection are extremely limited. CXCR5+ CD8+ T cell is a novel cell subtype and could possess strong cytotoxic properties in HIV infection. In this study, we investigated the role of CXCR5+ CD8+ T cells in CHB patients. Compared to healthy individuals, both CHB patients and hepatitis B virus (HBV)-infected hepatocellular carcinoma patients presented significant upregulation of CXCR5+ CD8+ T cells in peripheral blood, in which CXCR5+ CD8+ T cells were negatively correlated with the frequency of CXCR5+ CD4+ T cells in CHB patients. After PMA+ionomycin stimulation, CXCR5+ CD8+ T cells from CHB patients presented significantly higher transcription level of interferon gamma (IFN-γ), interleukin 10 (IL-10), and IL-21, as well as higher IL-10 and IL-21 protein secretion, than CXCR5− CD8+ T cells. Unlike CXCR5+ CD4+ T cells, when incubated with naive CD19+CD27− B cells, CXCR5+ CD8+ T cells alone did not upregulate IgM, IgG, and IgA secretion. However, addition of CXCR5+ CD8+ T cells in B cell-CXCR5+ CD4+ T cell coculture significantly increased the levels of secreted IgG and IgA, demonstrating that CXCR5+ CD8+ T cell could indirectly offer B cell help. Furthermore, high frequencies of CXCR5+ CD8+ T cells tended to associate with low HBV DNA load, and the frequency of CXCR5+ CD8+ T cells was negatively correlated with alanine aminotransferase (ALT) level. Together, these results suggested that CXCR5+ CD8+ T cells were involved in the antiviral immune responses in CHB and could potentially serve as a therapeutic candidate.
Introduction
D
HBV infection in newborns and adults results in profoundly different outcomes, with lifelong CHB in most newborns and resolved acute infection in adults. This striking difference suggests that the host immune response, which is significantly weaker in neonates, plays a crucial role in the resolution of HBV. A strong T cell response with specificity toward a wider range of antigens is associated with a higher likelihood of HBV clearance, than a weak T cell response with narrowly focused T cell specificity (Liu et al., 2014b). In chimpanzees, it has been shown that CD8+ T cell response during acute HBV infection is required for the clearance of infection, and both cytolytic and noncytolytic processes are involved (Thimme et al., 2003). In humans, HBV persistence is associated with weak or undetectable levels of HBV-specific CD8+ T cells (Jung et al., 1991; Penna et al., 1991). The mechanism through which CD8+ T cells mediate HBV clearance has not been fully elucidated.
Recently, a novel population of CXCR5+ CD8+ T cells is shown to possess antiviral capacity in chronic lymphocytic choriomeningitis virus (LCMV) infection, during which CD8+ T cells generally display exhausted phenotypes (He et al., 2016). It has been shown that this population that was able to migrate to the B cell follicles under the influence of the CXCR5-CXCL13 axis (Quigley et al., 2007) presented lower levels of exhaustion markers PD-1, Tim-3, and KLRG1, exhibited potent cytotoxicity, and demonstrated synergistic therapeutic potential when combined with PD-L1 neutralization (He et al., 2016). In HIV-infected human subjects, the frequency of CXCR5+ CD8+ T cells was inversely correlated with viral load. Therefore, in this study, we examined the presence and function of CXCR5+ CD8+ T cells in CHB patients.
Materials and Methods
Patients
This study recruited 14 CHB patients, 14 HBV-infected HCC patients (HBV-HCC), and 14 uninfected healthy controls. All groups were age and gender matched. All CHB and HCC patients were newly diagnosed at the time of sample collection, and no medication was given. Peripheral blood was harvested from all study participants. Diagnosis was conducted in Kunming General Hospital of PLA. Exclusion criteria included age <18 or >65 years, concurrent hepatitis C virus (HCV) and/or HIV infections, other malignancies, and presence of alcoholic liver disease. The study was approved by the Ethics Committees of Kunming General Hospital of PLA and The First People's Hospital of Yunnan. Written informed consent was obtained from all participants before sampling. Patient information was analyzed by DICAT (Vancouver, Canada). Peripheral blood mononuclear cells (PBMCs) were collected from heparinized blood using the standard Ficoll-Paque (GE Healthcare) centrifugation method. HBV DNA load was measured in the serum using the VERSANT HBV DNA 1.0 Assay (Siemens Healthineers).
FACS assay
For the evaluation of the frequencies of CXCR5+ CD8+ and CD4+ T cells, PBMCs were stained directly ex vivo with anti-human CD3, CD4, CD8, and CXCR5 antibodies (BD Pharmingen), as well as LIVE/DEAD Violet Viability/Vitality Kit (Thermo Fisher Scientific), for 30 min on ice, followed by two washings. Cells were then fixed in 2% formaldehyde and examined in FACSCanto cytometer. For the isolation of naive CD19+CD27− B cells, CXCR5+ CD8+ T cells, CXCR5− CD8+ T cells, and CXCR5+ CD4+ T cells, cells were sorted in FACSAria cytometer immediately after surface staining.
