
Abstract
Crape myrtle (Lagerstroemia indica) is a woody ornamental plant popularly grown because of its long-lasting, midsummer blooms and beautiful colors. The GL1 dominant mutant is the first chlorophyll-less mutant identified in crape myrtle. It was obtained from a natural yellow leaf bud mutation. We previously revealed that leaf color of the GL1 mutant is affected by light intensity. However, the mechanism of the GL1 mutant on light response remained unclear. The acclimation response of mutant and wild-type (WT) plants was assessed in a time series after transferring from low light (LL) to high light (HL) by analyzing chlorophyll synthesis precursor content, photosynthetic performance, and gene expression. In LL conditions, coproporphyrinogen III (Coprogen III) content had the greatest amount of accumulation in the mutant compared with WT, increasing by 100%. This suggested that the yellow leaf phenotype of the GL1 dominant mutant might be caused by disruption of coproporphyrinogen III oxidase (CPO) biosynthesis. Furthermore, the candidate gene, oxygen-independent CPO (HEMN), might only affect expression of upstream genes involved in chlorophyll metabolism in the mutant. Moreover, two genes, photosystem II (PSII) 10 kDa protein (psbR) and chlorophyll a/b binding protein gene (CAB1), had decreased mRNA levels in the GL1 mutant within the first 96 h following LL/HL transfer compared with the WT. Hierarchical clustering revealed that these two genes shared a similar expression trend as the oxygen-dependent CPO (HEMF). These findings provide evidence that GL1 is highly coordinated with PSII stability and chloroplast biogenesis.
Introduction
T
Chl-less mutant plants are important for exploration of the biochemistry and molecular processes of Chl metabolism and regulation. Chl-less mutants have been identified in almost all higher plants, including rice (Oryza sativa) (Zhou et al., 2013) and Arabidopsis thaliana (Wang et al., 2016). In Triticum monococcum, chlorophyll-deficient mutant (clm1) showed pale yellow leaves, early flowering, reduced number of tillers, trichome length and density, and different shape compared with the wild type (WT) (Ansari et al. 2013). In Cucumis sativus, chlorophyll-deficient mutant C528 with golden leaf color throughout its development showed relatively weaker and slower growth with a much shorter vine length and smaller foliage in the field (Gao et al., 2016).
GL1 was the first Chl-less mutant described in crape myrtle; it was obtained from a natural yellow leaf bud mutation (Wang et al., 2013). The leaves of GL1 were orange in May, yellow during June and July, light green in early August, light yellow during the middle of August through September, and yellow in October (Li et al., 2015). The height, cutting root rate, petal length, petal width, and claw length of GL1 are less than observed in WT plants (Wang et al., 2013, 2016). However, the branch number of GL1 was significantly more than observed in WT plants (Wang et al., 2013, 2016). Furthermore, the ultrastructure of chloroplasts in the mutant leaves contains ruptured thylakoid membranes, and indistinct or absent stromal lamellae when compared with the WT (Li et al., 2015). Analyses of the mutant leaf transcriptome identified 79 unigenes involved in chlorophyll biosynthesis and degradation, photosynthesis, and chloroplast development (Li et al., 2015).
In higher plants, Chl biosynthesis is a highly coordinated process. It is executed via a series of cooperative reactions catalyzed by numerous enzymes (Beale, 2005). 5-aminolevulinic acid (ALA) synthesis is the first committed step of Chl biosynthesis (Tanaka and Tanaka, 2006). Among three enzymes responsible for ALA synthesis, glutamyl-tRNA reductase (GluTR) is regarded the main target of regulatory mechanisms; its transcription is controlled by light (Ilag et al., 1994; Cornah et al., 2003). Porphobilinogen (PBG) synthase encoded by the 5-aminolevulinic acid dehydratase (HEMB) gene catalyzes condensation of two ALA molecules to form the PBG (Beale, 1999, 2005). Uroporphyrinogen III (Urogen III) synthase catalyzes cyclization of 1-hydroxymethylbilane, with a concomitant inversion of the fourth ring of the porphyrin macrocycle, giving rise to Urogen III (Battersby et al., 1979; Beale, 1999). Three enzymes, uroporphyrinogen decarboxylase III (UROD), coproporphyrinogen III oxidase (CPO), and protoporphyrinogen IX oxidase, sequentially convert Urogen III to protoporphyrin IX (Pro IX) (Beale, 1999).
CPO catalyzes oxidative decarboxylation of the two propionate side chains of rings A and B of Coprogen III to vinyl groups (Tanaka et al., 2011). There are two structurally unrelated CPOs: the oxygen-independent CPO, HEMN, and the oxygen-dependent CPO, HEMF (Troup et al., 1994, 1995). HEMN belongs to the radical S-adenosylmethionine (SAM) family, carries an oxygen-sensitive [4Fe–4S] cluster, and requires SAM for catalysis (Layer et al., 2002). HEMF belongs to a family of monooxygenases with binuclear irons and requires oxygen for oxidative decarboxylation (Ouchane et al., 2004).
Phenotypic expression of Chl-less mutants is markedly affected by manipulation of environmental factors such as light (Biswal et al., 2012). The Chl content and ratios of chlorophyll/carotenoids of 14 species of Chl-less ornamental plants were shown to be affected by light intensity (Hu et al., 2007). In Arabidopsis, the CPO deficiency mutant lesion initiation 2 (lin2) has yellow-green leaves when grown under long day conditions and yellow leaves under short day conditions (Ishikawa et al., 2001). The barley (Hordeum vulgare), Nanchong Yellow Barley (NYB), mutant becomes more yellow and has dwarfing of seedlings in high-light (HL) conditions (Yuan et al., 2010b). Meanwhile, thylakoid structures in chloroplasts are damaged by excessive light in the Chl-less mutant. Plastid development is arrested in HL and can be derepressed by transferring to low-light (LL) conditions in Ligustrum vicaryi golden leaves (Yuan et al., 2010a). GOLDEN2-LIKE (GLK) transcription factors are required for chloroplast development (Waters et al., 2009). The expression of GLK1 is markedly lower in pale-green true leaves in the atl31/atl6 double Arabidopsis mutant than in WT plants grown under medium-light conditions (Maekawa et al., 2015).
Previously, we revealed that leaf color of the GL1 mutant is affected by light intensity (Zhang et al., 2014). However, the mechanism of the GL1 mutant on light response remained unclear. In the present study, time series are used to study the dynamic changes of chlorophyll content and its synthesis precursor content in the response of the GL1 mutant toward HL. Photosynthesis parameters, chlorophyll fluorescence, and expression of genes were measured following transfer from LL to HL (LL/HL) conditions in GL1 mutant and WT plants. Our findings indicated that the yellow leaf phenotype of the GL1 dominant mutant might be caused by disruption of Coprogen III oxidase biosynthesis.
