
Abstract
Apelin, an endogenous ligand for apelin receptor (APJ), is reported to be involved in cardiomyocyte hypertrophy. In this study, we explored the mechanism of cardiomyocyte hypertrophy induced by apelin-13/APJ system. Left ventricular hypertrophy (LVH) rat model was established by constricting the abdominal aorta. Western blots were used for protein expression in LVH rats and cultured H9c2 cardiomyocytes. Transmission electron microscopy (TEM) was used to monitor morphological features of cells. In addition, the diameter and volume of H9c2 cells were detected by Scepter™ Handheld Automated Cell Counter. We found that the APJ was increased, but caveolin-1 was decreased in heart of LVH rats. In addition, caveolin-1 was suppressed by apelin-13, and this effect was reversed by APJ antagonist F13A in cultured H9c2 cardiomyocytes. Apelin-13 not only stimulated the formation of autophagolysosomes, autophagosome, and lysosomes but also increased the expression of autophagic markers Beclin-1 and LC3II/I. Besides, the increase of Beclin-1 and LC3 II/I was reversed by F13A or caveolin-1 overexpression and further enhanced by caveolin inhibitor. Furthermore, the cardiomyocyte hypertrophy index brain natriuretic peptide (BNP) induced by apelin-13 was blunt by F13A or autophagy inhibitor 3-methyladenine and further promoted by caveolin inhibitor. And caveolin-1 overexpression reduced the diameter and volume of H9c2 cells induced by apelin-13. Our study indicates that caveolin-1-autophagy pathway mediated cardiomyocyte hypertrophy induced by apelin-13/APJ system, which might provide a novel therapeutic target for cardiac hypertrophy disease.
Introduction
T
Caveolins (Caveolin-1, -2, -3), a major component protein of caveolae, regulated multiple cellular signaling pathways by redistribution and endocytosis of receptor–ligand complexes (Krajewska and Maslowska, 2004). Caveolins were largely enriched in cardiovascular system and involved in the development of atherosclerosis, cardiac hypertrophy, cardiomyopathy, as well as smooth muscle vascular cell proliferation (Williams and Palmer, 2014). Caveolin-1 was a major function protein of caveolae family. Study showed that the caveolin-1 (Cav1) knockout rats exhibited thickened left ventricular wall with a decreased systolic function (Cohen et al., 2003). Besides, loss of polymerase I and transcript release factor/Cavin-1 is sufficient to induce cardiomyocyte hypertrophy and cardiomyopathy (Takuya et al., 2016). However, the underlying mechanism(s) of caveolin-1 on cardiac hypertrophy remain poorly understood.
Autophagy was a process to maintain cellular homeostasis in response to stress, such as starvation and oxidative stress (Yang and Klionsky, 2010), which played a complicated but vital role in cardiac diseases. Our recent published article demonstrated that apelin-13/APJ induced H9c2 cardiomyocyte hypertrophy by activating PI3K/autophagy signaling pathway (Feng et al., 2015). Besides, LC3II, the autophagic marker protein, was significantly increased in siRNA interfered-caveolin-1 vascular endothelial cells (Shiroto et al., 2014). And high levels of Cav1 suppressed autophagy in astrocytes (Dias et al., 2016). These findings suggested that caveolin-1 was a negative regulator for autophagy and cardiomyocyte hypertrophy. Thus, we raised a novel hypothesis that cardiomyocyte hypertrophy induced by apelin-13/APJ might operate through the caveolin-1-autophagy pathway in this study.
Materials and Methods
Materials
DMEM, FBS, penicillin, and streptomycin were purchased from HyClone (Grand Island, NY, USA). The H9c2 cell was from ATCC (Manassas, VA, USA). Apelin-13 was purchased from Phoenix Pharmaceuticals (USA). β-cyclodextrin (β-CD) and 3-methyladenine (3-MA) were purchased from Sigma-Aldrich (USA). The caveolin-1 rabbit Antibody and the LC3A/B rabbit Monoclonal Antibody were from Cell Signaling (Beverly, MA, USA). The Beclin 1 antibody was from Santa Cruz Biotechnology (USA). The brain natriuretic peptide (BNP) antibody was from Millipore (USA). The β-Tubulin and β-Actin were from ComWin Biotech (Beijing, China).
