
Abstract
The nuclear factor of κB (NFκB) family has been well known for its significant role in regulating the expression of numerous genes that control many biological processes. However, it is unclear whether NFκB could regulate milk synthesis. In this study, we identified NFκB1 as a critical regulator for milk synthesis in bovine mammary epithelial cells (BMECs). Gene function study revealed that NFκB1 modulates the expression of mammalian target of rapamycin (mTOR), sterol response element-binding protein (SREBP)-1c, and β4Gal-T2 for milk synthesis. Furthermore, chromatin immunoprecipitation assays showed that both methionine (Met) and estrogen (E) triggered NFκB1 to bind to gene promoters of mTOR, SREBP-1c, and β4Gal-T2 in BMECs. In addition, we confirmed that Met and E triggered NFκB1 expression and phosphorylation via phosphatidylinositol-3-kinase (PI3K) but not mTOR signaling pathway. Taken together, our study reveals that NFκB1 acts as a PI3K but not mTOR-dependent critical mediator for the transcriptional activation of signaling molecules regulating milk synthesis in BMECs.
Introduction
T
The regulation of gene expression for milk synthesis continues to be a hot research area. The mammalian target of rapamycin (mTOR) has been well described as an important protein kinase that integrates signals from amino acids and growth factors to regulate multiple biological processes required for cell growth and protein translation (Hay and Sonenberg, 2004; Wullschleger et al., 2006). Related study has confirmed that insulin-like growth factor-1 promotes protein synthesis by phosphatidylinositol-3-kinase (PI3K) signaling pathway through mTOR in bovine mammary epithelial cells (BMECs) (Burgos and Cant, 2010). Our previous study has confirmed that methionine (Met) obviously activates mTOR to enhance milk protein synthesis in BMECs (Zhang et al., 2014). Sterol response element-binding protein (SREBP) is a family of nuclear transcription factors, which are critical to the regulation of fatty acid and cholesterol biosynthetic gene expression (Horton et al., 2002). In mammals, there are three SREBP isforms including SREBP-1a, SREBP-1c, and SREBP-2. SREBP-1c preferentially activates fatty acid synthesis (Horton et al., 2002), and our recent study has shown that SREBP-1c plays a key role in regulating milk fat synthesis in BMECs (Li et al., 2014). Prolactin and estrogen (E) also notably stimulate protein synthesis in BMECs (Khudhair et al., 2015). It is still largely unknown how environmental stimuli activates mTOR and SREBP-1c, and it has not been reported which transcription factor is involved in regulating mTOR and SREBP-1c for milk synthesis.
Previous reports have shown that NFκB plays an important role in mammary gland development (Brantley et al., 2001; Cao and Karin, 2003). However, whether NFκB and which member of NFκB plays a key role in regulating milk synthesis has not been reported. In this study, NFκB1 is identified as a positive regulator for milk synthesis in BMECs. We further confirm that p-NFκB1 binds to the promoters of mTOR, SREBP-1c, and β4Gal-T2, leading to the transcriptional activation of these genes, thereby promoting milk synthesis in BMECs. We also reveal that both Met and E trigger NFκB1 phosphorylation through PI3K but not mTOR signaling pathway.
Materials and Methods
Culture and treatments of BMECs
Primary BMECs were isolated from the mammary gland tissues of a mid-lactation Holstein dairy cow as previously described (Li et al., 2014; Zhang et al., 2014). Cells were cultured in Dulbecco's modified Eagle's medium-F12 (DMEM-F12; Gibco, Carlsbad, CA) supplemented with 10% fetal bovine serum (FBS; Gibco), 100 U/mL penicillin, and 100 μg/mL streptomycin. For experimental assays, cells in the logarithmic growth phase were cultured in cell flasks or plates at 37°C in a humidified atmosphere containing 5% CO2. Before nutrient treatments, culture medium was replaced with DMEM-F12 without FBS. To investigate the influence of Met and E on NFκB1 expression, cells in DMEM-F12 medium (containing 0.12 mM Met) were treated with Met (0.6 mM) or E (2.72 × 10−2 μg/mL) for 24 h, and processed for further experiments. Wortmannin and rapamycin were used as inhibitors of PI3K and mTOR, respectively. One hour before Met or E treatment, wortmannin or rapamycin of different concentrations was added to cells, respectively.