Materials and Methods
Plant materials and growth conditions
GL1 dominant mutant was obtained from a natural yellow leaf bud mutant of the L. indica cultivar “Fenjing” at the Institute of Botany, Jiangsu Province, and the Chinese Academy of Sciences, Nanjing, China (CNBG), in 2003 (Li et al. 2015). The mutant and WT “Fenjing” cutting seedlings were grown at the CNBG under natural conditions and subsequently irradiated for the times and at the light intensities specified later in the text in 2015.
All plants were initially grown for 8 weeks under LL conditions (40 μmol photons m−2 s−1). Subsequently, they were exposed to HL (1000–1500 μmol photons m−2 s−1). The third to fifth leaves were harvested from both mutant and WT plants in all experiments. Harvested samples were either immediately analyzed or kept at −80°C until required. Leaf numbers were counted from the top of the shoot.
Chlorophyll and carotenoid determination
Fresh leaves of WT and GL1 mutant plants were collected at four time points (0, 48, 96, and 168 h after LL/HL transfer). Pigments were extracted from mature leaves according to the methods described by Li (2000). Chl and carotenoids were extracted with 95% ethanol for 3 days, and spectrophotometrically quantified (DU-800; Beckman Coulter, Brea, CA) at 470, 649, and 665 nm.
Assay of antioxidant enzyme activities
Fresh leaves of WT and GL1 mutant plants were collected at four time points (0, 48, 96, and 168 h after LL/HL transfer). Superoxide dismutase (SOD) activity was measured using the xanthine oxidase method (Sun et al., 1988). Absorbance was determined at 550 nm. Catalase (CAT) activity was measured using the ammonium molybdate spectrophotometric method (Góth, 1991), which is based on the fact that ammonium molybdate can rapidly terminate the H2O2 degradation reaction catalyzed by CAT and reacts with residual H2O2 to generate a yellow complex; this was monitored by absorbance at 405 nm. Peroxidase (POD) activity was monitored by absorbance at 420 nm. All experiments were carried out using the following kits: SOD assay kit/A001-3, CAT assay kit/A007-2, and peroxidase assay kit/A084-3 (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) according to the manufacturer's instructions.
Content of chlorophyll synthesis precursors
Fresh leaves of WT and GL1 mutant plants were collected at 09:00 a.m. (0 h), 11:00 a.m. (2 h), 1:00 p.m. (4 h), and 4:00 p.m. (7 h). 5-aminolevulinic acid was extracted as described by Dei (1985). Isolations of PBG and uroporphyrinogen were performed using the method of Bogorad (1962). Coprogen III, Proto IX, Mg-proto, and Pchlide were determined according to the methods of Rebeiz et al. (1975). Three replicates were performed for each sample. Contents of Chl precursors were determined by spectrophotometrical methods (DU-800; Beckman Coulter, Brea, CA).
Transmission electron microscopy
Fresh leaves of WT and GL1 mutant plants were collected at four time points following LL/HL transfer (0, 48, 96, and 168 h). Leaf samples were fixed overnight at 4°C in 3% glutaraldehyde and 0.1 M sodium cacodylate buffer (pH 7.4) and then processed for electron microscopy as described by Sakamoto et al. (2009). Ultrathin sections were cut using an ultramicrotome (Ultracut, Reichert-Jung) and observed with a transmission electron microscope (TEM 300; Hitachi Comp., Japan) operating at 75 kV.
Leaf gas exchange measurement
The Li-Cor 6400 portable photosynthesis system (LI-COR Biosciences, Lincoln, NE) with a built-in light source set at 1000 μmol photons m−2 s−1 was used to determine net photosynthesis (Pn), stomatal conductance (Cond), intercellular CO2 concentration (Ci), and transpiration rate (Trmmol). All measurements were carried out between 09:00 a.m. and 11:00 a.m., at four time points following LL/HL transfer (0, 48, 96, and 168 h). During measurements, leaf temperature was adjusted to 25°C.
Chlorophyll fluorescence
Chlorophyll fluorescence parameters were measured using an OS1p fluorometer (Opti-Sciences, Hudson, NH). Leaves were dark adapted for 30 min before measurement to ensure all capable PSII reaction centers were fully oxidized. Fluorescence parameters were calculated based on methods described by Genty et al. (1989); Hendrickson et al. (2004); and Kramer et al. (2004) as follows: maximum quantum yield of PSII, Fv /Fm = (Fm –Fo )/Fm ; potential photochemical efficiency, Fv /Fo = (Fm –Fo )/Fo ; effective quantum yield of PSII, Y(II) = (Fm –Fs )/Fm ’; Y(NPQ) and Y(NO) represent the quantum yield of regulated and nonregulated energy dissipation in PSII, respectively; Y(NPQ) = Fs/Fm –Fs/Fm ; Y(NO) =Fs/Fm ; non-photochemical quenching; NPQ = (Fm –Fm )/Fm .
Fo and Fm are the minimum and maximum fluorescence measured after 30 min of dark adaptation. Fm’ is the maximum fluorescence after light adaptation and is measured on illumination of a saturating pulse; Fs is the steady-state fluorescence after light adaptation. All measurements were carried out at 09:00 a.m. (0 h), 11:00 a.m. (2 h), 1:00 p.m. (4 h), and 4:00 p.m. (7 h).
RNA extraction and quantitative real time-PCR analysis
Fresh leaves of WT and GL1 mutant plants were collected at four time points after LL/HL transfer (0, 48, 96, and 168 h). Total RNAs were extracted using the TRIzol reagent according to the manufacturer's instructions (Invitrogen, Carlsbad, CA). Total RNA concentration was quantified using One Drop OD–2000 Spectrophotometer (Nanjing Wuyi Technology Co., Nanjing, China) and quality checked by electrophoresis in a 1% agarose gel.
For each sample, 2 μg of total RNA were reverse transcribed into cDNA using M-MLV reverse transcriptase (Promega, Madison, WI). Reverse transcription was performed at 37°C for 60 min in a 20 μL reaction with a final denaturation step at 95°C for 5 min. Primers were designed using Primer premier 5 (
The Actin gene from L. indica was amplified as an internal control for data normalization. The 25 μL reaction volumes contained 12.5 μL SYBR Green PCR Master Mix, 9.5 μL deionized H2O, 1 μL primers, and 2 μL cDNA. Amplification reactions were initiated with a predenaturation step (95°C for 10 min), followed by 40 cycles of denaturing (95°C for 10 s), annealing (60°C for 35 s), and extension (72°C for 35 s). Data were processed using the 2−ΔΔCt method (Livak and Schmittgen, 2001). Heat maps for the gene expression profiles were generated using the MultiExperiment Viewer (MeV 4) software package (Saeed et al., 2003).