Left ventricular hypertrophy rat model establishment
Sprague-Dawley (SD) rats (60–90 days, 200–250 g) were randomly divided into operation group and sham operation group, with six rats in each group. The abdominal aorta was constricted in operation rats. And the tail arterial blood pressure, carotid arterial blood pressure, and heart and body weight ratio were tested in sham and operation group rats.
Cell culture
H9c2 cells were cultured in DMEM containing 10% FBS in a humidified incubator with 5% CO2 in air at 37°C.
The morphology detection
H9c2 cells were seeded onto six-well culture dishes at a density of 2.5 × 105 cells/mL. After 12 h apelin-13 or not treatment, cells were fixed with 2.5% glutaraldehyde and washed with PBS thrice. Subsequently, the cells were fixed with 1% osmium tetroxide and following dehydration with an ascending series of alcohol before embedding samples in Araldite. Ultrathin sections were cut and doubly stained with uranyl acetate and lead citrate. Finally the morphological features of cells were acquired by the FEI Tecnai G2 Spirit transmission electron microscope (TEM).
Caveolin-1 cDNA transfection
pcMV6-Cav-1 (OriGene, USA) and negative pcMV6 were transiently transfected into H9c2 cells using Lipofectamine 3000 transfection reagent (Life Technologies, USA) according to the manufacturer's instructions. The cells were collected for Western blotting or other test.
Western blotting
Total protein was extracted with a radio immunoprecipitation assay lysis buffer with phenylmethanesulfonyl fluoride (9:1) and was determined using a Bicinchoninic Acid (BCA) Protein Assay Kit. Aliquots containing 40 μg of protein were subjected to dodecyl sulfate, sodium salt-polyacrylamide gel electrophoresis and were transferred to polyvinylidene difluoride membranes (Millipore Corp., Bedford, MA, USA). The membranes were incubated with primary antibodies diluted in blocking solution for 2 h and then with horseradish peroxidase-conjugated anti-rabbit or anti-mouse IgG as the secondary antibody.
The cell diameter and volume detection
The cardiomyocyte hypertrophy was assessed by the increase of cell diameter, volume, and protein content. Briefly, 1 × 104 cells/well were seeded in six-well cell culture plates and were incubated with or without any treatment. After washing, the cells were resuspended and appropriately diluted in PBS so that the concentration was ∼5 × 104−1 × 105 cells/mL, and the single-cell suspension was placed in a 1.5-mL microcentrifuge tube. The diameter and volume of the H9c2 rat cardiomyocytes were detected using a Scepter™ Handheld Automated Cell Counter (Millipore, Billerica, USA). The protein content was detected by BCA Protein Assay Kit (Beyotime).
Statistical analysis
Experiments were repeated for a minimum of three times. All values are represented as the mean ± standard error of the mean. Statistical evaluation of the data was performed by analysis of variance, followed by the Tukey posttest for comparison between groups using Prism. A value of p < 0.05 was considered to be statistically significant.
Results
Cardiac hypertrophy increased APJ and decreased caveolin-1
The left ventricular hypertrophy (LVH) rat model was established by constricting the abdominal aorta. The systolic blood pressure (SBP) of tail artery was measured at 0, 2, 4, 6, and 8 weeks, respectively (Fig. 1A). And the SBP of operation group was significantly reduced at the 8 weeks. To detect the peripheral resistance of LVH rats, we recorded diastolic blood pressure (DBP) of carotid artery at 8 weeks before being executed (Fig. 1B). Compared to sham group, the DBP was greatly elevated in operation group rats. Next, the heart weight/body weight of these execute rats was tested. The significant increase in heart weight/body weight of operation group indicated that LVH SD rat model was successfully established (Fig. 1C). Then we found that the APJ was remarkably increased in left ventricle of rats (Fig. 1D). Interestingly, corresponding with a myocardial infarction repair regulator previously described, caveolin-1 was largely decreased in cardiomyocyte hypertrophy (Fig. 1E). Collectively, the changes of APJ and caveolin-1 might play an important role in the development of cardiac hypertrophy.