Reverse transcriptase-PCR
CXCR5+ and CXCR5− CD8+ T cells were sorted from unstimulated PBMCs or PBMCs stimulated by 2 ng/mL PMA and 100 ng/mL ionomycin (Sigma Aldrich) for 12 h. Total RNA was extracted from CD8+ T cells using the RNeasy Mini Kit (Qiagen). cDNA was synthesized using the SuperScript III Reverse Transcriptase Kit (Invitrogen). Quantification of the IFN-γ, interleukin 10 (IL-10), and IL-21 transcripts was performed in the iCycler iQ system using SYBR Green (Bio-Rad) as the detection agent. A series of four 10-fold dilutions of the cDNA template was used to generate the standard curve. All cytokine expressions were normalized to the GAPDH expression. Each experiment was performed in triplicates.
Enzyme-linked immunosorbent assay
The Human IFN-γ, IL-10, IL-21, IgM, IgG, and IgA enzyme-linked immunosorbent assay (ELISA) Ready-SET-Go Kits (eBioscience) were used to examine the secreted cytokine and antibody levels following instructions provided by the manufacturer.
T cell-B cell coculture
5 × 104 cells per well per 200 mL sorted CD19+CD27− or CD19+CD27+ B cells, CXCR5+ CD8+ T cells, and/or CXCR5+ CD4+ T cells were cocultured in the presence of 1 μg/mL Staphylococcal enterotoxin B (SEB; Sigma Aldrich) for 12 days, after which the supernatant was harvested for antibody detection.
Statistical analyses
Unpaired t test with Welch's correction was used for comparison between two groups. One-way or two-way analysis of variance (ANOVA) followed by Tukey's posttest was used for comparison between three or more groups. Correlation between two groups was examined by Pearson's correlation test. p Value <0.05 was considered significant.
Results
CXCR5+ CD8+ T cell frequency was elevated in CHB and in HBV-HCC patients
We first examined the frequency of CXCR5+ cells in circulating CD8+ T cells in HBV-uninfected healthy controls, CHB patients, and HBV-HCC patients. The frequency of CXCR5+ CD8+ T cells as a percentage in CD3+ T cells was very low in healthy individuals, but was significantly elevated in CHB and in HBV-HCC patients. In addition, the frequency of CXCR5+ CD8+ T cells in HBV-HCC patients was significantly lower than that in CHB patients (Fig. 1B). Interestingly, we found that the frequency of CXCR5+ CD4+ T cells as a percentage in CD3+ T cells was gradually downregulated from healthy controls, CHB patients, to HBV-HCC patients (Fig. 1C). To investigate whether the upregulation of CXCR5+ CD8+ T cells is to compensate for the loss of CXCR5+ CD4+ T cells in CHB and HBV-HCC patients, we examined the correlation between the frequencies of CXCR5+ CD4+ T cells and CXCR5+ CD8+ T cells in CHB and HBV-HCC patients. In CHB patients, the frequencies of CXCR5+ CD4+ T cells and CXCR5+ CD8+ T cells were inversely correlated (Fig. 1D), while in HBV-HCC patients, no such correlation was observed (Fig. 1E).