Sequence analysis of HEMN genes
The HEMN protein sequences from A. thaliana (At), O. sativa (Os), Populus trichocarpa (Pt), Theobroma cacao (Tc), Gossypium raimondii (Gr), Gossypium arboreum (Ga), Vitis vinifera (Vv), Brassica napus (Bn), Microcoleus sp. PCC7113, Zea mays (Zm), Malus domestica (Md), Eucalyptus grandis (Eg), Ziziphus jujube (Zj), Fragaria vesca (Fv), Capsella rubella (Cr), Citrus clementine (Cc), Citrus sinensis (Cs), and Rivularia sp. PCC 7116 were downloaded from GenBank (
Statistical analysis
One-way analysis of variance was used to determine the significance between the mean values of the GL1 mutant and WT. Comparison among means was performed using the Duncan's multiple range test and the two-tailed unequal variance t-test in SPSS 17.0 (SPSS, Inc., Chicago).
Results
LL/HL transfer leads to reduction of Chl b content and lesion formation in the GL1 mutant
Under LL conditions (t0), the GL1 dominant mutant leaf color was golden-yellow, while the WT was light yellow-green (Fig. 1a). When the mutant and WT were exposed to HL conditions for 1 week (t168), the leaf color of the mutant rapidly changed to a pale-green, whereas it changed to green in the WT (Fig. 1d). Under LL conditions, Chl a content of the mutant was markedly lower than observed in the WT; there was no difference in Chl b content between the mutant and WT (Table 1). Exposure of the LL-grown mutant plants to HL resulted in a drop in Chl a, Chl b, and total Chl content, but raised carotenoid content and the Chl a/b ratio (Table 1). Compared with LL conditions, after 168 h of HL, the Chl a and Chl b contents of the mutant were significantly reduced, by 47.89% and 70.26%, respectively, whereas carotenoid content increased by 62.61% (Table 1). These data suggested that changes of leaf color in the mutant might be caused by a significant reduction of Chl b content.
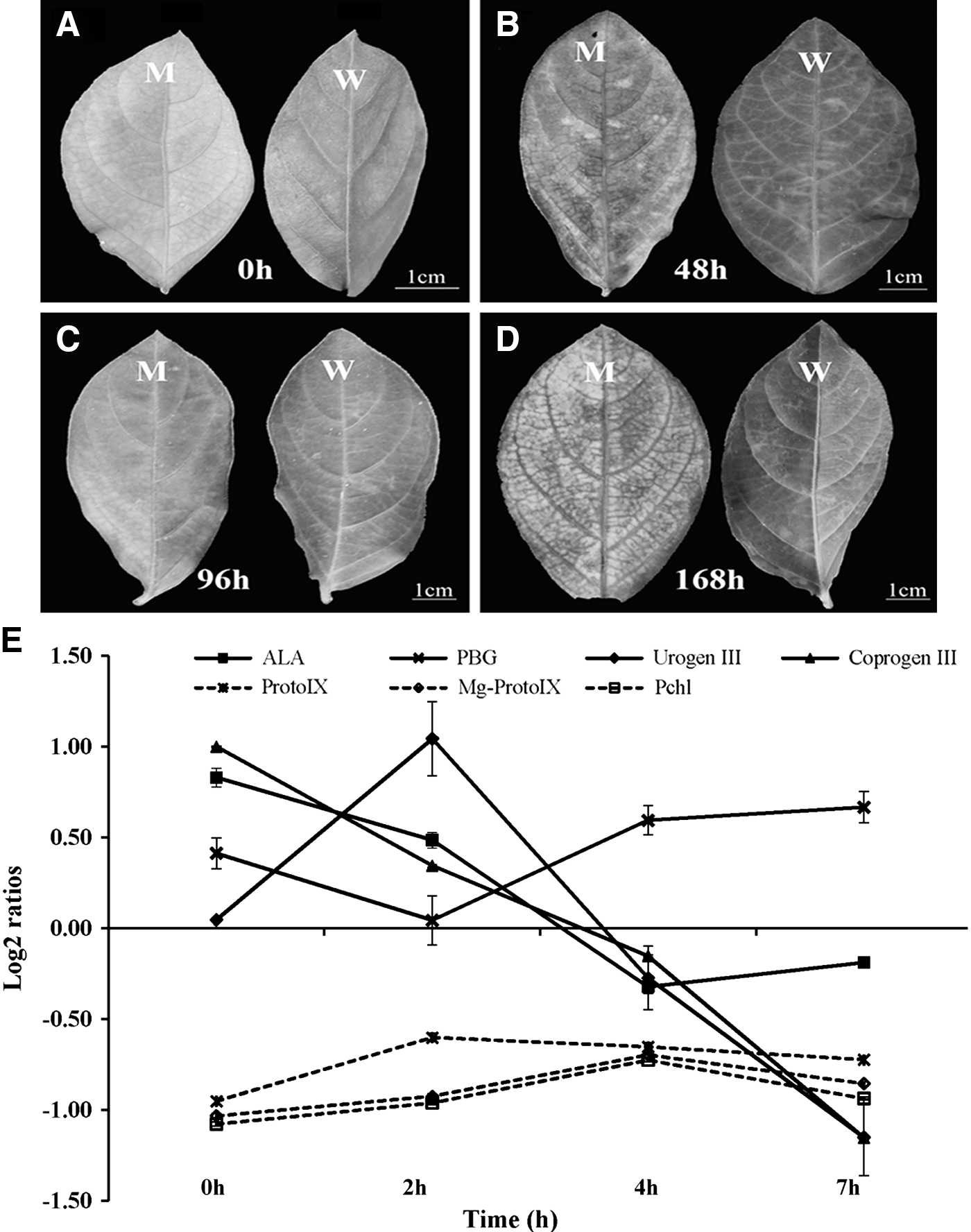
Phenotypic characterization of GL1 mutant after low-light/high-light transfer.
The data represent the mean ± SE of n = 3 replicates.
The letters mean a significant difference at p < 0.05 (Duncan's multiple test).
Asterisk means significantly different between mutant and wild type at p < 0.05 (two-tailed unequal variance t-test).
SE, standard error.