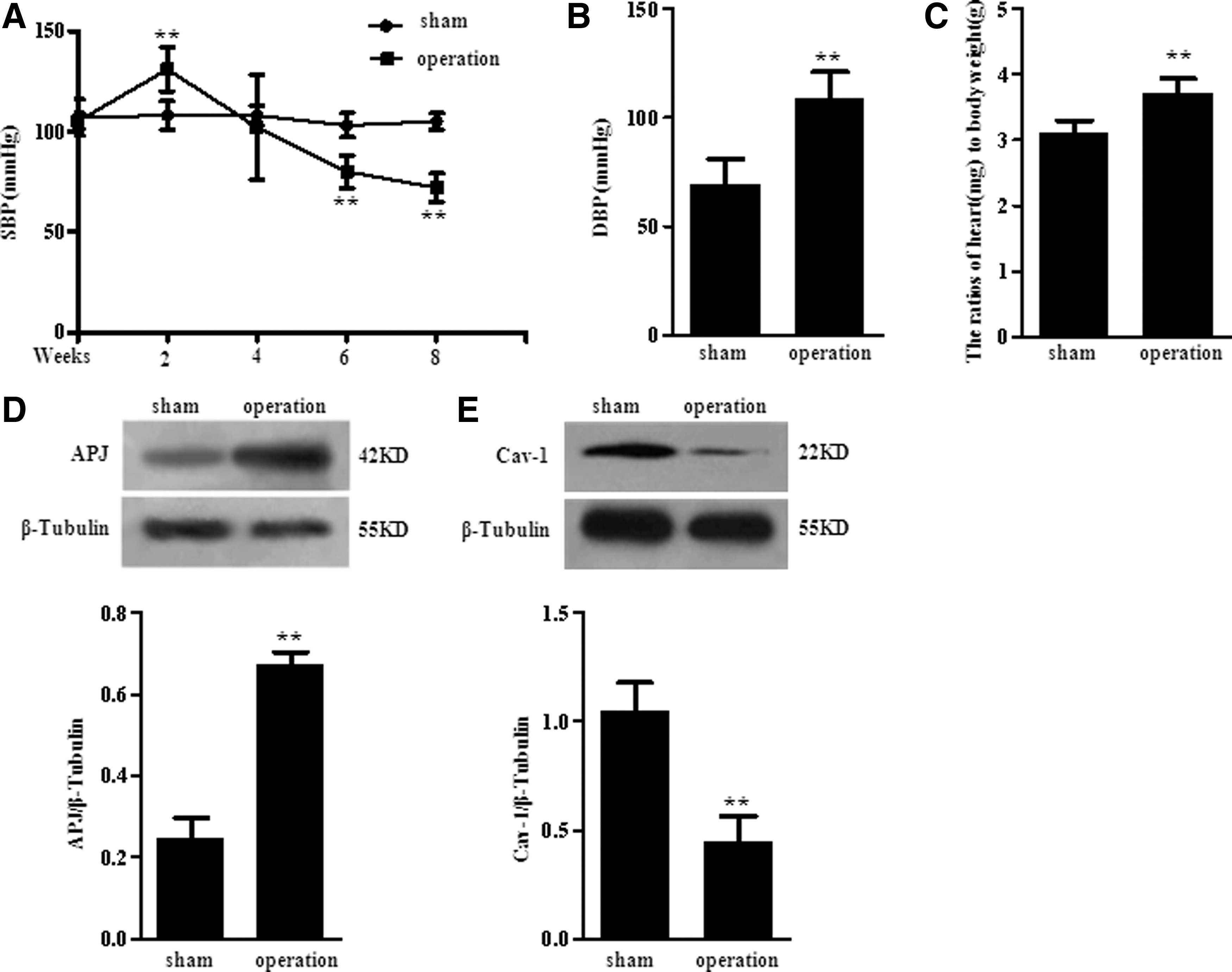
Cardiac hypertrophy increased APJ and decreased caveolin-1.