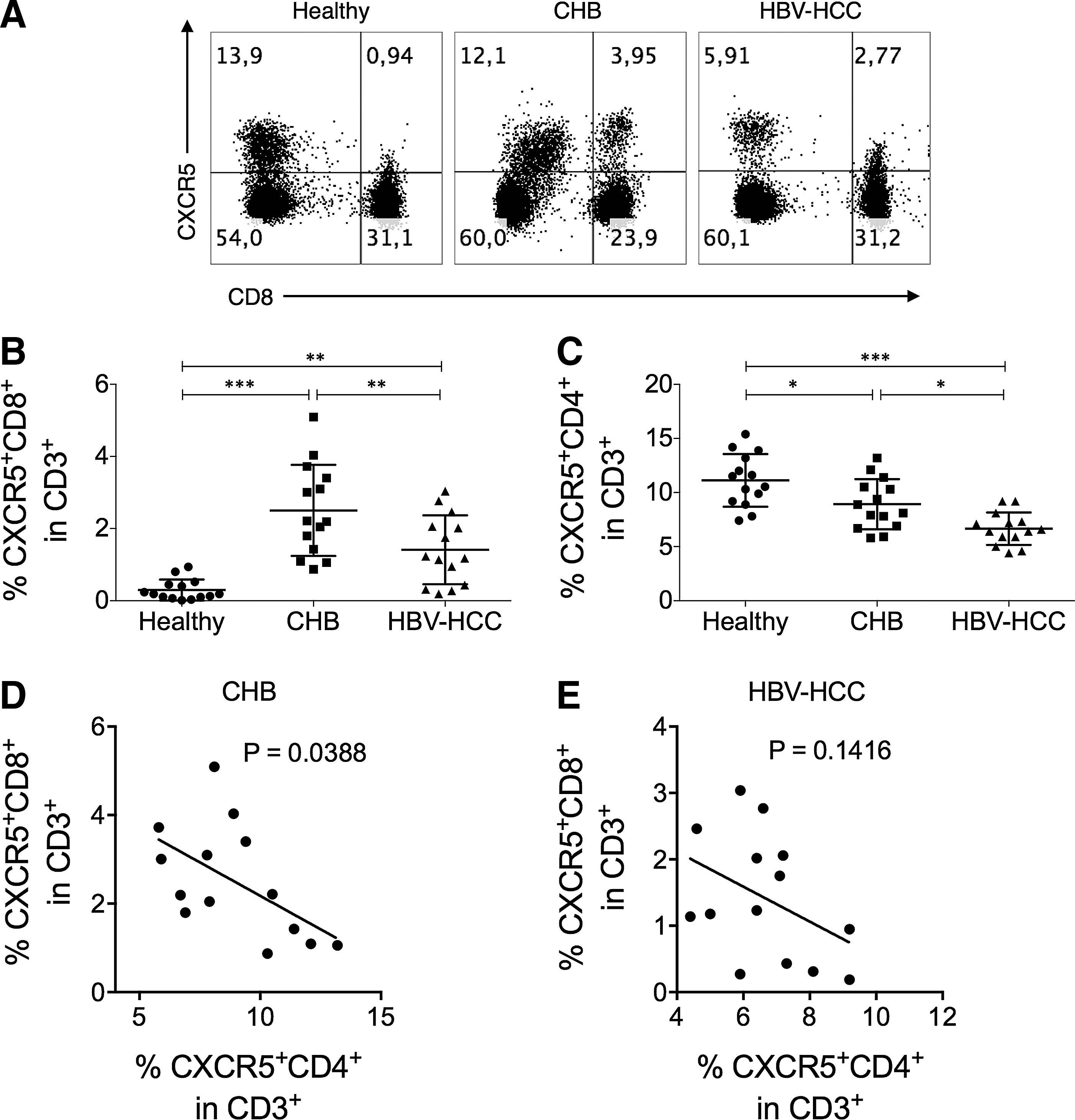
CXCR5+ CD8+ T cells were upregulated in CHB and HBV-HCC patients.
CXCR5+ CD8+ T cell displayed different cytokine expression profile from CXCR5− CD8+ T cells in CHB patients
Since we observed a strong upregulation of CXCR5+ CD8+ T cells in CHB patients compared to healthy controls, we next examined the functional characteristics of CXCR5+ CD8+ T cells in CHB patients and healthy controls. Under directly ex vivo conditions, CXCR5+ CD8+ T cells from CHB patients presented significantly higher levels of IL-21 transcription than autologous CXCR5− CD8+ T cells (Fig. 2A). After stimulation with PMA+ionomycin, the CXCR5+ CD8+ T cells from CHB patients expressed significantly higher transcription levels of IFN-γ, IL-10, and IL-21 than CXCR5− CD8+ T cells (Fig. 2B). Interestingly, CXCR5+ CD8+ T cells from CHB patients expressed significantly higher levels of IFN-γ, IL-10, and IL-21 mRNA than CXCR5+ CD8+ T cells from healthy subjects, either directly ex vivo or after PMA+ionomycin stimulation.