Lesions appeared on both mutant and WT leaves within the first 48 h of HL (Fig. 1b). Lesions in the mutant gradually became severe, with the leaf margin beginning to die on increased illumination time (Fig. 1c, d). Contrastingly, in the WT, lesion formation after LL/HL transfer peaked at t96, and was followed by a slow recovery (Fig. 1c, d). Thus, the yellow leaf was more sensitive to photodamage. To characterize photodamage in more detail, the antioxidant enzyme activities of SOD, CAT, and POD were investigated in GL1 mutant and WT plants after LL/HL transfer. SOD activity in the mutant gradually decreased following LL/HL transfer (Table 2). CAT activity in the mutant showed the greatest value at t48 and was always higher than in WT, except at t0 (Table 2). Initially, POD activity in the mutant was markedly decreased, but this was followed by a rapid recovery; activity was always higher than in the WT after LL/HL transfer, except at t48 (Table 2). After 1 week (168 h) in HL conditions, the activities of SOD, CAT, and POD in the mutant were 0.87-, 1.34-, and 1.94-fold of that observed in the WT, respectively (Table 2). These results further indicated that the GL1 mutant senses a higher level of photodamage than the WT.
The data represent the mean ± SE of n = 3 replicates.
The letters mean a significant difference at p < 0.05 (Duncan's multiple test).
Asterisk means significantly different between mutant and wild type at p < 0.05 (two-tailed unequal variance t-test).
SOD, superoxide dismutase; CAT, catalase; POD, peroxidase.
Precursors of the Chl biosynthesis pathway were measured in the mutant and the WT following LL/HL transfer. ALA, PBG, Urogen III, and Coprogen III contents in the GL1 mutant were significantly higher (log2 ratios >0) than in the WT, while Proto IX, Mg-protoporphyrin IX (Mg-proto), and protochlorophyllide (Pchlide) contents were lower in LL conditions (log2 ratios <0) (Fig. 1e; Supplementary Table S2). Among the precursors, Coprogen III had the greatest accumulation, increasing by 100% in the mutant compared with the WT in LL (Fig. 1e; Supplementary Table S2). However, the contents of ProtoIX, Mg-proto, and Pchlide in the mutant were always significantly lower than in the WT (log2 ratios <0) after LL/HL transfer (Fig. 1e; Supplementary Table S2). These results indicated that in L. indica, the GL1 mutant was a Coprogen III-less mutant. Interestingly, light responses of the precursors were different in the GL1 mutant, with the PBG content of the mutant being significantly higher than observed in WT (log2 ratios >0) after the LL/HL transfer (Fig. 1e). However, ALA, Urogen III, and Coprogen III content of the GL1 mutant was higher compared with WT (log2 ratios >0) within the first 2 h; this was followed by a rapid decrease (log2 ratios <0) (Fig. 1e).
The structure of grana lamellae was destroyed in the GL1 mutant after LL/HL transfer
How the GL1 mutant affected chloroplast development after LL/HL transfer was investigated by comparing the ultrastructure of chloroplasts in the mutant and WT at t0, t48, t96, and t168 using transmission electron microscopy. At t0, the structure of grana lamellae was similar in both GL1 and WT (Fig. 2). After LL/HL transfer, the structure of grana lamellae was distinctly different in the GL1 mutant and WT. The length and density of grana lamellae gradually decreased in the mutant on increased illumination times, with absence of grana lamellae structure at t168 (Fig. 2). In contrast, the length and density of grana lamellae in the WT gradually increased following the LL/HL transfer (Fig. 2). The data suggested that the structure of grana lamellae was destroyed in the GL1 mutant after LL/HL transfer.
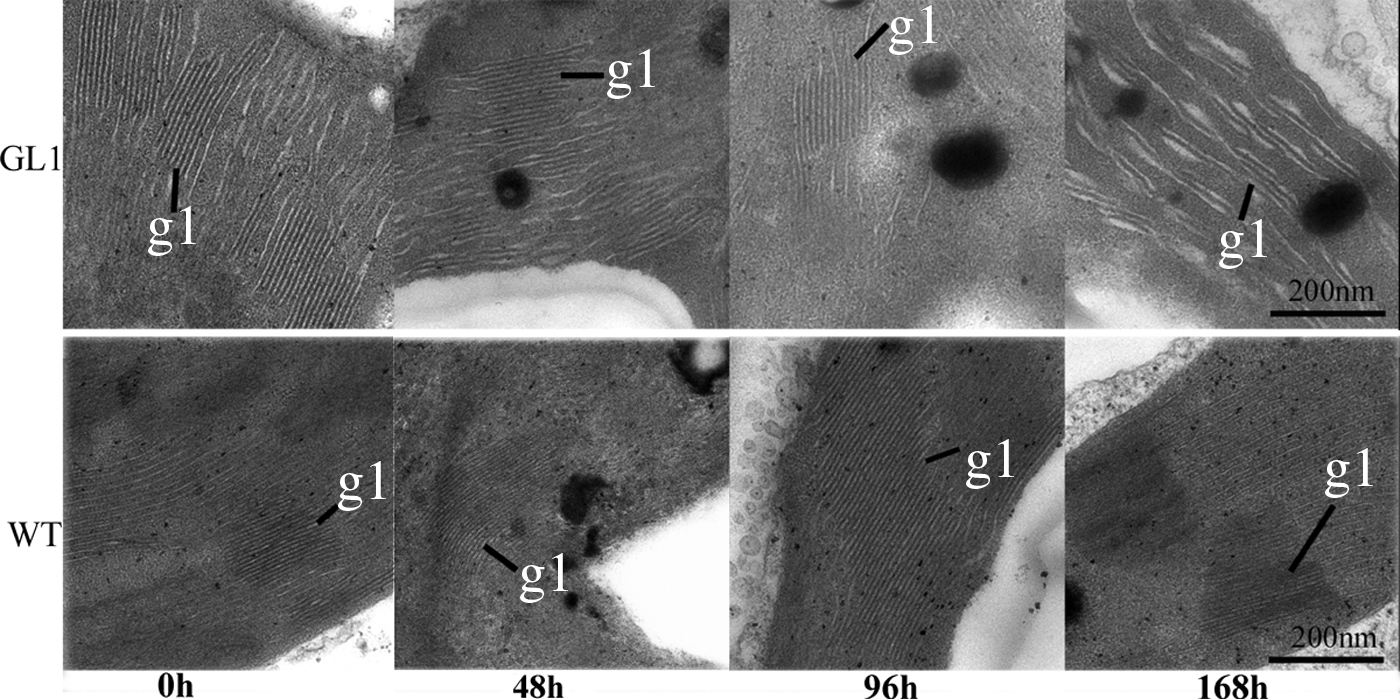
Ultrastructure of chloroplasts in the GL1 mutant and wild type after low-light/high-light transfer. gl, grana lamella, scale bar = 200 nm.