Apelin-13 suppressed caveolin-1 expression by stimulating APJ in H9c2 cardiac cells
To investigate whether apelin/APJ system could regulate caveolin-1 expression in heart, H9c2 cells were treated with apelin-13 in different concentrations (0.001, 0.01, 0.1, and 1 μM) or different times (0, 3, 6, 12, and 24 h). We observed that caveolin-1 was significantly decreased in a dose and time dependent manner (Fig. 2A, B). Moreover, the APJ specificity antagonists F13A reversed the decrease of caveolin-1induced by apelin-13 in H9c2 cells (Fig. 2C). Thus, these results showed that the apelin/APJ system could reduce caveolin-1 in H9c2 cells.
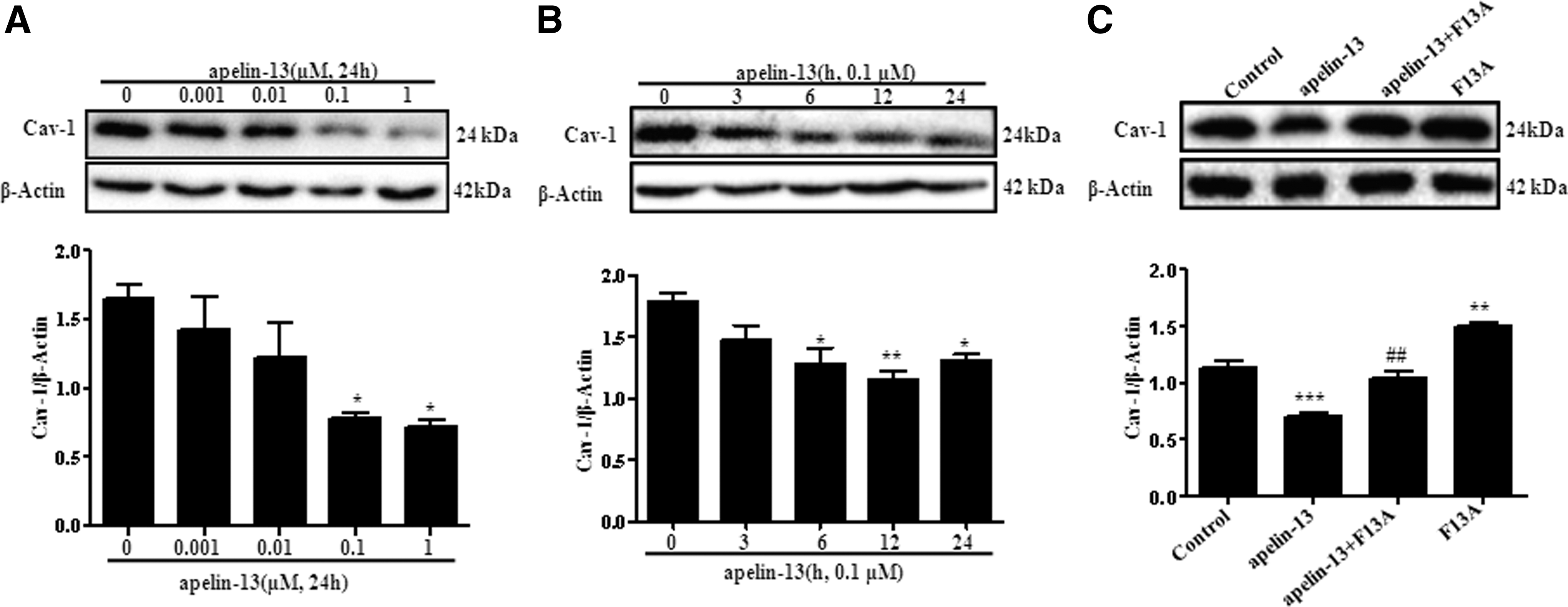
Apelin-13 suppressed caveolin-1 expression by stimulating APJ in H9c2 cells.