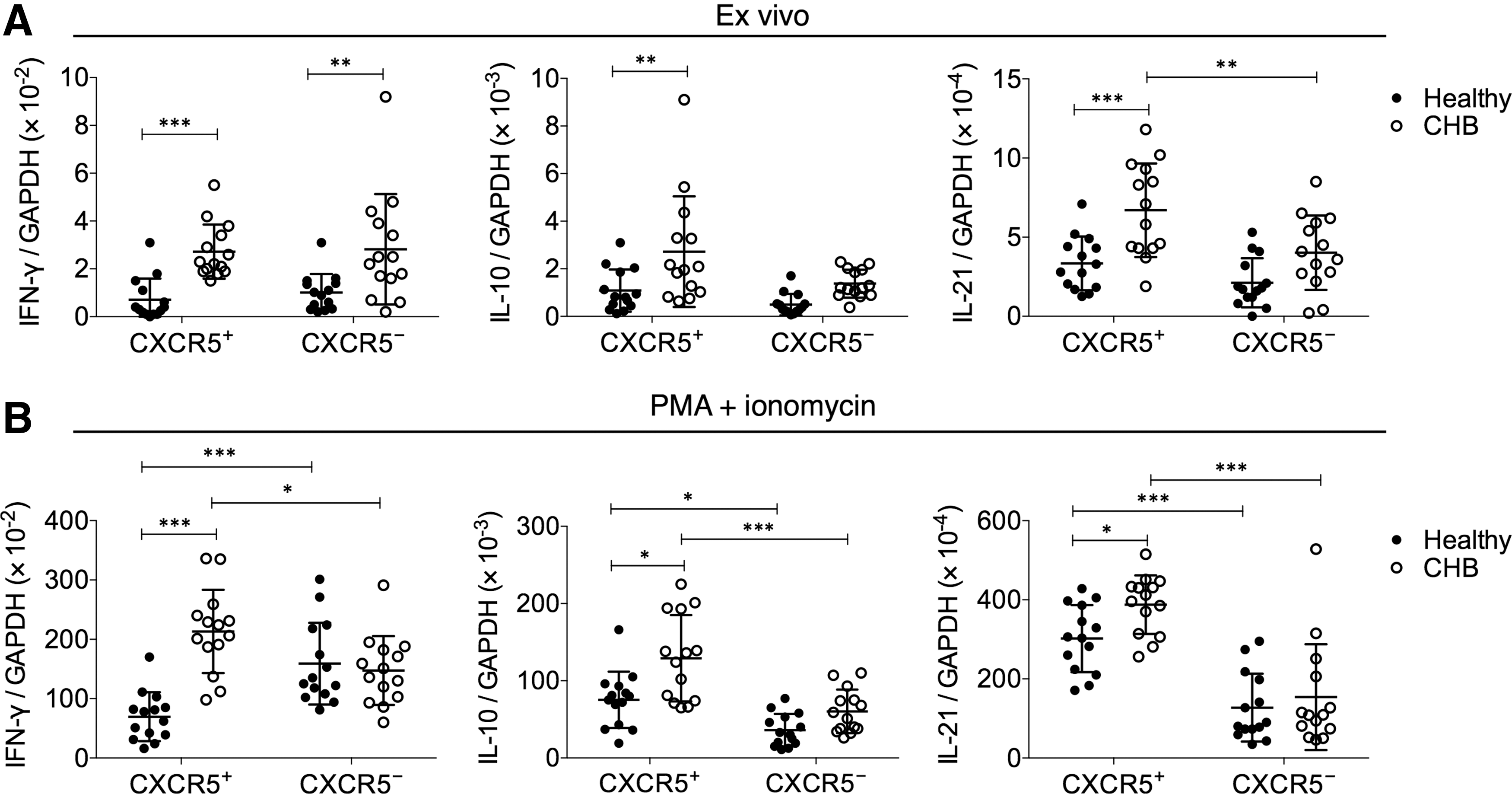
CXCR5+ CD8+ T cells in CHB patients expressed more IFN-γ, IL-10, and IL-21 transcripts than their counterparts in healthy subjects.
To corroborate the finding of transcription levels, we also examined the cytokine protein secretion in CXCR5+ versus CXCR5− CD8+ T cells. Under direct ex vivo conditions, no significant differences in IFN-γ, IL-10, or IL-21 were observed between CXCR5+ and CXCR5− CD8+ T cells, as the levels of secreted cytokines were below the detection limit in most CHB subjects (Fig. 3A). In healthy individuals, only 3 out of 14 subjects had detectable levels of IFN-γ secretion by CD8+ T cells directly ex vivo. No detectable IL-10 or IL-21 by CD8+ T cells directly ex vivo was found. Therefore, the data of CXCR5+ and CXCR5− CD8+ T cells from healthy individuals were not presented. After PMA+ionomycin stimulation, the CXCR5+ CD8+ T cells from CHB subjects expressed significantly higher levels of IL-10 and IL-21 than CXCR5− CD8+ T cells (Fig. 3B), while the IFN-γ secretion between the two groups was borderline insignificant (p = 0.056). Again, the CXCR5+ CD8+ T cells from CHB patients were more potent at secreting IFN-γ, IL-10, or IL-21 than their counterparts in healthy individuals. Together, these results demonstrated that CXCR5+ and CXCR5− CD8+ T cells presented significantly different cytokine secretion profile in CHB patients.