During LL/HL transfer, damages to photosystem II in the GL1 mutant lead to decreased photochemical capacity
There were no differences in the values of net photosynthesis (Pn), stomatal conductance (Cond), intercellular CO2 concentration (Ci), and transpiration rate (Trmmol) (log2 ratios ≈0) between the GL1 mutant and WT in the first 48 h after LL/HL transfer (Fig. 3a; Supplementary Table S3). However, after 96 h in HL, the values of Pn, Cond, and Trmmol (log2 ratios <0) in the mutant decreased significantly, whereas values of Ci (log2 ratios >0) increased (Fig. 3a; Supplementary Table S3). In comparison with LL, Pn decreased by 30.29%, Cond by 16.67%, and Trmmol by 29.17%, whereas Ci increased by 58.07% after 1 week (168 h) in HL in the mutant plants (Supplementary Table S3). The above data suggested that the reduction of net photosynthesis in the GL1 mutant was caused by nonstomatal factors.
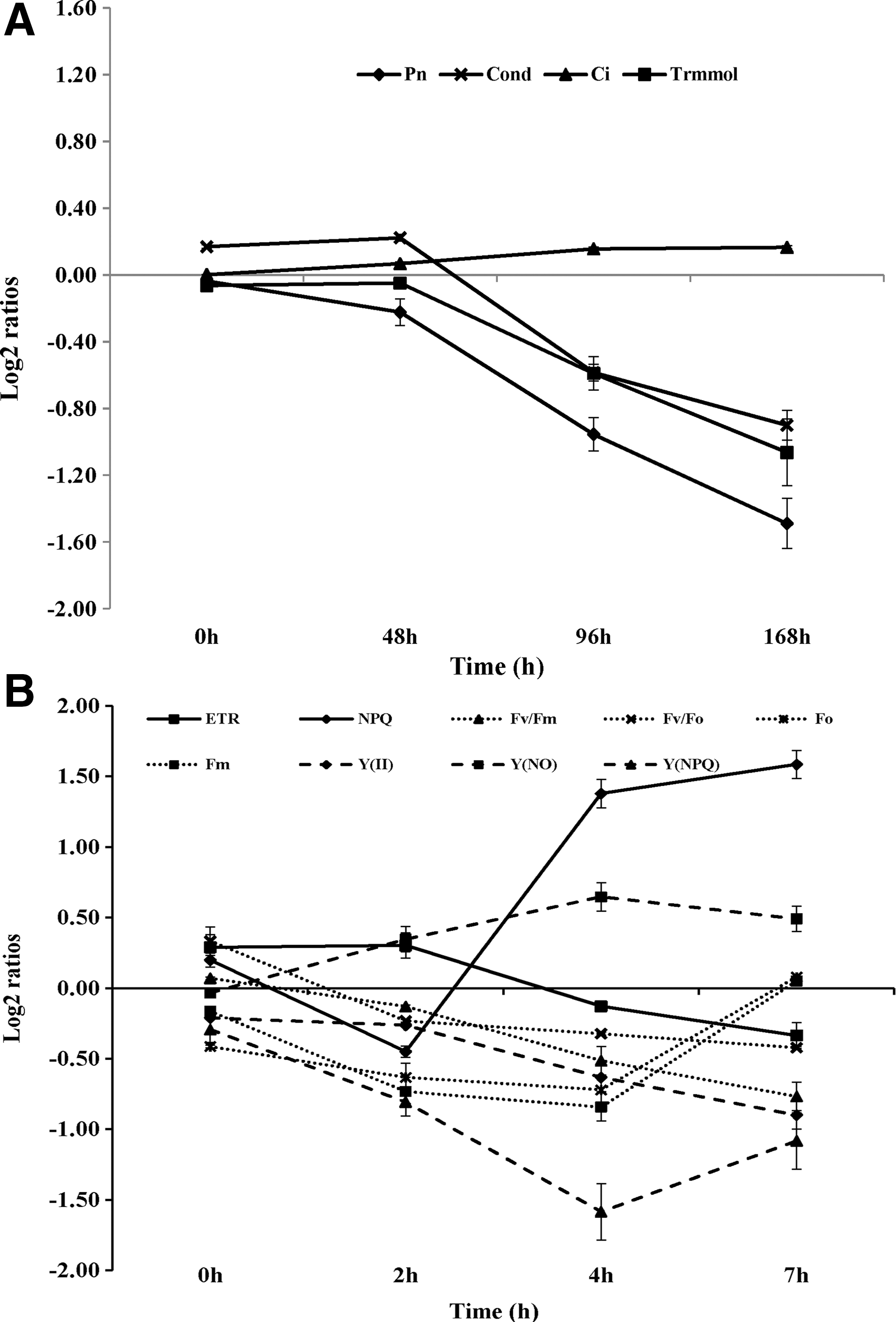
Photosynthetic characters and chlorophyll fluorescence parameters of GL1 mutant and wild type after low-light/high-light transfer.
Chlorophyll fluorescence parameters were measured to better understand the reasons for decreased net photosynthesis in the mutant following LL/HL transfer. The maximum quantum yield of PSII, as measured by Fv /Fm , did not differ between the GL1 mutant and WT (log2 ratios ≈0) with values being close to 0.8 under LL conditions (Fig. 3b; Supplementary Table S4). However, Fo and Fm values in the mutant were significantly lower than in the WT under LL (Fig. 3b; Supplementary Table S4). The ETR value was also significantly higher in the mutant than in the WT under LL conditions (Fig. 3b; Supplementary Table S4). Following the LL/HL transfer, the Fv /Fm and ETR values in the mutant were decreased, while those of the WT initially decreased rapidly and then recovered (Fig. 3b; Supplementary Table S4). Fo values were increased significantly in the GL1 mutant, while Fm values were rapidly decreased after the LL/HL transfer (Fig. 3b; Supplementary Table S4). In the mutant plants, the Fv /Fm value decreased by 53.75%, ETR by 34.58%, and Fm by 61.34% after 1 day in HL (7 h), while Fo increased by 16.95% (Supplementary Table S4). The data suggested that the decrease of Fv /Fm was mainly due to a large decrease in maximal chlorophyll fluorescence.