Immunofluorescence assays
BMECs were seeded on sterile coverslips in a six-well culture plate until a confluency of 70%–80%. Cells were fixed with precooled carbinol for 10 min and then blocked in Tris buffer saline (TBS) buffer with 5% bovine serum albumin and 0.1% Triton X-100 for 1.5 h at 37°C. Cells were incubated with primary antibodies overnight at 4°C. The primary antibodies included NFκB1 (Cell Signaling Technology, Beverky, MA) and p-NFκB1 (Ser337; Abcam Technology, Cambridge, MA). After washing with TBS with 0.1% Tween (TBST), the samples were incubated with second antibody conjugated with Alexa Fluor-488 or Alexa Fluor-647 (Bioss, Beijing, China) for 1 h at 37°C and then incubated with 4,6-diamidino-2-phenylindole (DAPI) for 10 min. Images were captured under a laser scanning confocal microscope (Leica Microsystems, Heidelberg, Germany).
Transfection of pGCMV-IRES-EGFP-NFκB1
The full-length of NFκB1 coding region (NCBI accession number: NM_001076409) was amplified by the primer sequences as follows, sense: 5′-CGGAATTCATGGCAGAAGACGACCCGTATTT-3′ (EcoR I); antisense: 5′-ACGCGTCGACAATTTTGCCTTCTATAGGTCCTTCC-3′ (Sal I). The NFκB1 gene segment was finally subcloned into pGCMV-IRES-EGFP vector (GenePharma, Shanghai, China). The recombinant plasmid was subjected to sequencing for verification. BMECs were plated in six-well plates until a confluence of 70%–80%. They were transfected with the pGCMV-IRES-EGFP-NFκB1 or pGCMV-IRES-EGFP using Lipofectamine 2000 (Invitrogen, Carlsbad, CA) according to the manufacturer's instruction. Cells were harvested after 24 and 48 h post-transfection for quantitative polymerase chain reaction (qPCR) and western blot analysis, respectively.
Tranfection of small interfering RNAs
The NFκB1 small interfering RNA (siRNA) and negative control RNA were purchased from GenePharma. The sequences of NFκB1 siRNA were as follows: si-NFκB1, sense 5′-GGAAACGACAGAAGCUCAUTT-3′, antisense 5′-AUGAGCUUCUGUCGUUUCCTT-3′. Cells were transfected with siRNA against NFκB1 or negative control siRNA using Lipofectamine 2000 according to the manufacturer's protocol. Nontreated cells were also included as control groups. The transfection efficiency was assessed by qPCR after 24 h and western blot after 48 h.
qPCR assays
Total RNA from BMECs was extracted with TRIzol Reagent. The quantity and purity of RNA samples were analyzed by ultraviolet spectrophotometer. One microgram of total RNA was reverse transcribed into cDNA using PrimeScript RT Reagent kit (Takara, Tokyo, Japan). qPCR assays were performed using an ABI PRISM 7300 Real-Time PCR System (Applied Biosystem, Foster City, CA).
The primer sequences for the target genes were as follows: NFκB1, sense 5′-CCGTAGGAGTAAAGGAAGAGAAC-3′, antisense 5′-CTGATTATGAAGGTGGATGATTGC-3′; IκBα, sense 5′-ATGAAGGACGAGGAGTATGAGC-3′, antisense 5′-GTTGTTCTGGAAGTTGAGGAAGG-3′; mTOR, sense 5′-TGCCTTCACAGATACCCAG-3′, antisense 5′-TCAGACCTCACAGCCACAG-3′; SREBP-1c, sense 5′-CAGTAGCAGCGGTGGAAGTG-3′, antisense 5′-GAGAGACAGAGGAAGACGAGTG-3′; β-casein, sense 5′-GTACCTGGTGAGATTGTGG-3′, antisense 5′-CTGTTTGCTGCTGTTCCT-3′; β4Gal-T2, sense 5′-CGCTACTGGCTCCACTACCTGC-3′, antisense 5′-CCTTGAGTGCCTCTAGGAAACCC-3′; β-actin, sense 5′-AAGGACCTCTACGCCAACACG-3′, antisense 5′-TTTGCGGTGGACGATGGAG-3′. The PCR cycle was as follows: 95°C for 5 min, 40 cycles of 95°C for 10 s and 60°C for 34 s and followed by one cycle at 95°C for 15 s, at 60°C for 60 s and at 95°C for 15 s. The relative level of mRNA expression was analyzed by the 2−ΔΔCt method. All reactions were performed in triplicate.