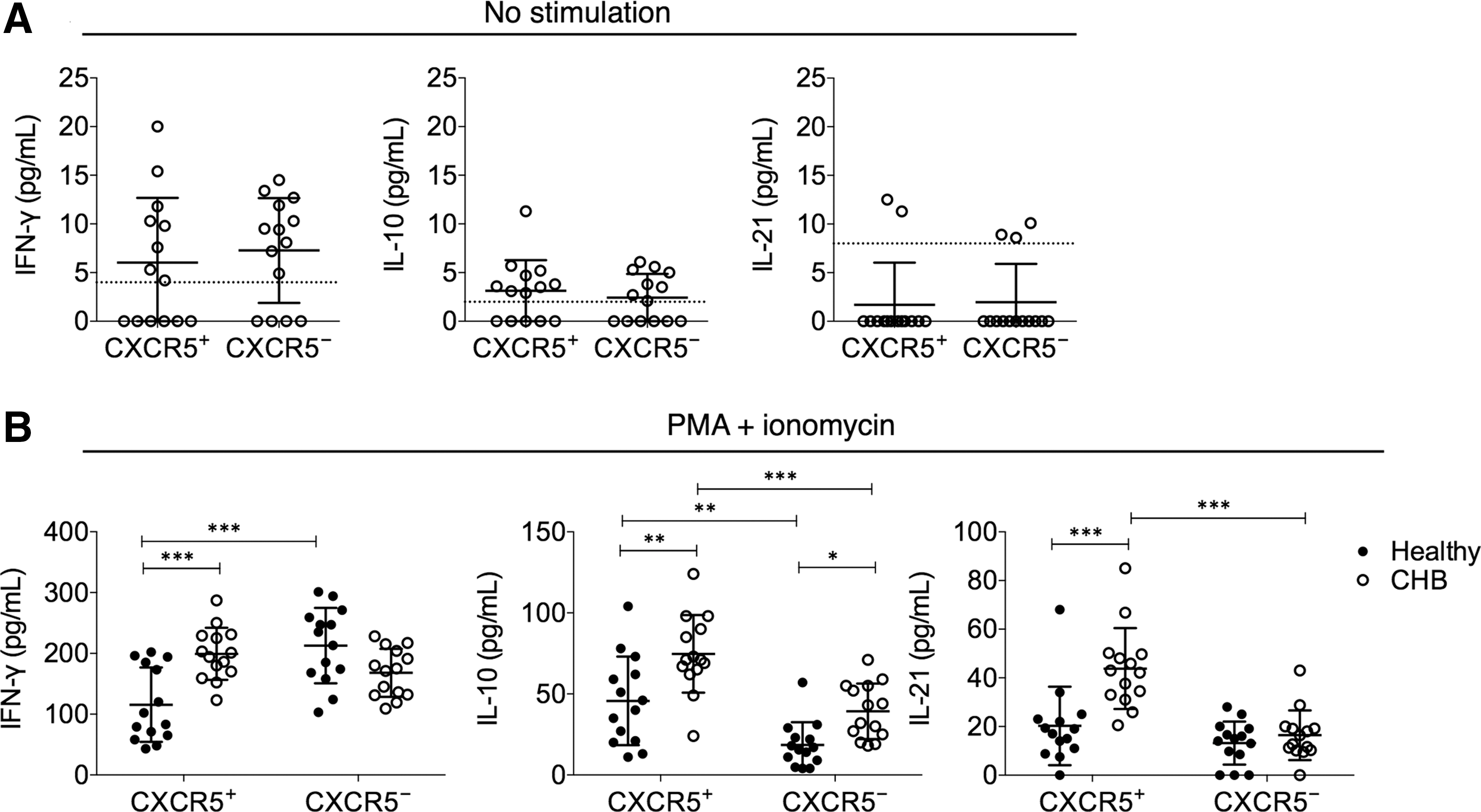
CXCR5+ CD8+ T cells in CHB patients were more potent at IFN-γ, IL-10, and IL-21 secretion than their counterparts in healthy subjects.
CXCR5+ CD8+ T cells offered B cell help in collaboration with CD4+ T cells
Previous results demonstrated that the frequency of CXCR5+ CD8+ T cells was negatively correlated with the frequency of CXCR5+ CD4+ T cells and secreted high levels of IL-10 and IL-21, which were different from CXCR5− CD8+ T cells but similar to CXCR5+ CD4+ T cells (Tangye et al., 2002; Morita et al., 2011). We wondered whether CXCR5+ CD8+ T cells could offer B cell help similar to CXCR5+ CD4+ T cells. Therefore, CD19+CD27− naive B cells were cocultured with autologous CXCR5+ CD8+ T cells or CXCR5+ CD4+ T cells. SEB was added as a superantigen. We found that unlike CXCR5+ CD4+ T cells alone, CXCR5+ CD8+ T cells alone did not significantly increase the levels of IgM, IgG, or IgA (Fig. 4A), demonstrating that CXCR5+ CD8+ T cells were not functionally equivalent to CXCR5+ CD4+ cells. Interestingly, cultures with both CXCR5+ CD8+ T cells and CXCR5+ CD4+ cells presented significantly higher levels of IgG and IgA, than cultures with CXCR5+ CD4+ cells alone, demonstrating that CXCR5+ CD8+ T cells assisted in the naive B cell help function with CXCR5+ CD4+ cells. We next examined the effect of CXCR5+ CD8+ and CXCR5+ CD4+ T cells on antibody secretion from CD19+CD27+ memory B cells. We found that memory B cells from CHB patients did not require the presence of CXCR5+ CD8+ or CXCR5+ CD4+ T cells to secrete large amounts of antibodies, but with CXCR5+ CD8+ and CXCR5+ CD4+ T cells or CXCR5+ CD4+ T cells alone, the levels of IgM and IgA were significantly elevated (Fig. 4B).