There was no significant difference in NPQ values (log2 ratios ≈0) between the mutant and WT at t0 following the LL/HL transfer (Fig. 3b). However, NPQ values of the mutant were significantly lower at t2 (log2 ratios <0) and markedly higher at t4 and t7 (log2 ratios >0) compared with the WT after LL/HL transfer (Fig. 3b; Supplementary Table S4). Y(II), Y(NO), and Y(NPQ) values of the GL1 mutant were always lower than in WT plants (log2 ratios <0) after the LL/HL transfer (Fig. 3b; Supplementary Table S4). The Y(NO) values of the mutant were significantly higher than in WT plants under HL conditions (Fig. 3b; Supplementary Table S4). In mutant plants grown in HL for 1 day (7 h), Y(II) decreased by 60.53% and Y(NPQ) by 45.16%, whereas Y(NO) increased by 43.90% (Supplementary Table S4). The above data indicated that damages to photosystem II (PSII) in the GL1 mutant lead to decreased photochemical capacity after LL/HL transfer.
Expression analysis of chlorophyll- and photosynthesis-associated genes in the GL1 mutant and WT
In our previous study, the leaf transcriptome of the GL1 mutant was sequenced. This identified 79 unigenes involved in chlorophyll biosynthesis and degradation, photosynthesis, and chloroplast development (Li et al., 2015). In this study, the transcript levels of 23 genes were selected for assay in the mutant and WT after LL/HL transfer. Transcript levels of 11 chlorophyll-associated genes were assayed: glutamyl-tRNA synthetase (GluRS), glutamate-1-semialdehyde aminotransferase (HEML), glutamyl-tRNA reductase (HEMA), HEMB, hydroxymethylbilane synthase (HEMC), uroporphyrinogen-III synthase (HEMD), HEMF, HEMN, protoporphyrinogen oxidase (HEMG), chlorophyllide a oxygenase (CAO), and chlorophyllase (Chlase) (Fig. 4). In comparison with the wild type, the expression levels of HEMA, HEMD, and HEMN were decreased in the mutant following the LL/HL transfer (Fig. 4). Interestingly, the expression level of HEMA in the mutant was significantly decreased at t0 and that of HEMD greatly decreased at t168 compared with the WT following the LL/HL transfer (Fig. 4). Furthermore, the expression levels of HEMF in the mutant increased after the LL/HL transfer (Fig. 4).
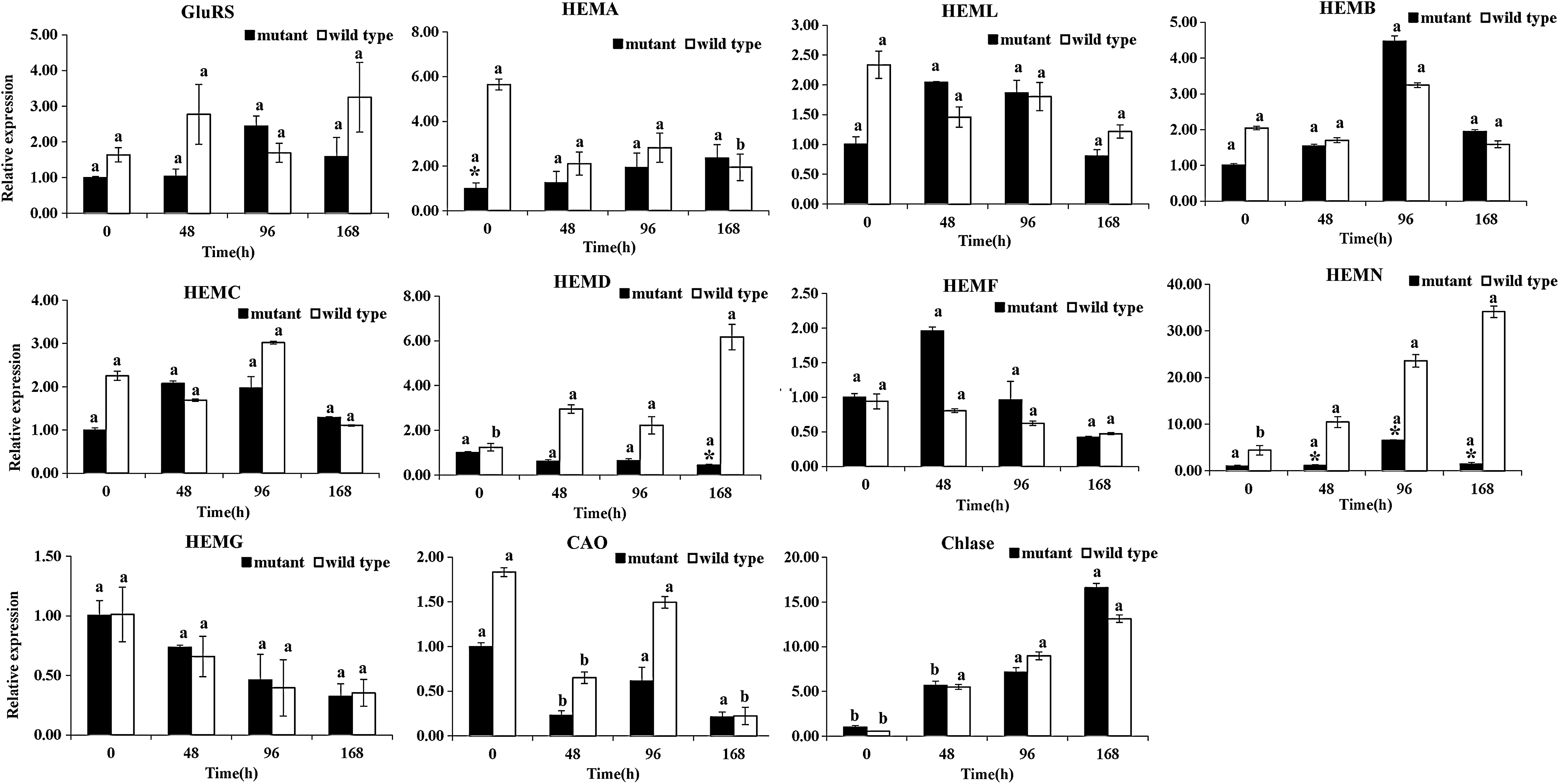
Expression of chlorophyll-associated genes in GL1 mutant and wild type after LL/HL transfer. The Actin gene from L. indica was amplified as an internal control for data normalization. Each value represents the mean of three independent replicates ± SE. The letters mean a significant difference at p < 0.001 (Duncan's multiple test). Asterisk means significantly different between mutant and wild type at p < 0.001 (two-tailed unequal variance t-test). SE, standard error.