Western blot analysis
Cells were washed with cold phosphate buffer saline and lysed with a lysis buffer (Beyotime, Shanghai, China) at 4°C. After centrifugation, samples containing 30 μg of proteins were subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Proteins were then transferred to nitrocellulose membranes blocked in 5% nonfat milk dissolved in TBST and then incubated with primary antibodies at 4°C overnight. Membranes were washed with TBST and then incubated with horseradish peroxidase (HRP)-conjugated secondary antibodies (ZSGB-Bio, Beijing, China) for 1 h at 37°C. ECL Substrate (SageBrightness, Sagecreation, Beijing, China) was used for the detection of HRP on immunoblots.
The primary antibodies were as follows: SREBP-1c, Akt1, p-Akt1 (Thr308), S6K1, p-S6K1 (Thr389), and β-actin (Santa Cruz Biotechnology, Santa Cruz, CA); NFκB1, IκBα, p-IκBα (Ser32/36), mTOR, and p-mTOR (Ser2448; Cell Signaling Technology); p-NFκB1 (Ser337; Abcam Technology); β-casein (Abbiotec, San Diego, CA).
Determination of triglyceride and lactose
Following different treatments or transfection, cell-free culture medium was collected to determine the contents of secreted triglyceride (TG) and lactose using TG GPO-POD Assay Kit (Applygen Tech, Inc., Beijing, China) and Lactose/
BODIPY staining of lipid droplets
Cells were fixed with 3% formaldehyde for 20 min, then stained with 4, 4-difluoro-1, 3, 5, 7, 8-pentamethyl-4-bora-3a, 4a-diaza-s-indacene (BODIPY 493/503; Invitrogen) in a working solution of 1 μg/mL for 10 min and then stained with DAPI for 15 min. Images were captured using a leica TCS-SP2 AOBS confocal microscope.
Chromatin immunoprecipitation assays
Chromatin immunoprecipitation (ChIP) assay was carried out using an EpiQuikTM Chromatin Immunoprecipitation Kit (Epigentek, Farmingdale, NY) and an antibody against p-NFκB1 (Ser337; Abcam Technology). Anti-RNA polymerase antibody was used as a positive control and immunoglobulin G as a negative control. The amplification of predicted sequence using these primers were verified by ChIP-PCR. ChIP-qPCR was applied to determine the percentage of input that was immunoprecipitated.
The predicted κB sites in gene promoters of mTOR, SREBP-1c, and β4Gal-T2 are GGGGCACCCC, GGGCCTGTCC, and GGGCCCTCCC, respectively. The qPCR primers to amplify the predicted κB sites are as follows: mTOR, sense 5′-GCGGCGGAATGTTCAGAC-3′, antisense 5′-CCA GAAACGCACGATAGGCT-3′; SREBP-1c, sense 5′-CCC TGAGCAGGACCACTTGG-3′, antisense 5′-TGACATTTG TGGCGCACTTTG-3′; β4Gal-T2, sense 5′-AGAGTGAAG ATGGGTGATTAGATG-3′, antisense, 5′-CAGCACGAAG TGAGTGTAGG-3′.
Statistical analysis
Data are presented as mean ± standard error for each group from three independent experiments. Statistical analyses were performed using one-way analysis of variance with SPSS 17.0 software (SPSS, Inc., Chicago, IL). Tukey post hoc tests were used for analyzing differences between the means of individual groups. Differences with p < 0.05 and p < 0.01 were considered statistically significant and extremely significant, respectively.