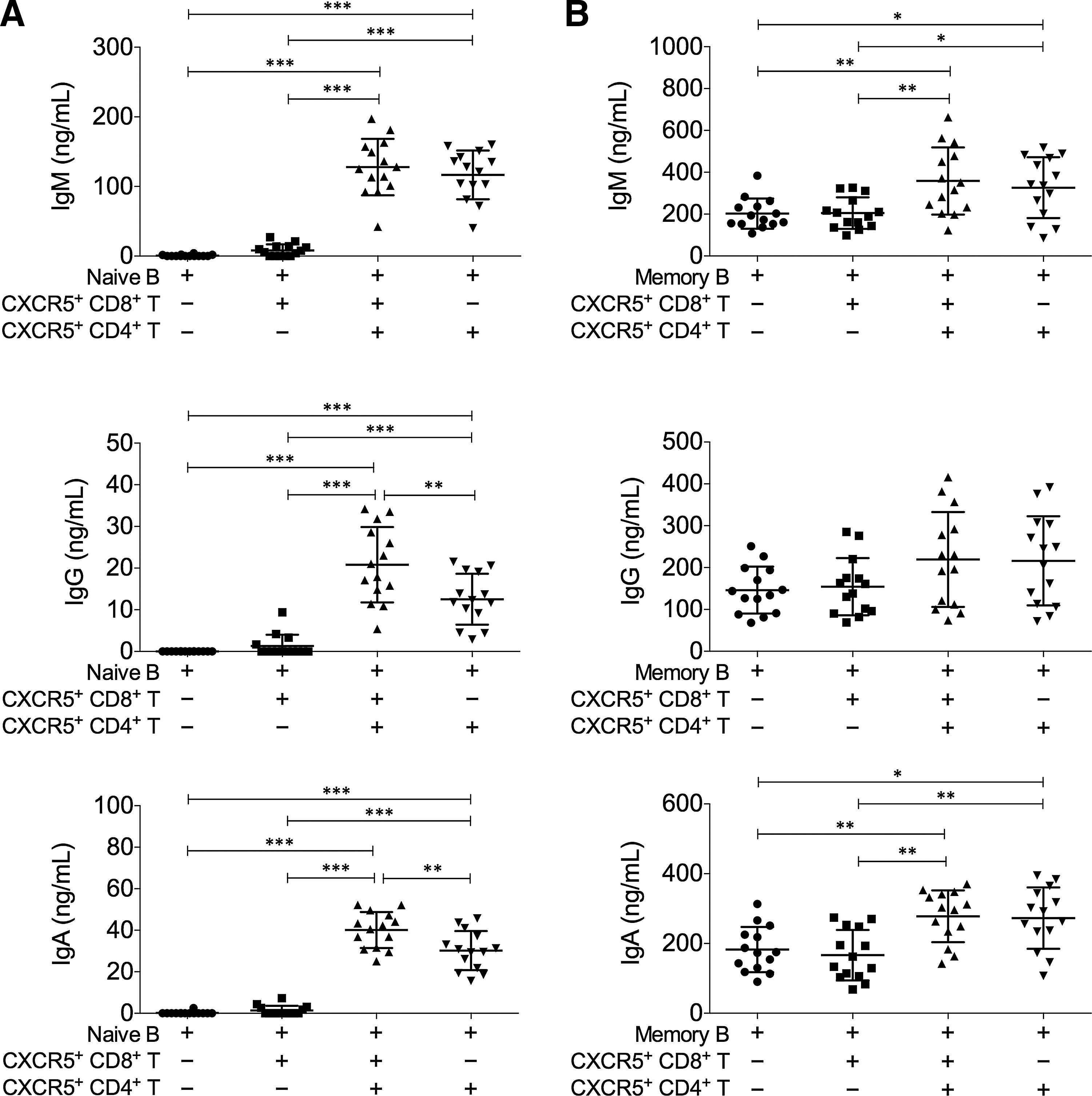
CXCR5+ CD8+ T cells assisted the naive B cell help function of CXCR5+ CD4+ T cells.