To under the structural difference between the GL1 mutant and WT, the full-length LiHEMN cDNA sequences were cloned by RT-PCR. The LiHEMN cDNA from mutant and WT was 1431 and 1407 bp in length, respectively (Supplementary Fig. S1). Sequence comparison revealed that two inserted fragments were detected from the GL1 mutant sequence (Supplementary Fig. S1). Based on the phylogenetic tree of HEMN, we found that LiHEMN and EgHEMN showed the closest relationship (Supplementary Fig. S2). Therefore, the data suggested that the HEMN gene might be responsible for the yellow leaf phenotype of the GL1 mutant.
Twelve photosynthesis-associated genes were assessed: cytochrome b6-f complex iron–sulfur subunit (petC), ferredoxin—NADP+ reductase (petH), photosystem I P700 chlorophyll a apoprotein A1 (psaA), photosystem I subunit XI (psaL), photosystem II CP47 chlorophyll apoprotein (psbB), photosystem II CP43 chlorophyll apoprotein (psbC), photosystem II 10 kDa protein (psbR), light-harvesting complex I chlorophyll a/b binding protein 2 (LHCA2), light-harvesting complex II chlorophyll a/b binding protein 1 (LHCB1), chlorophyll a/b binding protein gene (CAB1), stomatal density and distribution 1 (SDD1), and GLk2. The transcript levels of LHCB1, psbR, CAB1, SDD1, and GLk2 were significantly changed in the mutant after the LL/HL transfer compared with the WT (Fig. 5). Expression level of LHCB1 in the mutant was greatly decreased at t48, while the psbR gene had significantly decreased mRNA levels at t96 (Fig. 5). The CAB1 gene showed remarkably decreased mRNA levels for the first 96 h after LL/HL transfer, this was followed by significantly enhanced mRNA levels (Fig. 5). In addition, the expression levels of SDD1 in the mutant were markedly reduced at t48 and t96 and those of GLk2 significantly decreased at t48 (Fig. 5).

Expression of photosynthesis-associated genes in GL1 mutant and wild type after LL/HL transfer. The Actin gene from L. indica was amplified as an internal control for data normalization. Each value represents the mean of three independent replicates ± SE. The letters mean a significant difference at p < 0.001 (Duncan's multiple test). Asterisk means significantly different between mutant and wild type at p < 0.001 (two-tailed unequal variance t-test).
Hierarchical clustering revealed that HEMN shared a similar expression trend as GluRS, HEML, HEMB, HEMC, petC, and petH (Supplementary Fig. S3). In addition, HEMF showed a similar expression trend to CAB1 and psbR (Supplementary Fig. S3). These findings indicated that the molecular mechanism of the GL1 dominant mutant is complicated.
Discussion
Chl plays significant role in absorption of sunlight by photosynthetic reaction centers (Liu et al., 2007). Chl-deficient mutants or leaf-color mutants are ideal materials for fundamental research in photosynthesis, photomorphogenesis, hormone physiology, resistance mechanism, and identification of gene functions (Ansari et al., 2013; Guo et al., 2013; Gao et al., 2016). In addition, Chl-deficient mutants could be applicable as a leaf-color marker gene for efficient identification and elimination of false hybrids in commercial hybrid rice and cucumber production (Wan et al. 2015; Gao et al., 2016). Meanwhile, Chl-deficient mutants can be bred as ornamental plant varieties such as L.indica “Jinhuang” (Wang et al., 2016). Crape myrtle is well known for its long-lasting midsummer blooms and beautiful colors. GL1 was the first golden-leaf mutant described in crape myrtle (Wang et al., 2013). We have researched on the mutant for many years. However, the molecular mechanism of GL1 has not been illuminated.
The yellow leaf phenotype of the GL1 dominant mutant might be caused by disruption of CPO biosynthesis
Chl metabolism is a highly coordinated process executed via a series of cooperative reactions catalyzed by numerous enzymes (Beale, 2005). CPO catalyzes the oxidative decarboxylation of Coprogen III into Proto IX, and is an important regulatory step in the Chl biosynthetic pathway. Coprogen III belongs to a group of photosensitive tetrapyrroles, and reducing its levels causes light-dependent yellow leaf and lesion formation in tobacco (Nicotiana tabacum), Arabidopsis, and rice (Kruse et al., 1995b; Ishikawa et al., 2001; Guo et al., 2013; Wang et al., 2015).
In CPO-deficient lin2 plants, young leaves are pale green and develop lesions. Lesions begin as a small spot or stripe of collapsed tissue, usually first appearing in the middle part of young leaves (Ishikawa et al., 2001). Similarly, lesions were generated and the leaf color changed from yellow- to pale-green in the GL1 mutant following LL/HL transfer (Fig. 1a–d). Thus, the phenotypic characteristics of the GL1 mutant were similar to those of the Arabidopsis lin2 mutant. Transgenic tobacco plants with reduced CPO activity are characterized by antioxidative responses and necrotic leaf lesions (Kruse et al., 1995a). In this study, we investigated antioxidant enzyme activities of SOD, CAT, and POD in GL1 mutant and WT plants following LL/HL transfer. After 1 week (168 h) in HL conditions, SOD, CAT, and POD activities in the mutant were 0.87-, 1.34-, and 1.94-fold of that observed in the WT, respectively (Table 1).
In the rice PORB-deficient (fgl) mutant, contents of ALA, Proto IX, and Mg-ProtoIX are greatly decreased under HL conditions (Sakuraba et al., 2013). In the maize (Zea mays) UROD-deficient (Les22) dominant mutant, there is a two- to threefold increase in uroporphyrin levels, while levels of Coprogen III are unaffected under HL in comparison with WT controls (Hu et al., 1998). Antisense COP RNA in tobacco leads to accumulation of Coprogen III (Kruse et al., 1995a). In the present study, Coprogen III levels in the GL1 mutant in HL conditions were significantly higher in the first 2 h and this was followed by a rapid decrease, while Proto IX content was unaffected by the shift in light conditions compared with the WT (Fig. 1e; Supplementary Table S2).
In eukaryotes and bacteria, CPO enzymes are encoded by HEMF and HEMN genes, respectively (Tanaka et al., 2011). The transcript levels of HEMF and HEMN were assayed in WT and GL1 mutant using qRT-PCR in the present study. Compared with the WT, the expression levels of HEMN were remarkably decreased, whereas HEMF showed enhanced mRNA levels after the LL/HL transfer (Fig. 4). Sequence comparison revealed that two inserted fragments were detected from the GL1 mutant sequence (Supplementary Fig. S1). Therefore, the GL1 mutant is a CPO-deficient mutant in L. indica. The HEMN gene may be responsible for the yellow leaf phenotype of the mutant.