Results
Overexpression of NFκB1 upregulates milk synthesis in BMECs
To investigate whether NFκB1 affected milk synthesis in BMECs, we detected the expression of signaling molecules related to milk synthesis in NFκB1-overexpressing BMECs. From the observations, we found that NFκB1 overexpression obviously increased the mRNA levels of NFκB1, IκBα, mTOR, SREBP-1c, β4Gal-T2, and β-casein compared with the empty vector group (Fig. 1A). The protein levels of NFκB1, p-NFκB1, IκBα, mTOR, p-mTOR, SREBP-1c, and β-casein were all notably upregulated, whereas p-IκBα expression was found decreased, showing no significant difference (Fig. 1B, C). TG and lactose secretion (Fig. 1D, F), and lipid droplet accumulation (Fig. 1E) were all increased in NFκB1-overexpressing group. These data reveal that NFκB1 overexpression positively upregulates milk synthesis in BMECs.
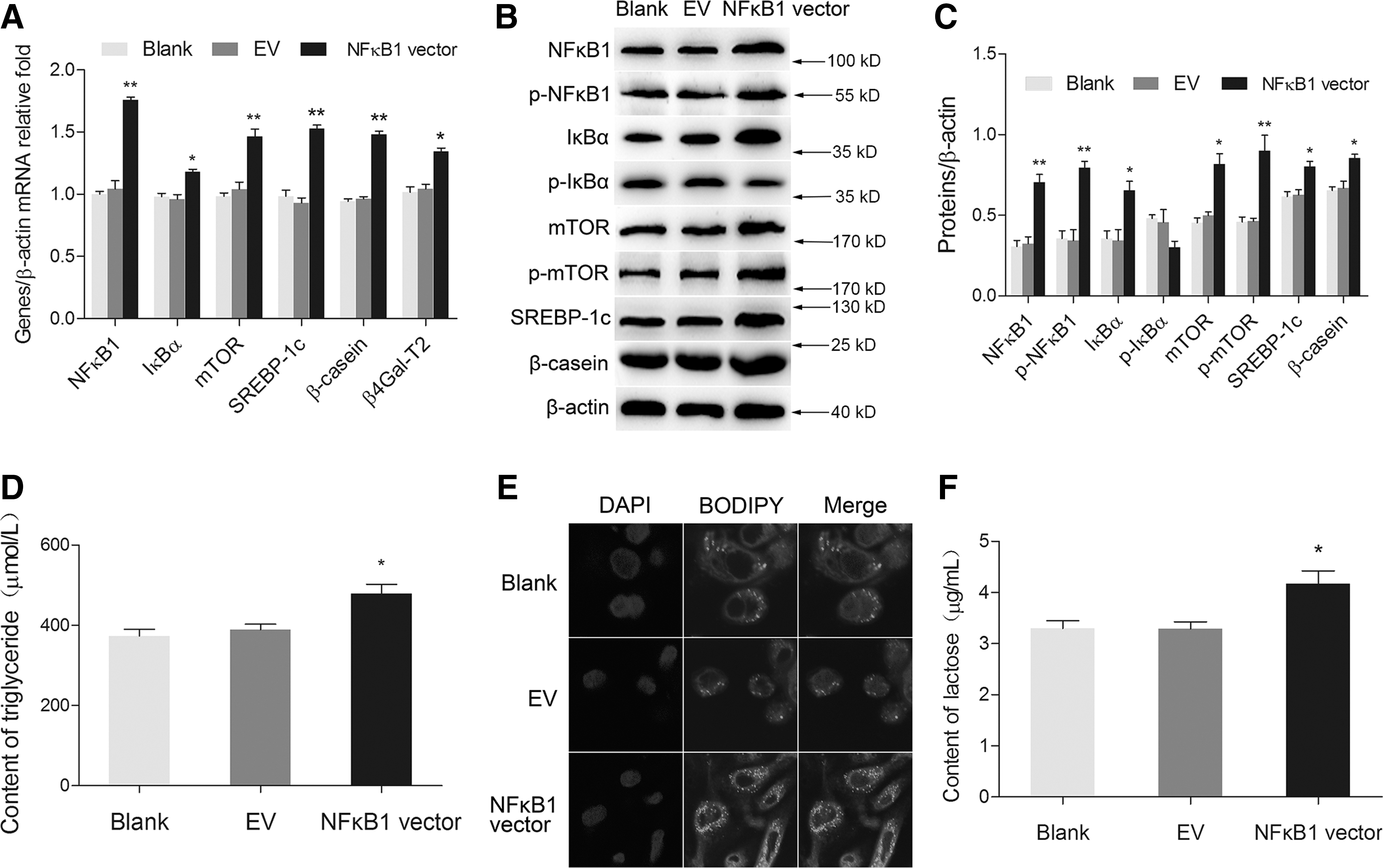
Overexpression of NFκB1 upregulated milk synthesis in BMECs.