CXCR5+ CD8+ T cell frequency was inversely correlated with the alanine aminotransferase levels in 14 CHB patients
A positive HBV DNA load is indicative of active disease. The risk of cirrhosis and HCC is significantly correlated with the HBV viral load (Chen et al., 2006; Mendy et al., 2010). Therefore, the potential association between the frequency of CXCR5+ CD8+ T cells and the HBV viral load was examined in the 14 CHB patients in this study. We did not find a correlation between HBV DNA load and the frequency of CXCR5+ CD8+ T cells (Fig. 5A). The level of alanine aminotransferase (ALT) is frequently used to indicate the level of liver injury. In our CHB cohort, the level of ALT was inversely correlated with the frequency of CXCR5+ CD8+ T cells (Fig. 5B).
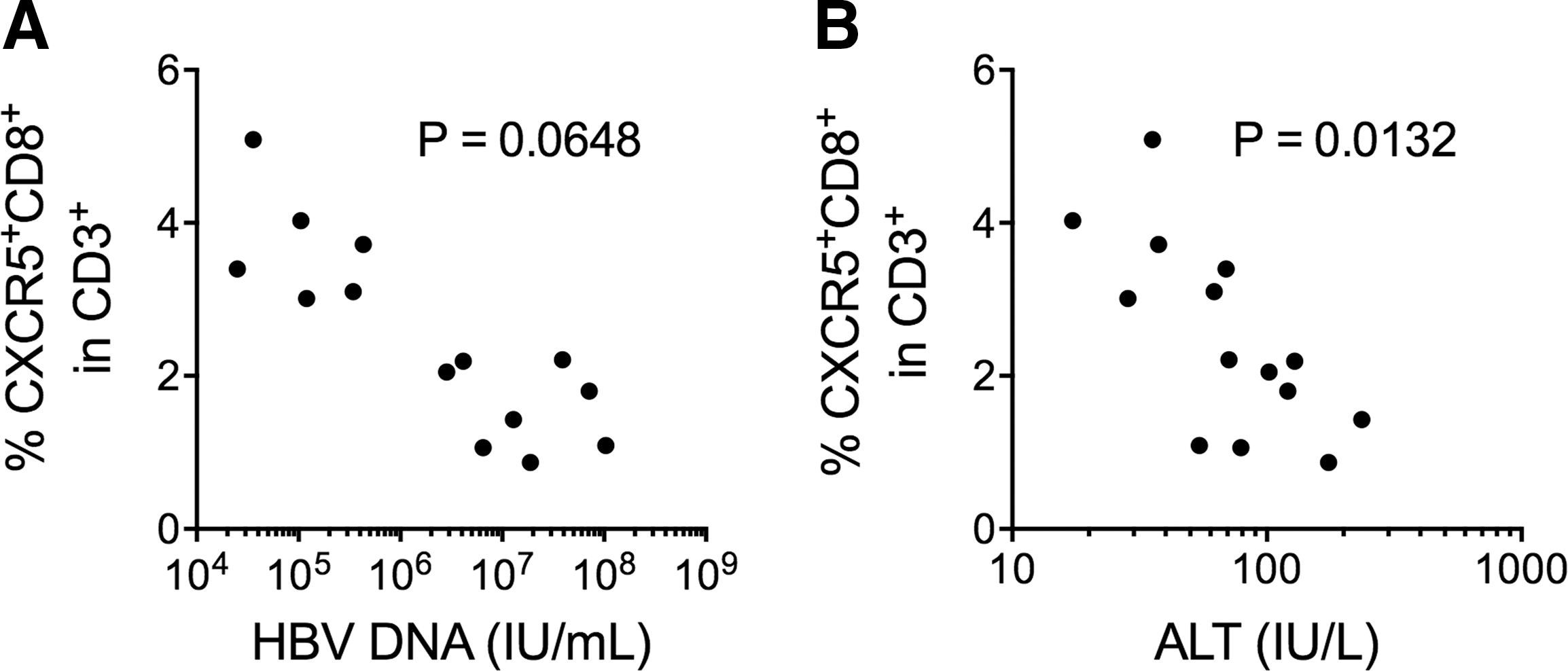
The frequency of CXCR5+ CD8+ T cells was negatively correlated with CHB ALT level.
Discussion
In this study, we observed several interesting features of CXCR5+ CD8+ T cells in CHB patients. First, we found that in healthy controls, very low frequencies of CXCR5+ CD8+ T cells could be found, while in CHB and HBV-HCC patients, the levels of CXCR5+ CD8+ T cells were significantly upregulated. These CXCR5+ CD8+ T cells in CHB patients secreted significantly higher levels of IL-10 and IL-21 than CXCR5− CD8+ T cells after stimulation, suggesting that they had different effector functions from CXCR5− CD8+ T cells.