Apelin-13 mediated autophagy was inhibited by caveolin-1 in H9c2 cells
Our laboratory was devoted to researching the effect of apelin-13/APJ system on cardiac hypertrophy for a long time. We demonstrated that apelin-13 promoted autophagy (Feng et al., 2015). Besides, increased LC3II/I and Beclin-1 induced by apelin-13 were blocked by F13A (Fig. 3A). To further characterize the autophagy induced by apelin-13, we detected the morphological changes of H9c2 cells with TEM. TEM images showed that increased autophagolysosomes and autophagosome, as well as lysosomes, were observed after apelin-13 treatment in H9c2 cells, suggesting that apelin-13 indeed promoted autophagy (Fig. 3B).

Autophagy activated by apelin-13 was blocked by caveolin-1 in H9c2 cells.
Next, we made the caveolin-1 overexpress by transfecting pcMV6-Cav-1. The overexpression of caveolin-1 was reduced by apelin-13 (Fig. 3C). In addition, Beclin-1 and LC3II/I induced by apelin-13 were significantly downregulated by pcMV6-Cav-1 (Fig. 3D). Moreover, we found that beclin-1and LC3I/II were greatly increased after treating with a caveolae disruption β-CD, which damaged caveolin proteins. Besides, effect of β-CD on LC3I/II was further increased when treated with apelin-13 (Fig. 3E), but there was no significance for beclin-1 expression. Taken together, these observations implicated that autophagy induced by apelin-13/APJ system was prevented by caveolin-1 in cells.
Caveolin-1-autophagy pathway mediated cardiomyocyte hypertrophy induced by apelin-13
Finally, we estimated whether cardiomyocyte hypertrophy induced by apelin-13 was mediated by caveolin-1-autophagy pathway. We found that hypertrophy index BNP induced by apelin-13 was blocked by F13A (Fig. 4A). In addition, the BNP production was further enhanced after β-CD treatment (Fig. 4B). Correspondingly, autophagy inhibitor 3-MA