Silencing of NFκB1 suppresses milk synthesis in BMECs
Silencing of NFκB1 by siRNA transfection obviously decreased mRNA levels of NFκB1, IκBα, mTOR, SREBP-1c, β4Gal-T2, and β-casein compared with the negative control group (Fig. 2A). The protein levels of NFκB1, p-NFκB1, IκBα, mTOR, p-mTOR, SREBP-1c, and β-casein were all significantly downregulated. However, p-IκBα expression was found upregulated (Fig. 2B, C). TG and lactose secretion (Fig. 2D, F), and lipid droplet accumulation (Fig. 2E) were all decreased in NFκB1 silencing group. These results provide further evidence that NFκB1 positively regulates milk synthesis in BMECs.
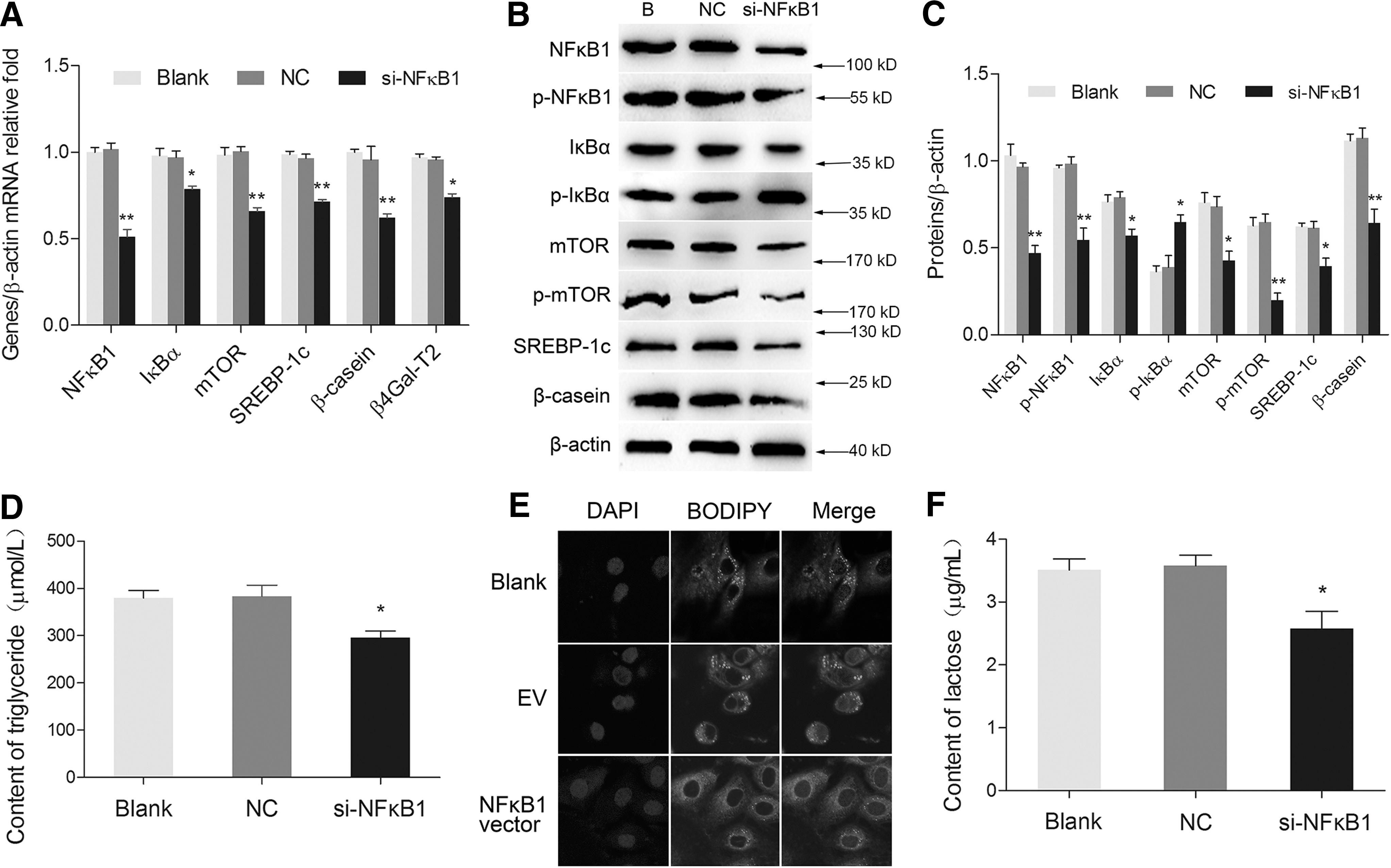
Silencing of NFκB1 suppressed milk synthesis in BMECs.
NFκB1 controls the transcriptional activation of milk synthesis
To explore the mechanism how NFκB1 mediates milk synthesis in BMECs, we carried out ChIP-PCR and ChIP-qPCR assays. We first predicted the consensus κB binding sites (GGGRNNYYCC, R: purine, Y: pyrimidine, N: any base) in the gene promoters of mTOR, SREBP-1c, and β4Gal-T2 (Fig. 3A). The predictions were verified by ChIP-PCR using the antibody against p-NFκB1. ChIP-qPCR assays were then applied to detect the enrichment of p-NFκB1 binding sequences in BMECs treated with Met or E. The enrichment of these gene promoters in the treated groups were all notably increased compared with the blank group (Fig. 3B). Moreover, the mRNA levels of NFκB1 and its target genes were also promoted by Met or E stimulation (Fig. 3C), consistent with the ChIP-qPCR result. Western blot analysis showed an obviously increasing expression of NFκB1 and IκBα by the treatment of Met and E, however, p-IκBα expression was found downregulated (Fig. 3D). Immunofluorescence assays showed that Met and E enhanced the nuclear localization of both NFκB1 (Fig. 3E) and p-NFκB1 (Fig. 3F). These results reveal that NFκB1 responses to environmental stimuli to control its target gene expression for milk synthesis.