Although IL-10 was generally regarded as an anti-inflammatory/tissue-protection cytokine, several recent studies suggested that IL-10 contributed to antitumor immunity by assisting the expansion of tumor-resident CD8+ T cells (Mumm et al., 2011; Emmerich et al., 2012). In mice with coronavirus-induced acute encephalitis, IL-10 expressing CD8+ T cells demonstrated more potent activation and cytolytic activity than IL-10 nonexpressing CD8+ T cells (Trandem et al., 2011). These studies seem to suggest that IL-10 might improve CD8+ T cell survival and activation during acute inflammation. In this study, we observed that CXCR5+ CD8+ T cells in CHB patients represented one of the CD8+ T cell subset that expressed high IL-10, while at the same time, the level of IFN-γ, a canonical CD8+ T cell activation marker, was not reduced as would be expected under high IL-10 conditions (Inoue et al., 2006; Jarnicki et al., 2006). A recent study demonstrated that CXCR5+ CD8+ T cells in chronic LCMV-infected mice presented higher cytotoxic potential and expressed lower levels of exhaustion markers (He et al., 2016), although they did not examine whether the CXCR5+ CD8+ T cells in LCMV-infected mice expressed high levels of IL-10. Combining these results, CXCR5+ CD8+ T cells might have contributed to HBV control through IL-10-mediated stimulation but not suppression. Indeed, we observed a trend that higher frequency of CXCR5+ CD8+ T cells was associated with lower ALT levels in our cohort. However, this result should be interpreted with caution, since first, CHB is a complicated disease requiring the coordination between many compartments of the immune system, and second, only 14 CHB patients were included in the study. Whether CXCR5+ CD8+ T cells could mediate the elimination of HBV-infected hepatocytes, or whether CXCR5+ CD8+ T cell-secreted IL-10 offered tissue protection (Ouyang et al., 2011), still requires further studies.
Interestingly, in CHB patients, the frequencies of CXCR5+ CD8+ T cells were negatively correlated with the frequencies of CXCR5+ CD4+ T cells, which made us wonder whether CXCR5+ CD8+ T cells were upregulated to compensate for the gradual loss of CXCR5+ CD4+ T cells from healthy individuals, CHB patients, to HBV-HCC patients. Interestingly, we found that CXCR5+ CD8+ T cells could not stimulate antibody production from naive B cells by themselves, but adding CXCR5+ CD8+ T cells to the coculture of CXCR5+ CD4+ T cells and naive B cells significantly increased the amount of IgG and IgA produced. IL-10 and IL-21 are essential B cell development and class switching cytokines (Itoh and Hirohata, 1995; Linterman et al., 2010). It is possible that CXCR5+ CD8+ T cells promoted IgG and IgA production by increasing the concentration of IL-10 and IL-21. The specific mechanisms of action by CXCR5+ CD8+ T cells, and whether it contributed to the humoral response in CHB patients, require further studies.
There are a few unresolved issues regarding the role of CXCR5+ CD8+ T cells in CHB. Previously, it was shown that follicular-homing CD8+ T cells were increased following TLR agonist and anti-CD40 stimulation and upregulated CXCR5, PD-1, and ICOS expression in mice. Depletion of CD8+ T cells in TLR/CD40-immunized mice significantly impaired the production of IgG2c, an isotype thought to be under IL-21 control, which is consistent with our results. However, in another study, the authors suggested that CXCR5+ CD8+ T cells suppressed the B cell help function of canonical Tfh (CXCR5hiPDhiBcl-6+CD4+) cells, which is in disagreement with our results. It is possible that the function of CXCR5+ CD8+ T cells is context dependent. For example, the normal function of CXCR5+ CD4+ T cells could be suppressed in CHB patients, and the enrichment of CXCR5+ CD8+ T cells could offer compensatory B cell help in CHB, but not in healthy individuals or patients with other diseases. In addition, other studies and this one suggested that CXCR5+ CD8+ T cells could serve as a novel antiviral/antitumor candidate (He et al., 2016). The antigen specificity and cytotoxicity of CXCR5+ CD8+ T cells in CHB patients, therefore, should be examined in future studies. In our study, the CXCR5+ CD8+ T cells were stimulated by PMA+ionomycin, which resulted in robust and easily detectable cytokine responses, but did not provide useful information on the antigen specificity of CXCR5+ CD8+ T cells. Using HBV peptide and protein antigens, however, did not significantly activate CD8+ T cells (Maini et al., 2000), which made the detection of HBV-specific responses in a minor CXCR5+ subset of CD8+ T cells more technically difficult. The use of tetramers loaded with HBV peptides might resolve this issue.
Footnotes
Disclosure Statement
No competing financial interests exist.