Analysis of the complete Arabidopsis genome revealed 15 enzymes, encoded by 27 genes, for biosynthesis of Chl from glutamyl-tRNA to Chl b (Nagata et al., 2005). In higher plants, HEMF genes have been cloned from maize (Williams et al., 2006) and Arabidopsis (Ishikawa et al., 2001). However, the role of HEMN in higher plants remained unknown. In the present study, the leaf color of GL1 mutant might be caused by a deficiency of HEMN. To study the relationship of HEMN and Chl-related genes, the transcripts levels of 11 Chl-associated genes were assayed using qRT-PCR. In comparison with the WT, the expression levels of HEMA and HEMD in the GL1 mutant were decreased at all times except for t168 following LL/HL transfer (Fig. 4).
Previously, we found that HEMA and HEMD had significantly reduced mRNA levels in the mutant in different months (Li et al., 2015). HEMA1 transcripts are reduced at different levels in the rice chlorophyll synthase-deficient mutant (ygl1 mutant) at the seedling stage (Wu et al., 2007). Furthermore, HEMA1 expression is markedly downregulated under medium-light conditions in the Arabidopsis atl31/atl6 mutant (Maekawa et al., 2015). Hierarchical clustering revealed the HEMN gene shared a similar expression trend as GluRS, HEML, HEMB, and HEMC (Supplementary Fig. S1). This suggested that expression of HEMN affected expression of upstream genes.
GL1 mutant and chloroplast development
In the Chl-less mutant, chloroplast development is always hindered following LL/HL transfer. In L. vicaryi golden leaves, plastid development is arrested in HL conditions and can be derepressed by transferring to LL (Yuan et al., 2010a). Under LL conditions, the structures of Arabidopsis atl31/atl6 plastids are similar to those of WT, whereas in medium-light conditions, the Arabidopsis atl31/atl6 leaf plastids in the apex section show an almost total loss of stromal thylakoids and a reduction in grana stacks (Maekawa et al., 2015). In this study, the length and density of grana lamellae in the GL1 mutant gradually decreased with increased illumination time (Fig. 2). Following the LL/HL transfer, the grana lamellae structure was absent in the GL1 mutant at t168 (Fig. 2). This provides evidence that both GL1 and chloroplast development are highly coordinated.
The CAB gene family is a marker for chloroplast development and is tightly controlled by both light and plastid signals (Dijkwel et al., 1997). In the rice ygl1 mutant, CAB1 transcript levels are decreased at the young seedling stage (Wu et al., 2007). Similarly, CAB1 transcripts are significantly reduced in the rice vyl mutant (Dong et al., 2013). In contrast, CAB gene transcripts are not significantly reduced in the Arabidopsis atl31/atl6 double mutant (Maekawa et al., 2015). In this study, greatly decreased CAB1 mRNA levels were observed in the GL1 mutant compared with WT within the first 96 h after LL/HL transfer, and there was then a significant enhancement of mRNA levels (Fig. 5). According to the hierarchical clustering, HEMF shows a similar expression trend with CAB1 (Supplementary Fig. S1).
The GLK gene family encodes a pair of partially redundant nuclear transcription factors required for expression of nuclear photosynthetic genes and for chloroplast development (Waters et al., 2008). In an Anthurium andraeanum etiolated mutant, the expression level of GLK is lower than observed in WT (Yang et al., 2015). GLK1 is specifically downregulated in the Arabidopsis atl31/atl6 double mutant under medium-light conditions (Maekawa et al., 2015). In the present study, GLk2 showed decreased mRNA levels at t48 in the mutant compared with WT following LL/HL transfer (Fig. 5). This suggests GL1 may regulate chloroplast biogenesis through CAB1 and GLk2.
GL1 mutant and PSII stability
Chl-less mutants make valuable tools for photosynthetic studies (Havaux and Tardy, 1999). PSII functions as a light-driven water plastoquinone oxidoreductase (Liu et al., 2009). The Fv/Fm ratio is a useful relative measure of maximal quantum efficiency of PSII photochemistry, providing a simple and rapid way of monitoring environmental stress (Baker, 2007). Other fluorescence parameters, such as ETR, NPQ, and Y(II), are also important for evaluating PSII stability. In the rice vyl mutant, the values of ETR and NPQ are significantly lower than in WT, while the Fv/Fm value is similar; this is because of smaller amounts of Chl in the vyl mutant than in WT at the various growth stages (Dong et al., 2013). In a maize yellow-green mutant, values of ETR, NPQ, and Fv/Fm were markedly reduced at different growth stages compared with WT, consistent with differences in the Chl content (Zhong et al., 2015). In the present study, the Fv /Fm value in the mutant plants was lower by 53.75%, ETR by 34.58%, and Y(II) by 60.53% after 1 day in HL (7 h) (Fig. 3b; Supplementary Table S4). Meanwhile, Chl a and Chl b contents of the mutant were reduced by 47.89% and 70.26%, respectively, after 168 h in HL conditions (Table 2).
PSII is a supramolecular pigment-protein complex embedded in the thylakoid membrane. It catalyzes light-induced electron transfer from water to plastoquinone (Suorsa et al., 2006). PsbR, a fourth nuclear-encoded lumenal protein, was first identified in PSII complexes of spinach (Ljungberg et al., 1984). Absence of psbR decreases the oxygen evolution capacity of thylakoid membranes, indicating psbR is essential for photosynthetic water splitting and electron transfer in PSII (Suorsa et al., 2006; Allahverdiyeva et al., 2007). In the Chl-less tea (C. sinensis) variety “Baijiguan,” HL intensity inhibits expression of psbR, thus affecting PSII stability, chloroplast development, and chlorophyll biosynthesis (Wu et al., 2016). In our previous study, the mRNA levels of psbR were greatly suppressed in the mutant in different months (Li et al., 2015).
In the present study, the psbR gene showed decreased mRNA levels in the mutant at t96 following LL/HL transfer compared with the WT (Fig. 5). In addition, hierarchical clustering revealed that HEMF and psbR shared a similar expression trend (Supplementary Fig. S1). This provides evidence that GL1 and PSII stability is highly coordinated.
Footnotes
Acknowledgments
This work was supported by the Agricultural science and technology independent innovation fund of Jiangsu province (grant no. CX(14)2032), Sanxin forestry engineering of Jiangsu province (grant no. LYSX[2015]13), the doctorate fellowship foundation of Nanjing Forestry University, and the Natural Science Foundation of Jiangsu Province (grant no. BK20161382).
Disclosure Statement
The authors declare that there are no conflicts of interest regarding the publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.