restrained BNP induced by apelin-13 in H9c2 cells (Fig. 4C). Besides, the diameter and volume of pcMV6-Cav-1 transfected H9c2 cells were tested by Scepter Handheld Automated Cell Counter. And we found that overexpressed caveolin-1 inhibited cell diameter and volume induced by apelin-13 (Fig. 4D, E; Table 1). Combining these results, we supported that caveolin-1-autophagy pathway was involved in cardiomyocyte hypertrophy induced by apelin-13.
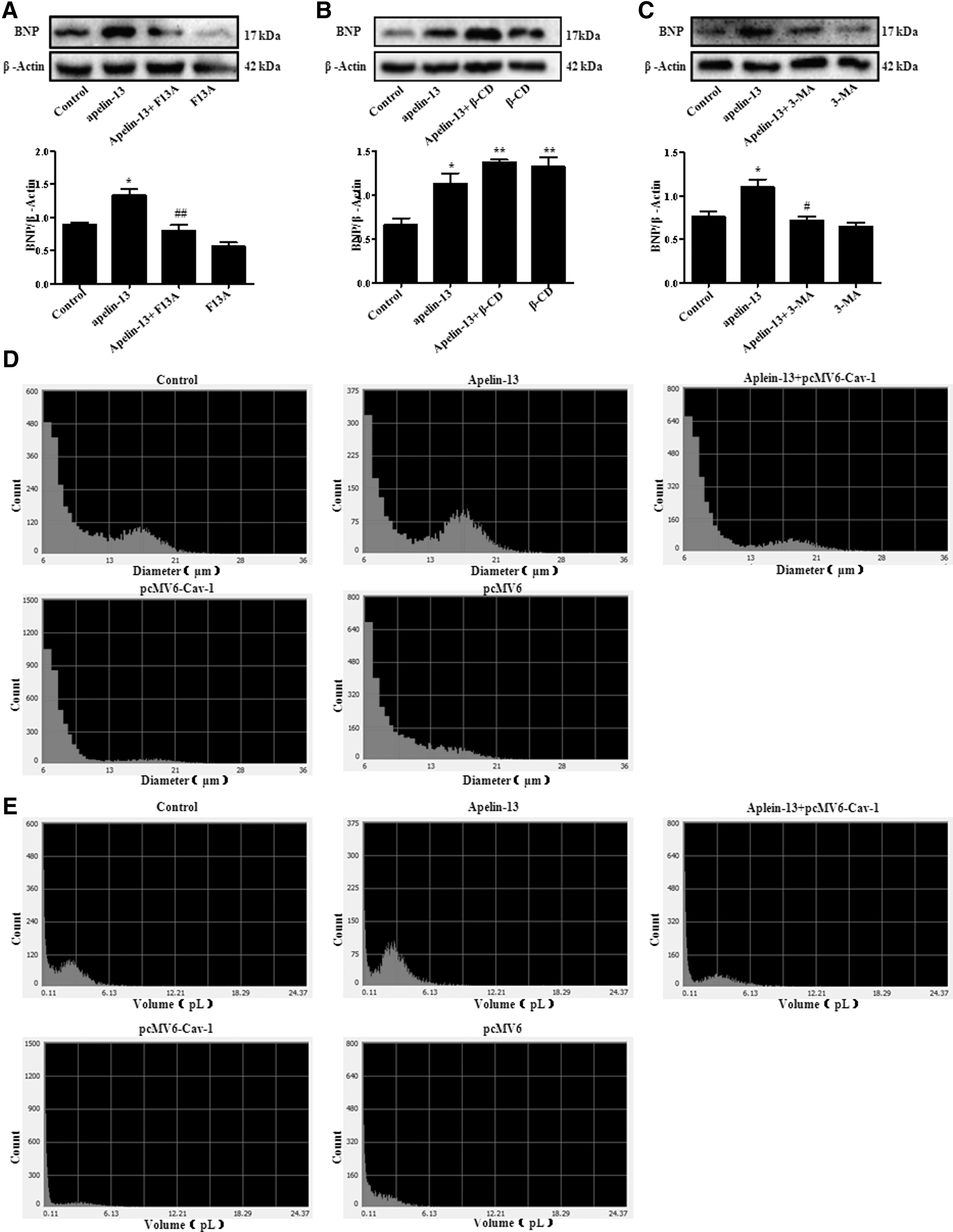
Caveolin-1-autophagy pathway mediated cardiomyocyte hypertrophy stimulated by apelin-13.
Data are mean ± standard deviation, n = 3.
p < 0.05 versus the control group.
p < 0.01 versus the control group.
p < 0.05 versus the apelin-13 group.
p < 0.01 versus the apelin-13 group.
Discussion
Apelin/APJ system played an important but complex role in cardiac hypertrophy; its functional mechanisms were studied in recent years. In this work, we identified a novel function of apelin-13 on caveolin-1, a vital regulation molecule for cardiac hypertrophy development. Besides, autophagy induced by apelin-13 was reversed by caveolin-1. More importantly, we clarified that caveolin-1-autophagy pathway participated in cardiac hypertrophy induced by apelin-13 (Fig. 5). Thus, these observations confirmed that targeting for apelin/APJ system or its downstream signal molecules would be a therapeutic strategy of cardiac hypertrophy. And agonists and antagonist of APJ provided a mechanism and therapeutic tools for managing apelin/APJ system related diseases (Iturrioz et al., 2010; Khan et al., 2010; Maloney et al., 2012; Cao et al., 2015).

Schematic illustrating: caveolin-1-autophagy pathway was involved in regulation of cardiac hypertrophy induced by apelin-13/APJ. Apelin-13/APJ system was associated with decreased caveolin-1, leading to autophagy and subsequent cardiac hypertrophy.
The hypertrophy factors and heart weight/body weight ratio were significantly increased after exogenous injection with apelin-13 in mice 14 days post-MI compared to sham group (Li et al., 2012). And Scimia et al. (2012) showed that the APJ-KO mice alleviated cardiac hypertrophy after long-term transverse abdominal aortic constriction, but apelin-KO mice still presented remarkable signs of heart failure. Their further study indicated that APJ was a stress-sensitive receptor, thus lack of APJ obviously inhibited cardiac hypertrophy induced by stretch. Our laboratory was devoted to studying the role of apelin/APJ system in cardiac hypertrophy. We reported that apelin-13 or APJ activated by static pressure promoted H9c2 cardiomyocyte hypertrophy, which was in line with Scimia et al.'s study (Xie et al., 2014a; Feng et al., 2015). However, in some conditions, apelin/APJ system could prevent the development of cardiac hypertrophy. APJ was highly homologous to angiotensin II type 1 receptor, usually apelin/APJ system resisted Angiotensin II (Ang II) action (Tatemoto et al., 1998). A study showed that cardiac cellular protein content and cellular size increased by Ang II were reversed by apelin treatment (Ye et al., 2015). Another study indicated that apelin could reduce Ang II, which induced the production of reactive oxygen species, a vital inducing factor for cardiac hypertrophy (Foussal et al., 2010; Wang et al., 2013). Besides, high fat diet-induced cardiac hypertrophy was improved by exogenous apelin. Recently, a result demonstrated that long-term (Pyr1)-apelin-13 treatment could prevent heart failure induced by pressure overload (Serpooshan et al., 2015). Taken together, it seemed that the role of apelin/APJ system in cardiac hypertrophy was homologous.
APJ as dual receptor could be activated by pressure and its nature ligands. Apelin, a novel APJ ligand Elabela/Toddler, was discovered to play an essential role in embryonic cardiac development (Chng et al., 2013; Pauli et al., 2014; Xie et al., 2014b). Moreover, the different length of apelin and stimulating factors would also affect the role of apelin in cardiac hypertrophy. In this study, we found that the APJ expression was significantly increased in heart of LVH rats. More importantly, increased cardiomyocyte hypertrophy index BNP induced by apelin-13 was blocked by F13A. These data suggested that apelin-13 induced cardiac hypertrophy by activating APJ.
Previously, the inhibition effect of caveolin-1 on cardiac hypertrophy had been proposed (Pojoga et al., 2010; Shivshankar et al., 2014); however, the exact mechanism of caveolin-1was unclear. Although there was no direct link between apelin and caveolin-1, a study by Okamoto et al. (1998) showed that caveolin-1 might be against the GPCRs APJ in vivo. In this study, we first observed that caveolin-1 was decreased in SD LVH rats after APJ overexpression. In addition, the caveolin-1 was suppressed by apelin-13 in H9c2 cells. Accordingly, we proposed that cardiac hypertrophy induced by apelin-13 might occur by restraining caveolin-1. Our previous studies indicated that autophagy participated in cardiac hypertrophy induced by apelin-13/APJ system (Xie et al., 2014a; Feng et al., 2015). The inhibitory effect of caveolin-1 on autophagy was discussed (Shiroto et al., 2014; Dias et al., 2016). Our findings suggested that beclin-1and LC3II/I induced by apelin-13 were decreased in caveolin-1 cDNA transfected H9c2 cells, but further enhanced by β-CD in H9c2 cells. There was no doubt that apelin, as a novel adipocytokine, was involved in lipid metabolism (Guo et al., 2014). In addition, caveolin-1, as a member of caveolae construct proteins, was involved in cholesterol transport (Cha et al., 2015). Therefore, apelin-caveolin-1 signaling might regulate the selective autophagy–lipophagy formation during cardiac hypertrophy.
In summary, our study demonstrated that caveolin-1-autophagy pathway mediated cardiomyocyte hypertrophy induced by apelin-13/APJ, which might provide a novel therapeutic target for cardiovascular diseases.
Footnotes
Acknowledgments
This work was supported by the grants from the National Natural Science Foundation of China (81503074, 81470434, 81670265), Hunan Provincial Natural Science Foundation (2017JJ2227), the Construct Program of the Key Discipline in Hunan Province, Hunan Province Cooperative innovation Center for Molecular Target New Drug Study (Hunan Provincial Education Department document) (2014-405), Administration of Traditional Chinese Medicine of Hunan Province (201578), and Health and Family planning commission of Hunan Province (B2015-48).
Disclosure Statement
The authors declare there are no conflicts of interest. This article does not contain any studies with human participants or animals performed by any of the authors.