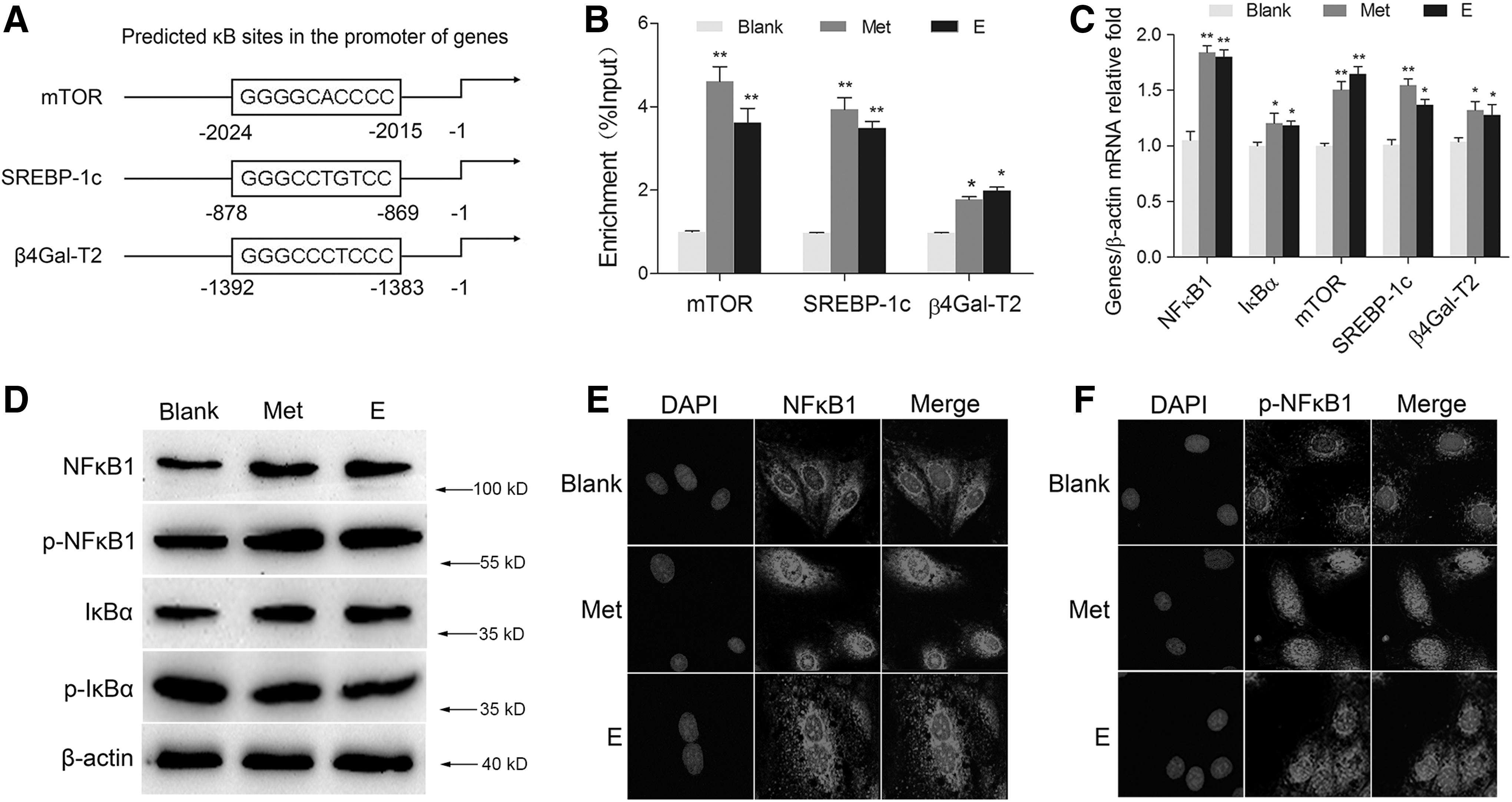
Met and E increased the NFκB1-mediated gene expression.
NFκB1 is a PI3K but not mTOR-dependent critical mediator for milk synthesis
To detect the mechanism by which environmental stimuli such as Met and E activate NFκB1, wortmannin and rapamycin were used as the inhibitors of PI3K and mTOR signaling pathways, respectively. We first screened the optimal concentration of each inhibitor to obtain the best inhibitory effect on PI3K and mTOR in BMECs. Western blot analysis showed that wortmannin at the concentration of 400 nM significantly repressed the protein level of p-Akt1 (which level represented the activity of PI3K) and markedly suppressed the protein levels of NFκB1 and p-NFκB1 (Fig. 4A). Rapamycin at the concentration of 6 and 8 nM both notably reduced the expression of p-S6K1 (which level represented the activity of mTOR), however, the protein levels of NFκB1 and p-NFκB1 had no obvious change after rapamycin treatment (Fig. 4B). These results suggest that NFκB1 is downstream from PI3K but not mTOR signaling pathway. After wortmannin treatment, both NFκB1 and p-NFκB1 levels induced by Met and E were almost abrogated (Fig. 4C, D). These above data reveal that amino acids and hormones trigger NFκB1 phosphorytion by activating PI3K but not mTOR signaling pathway.
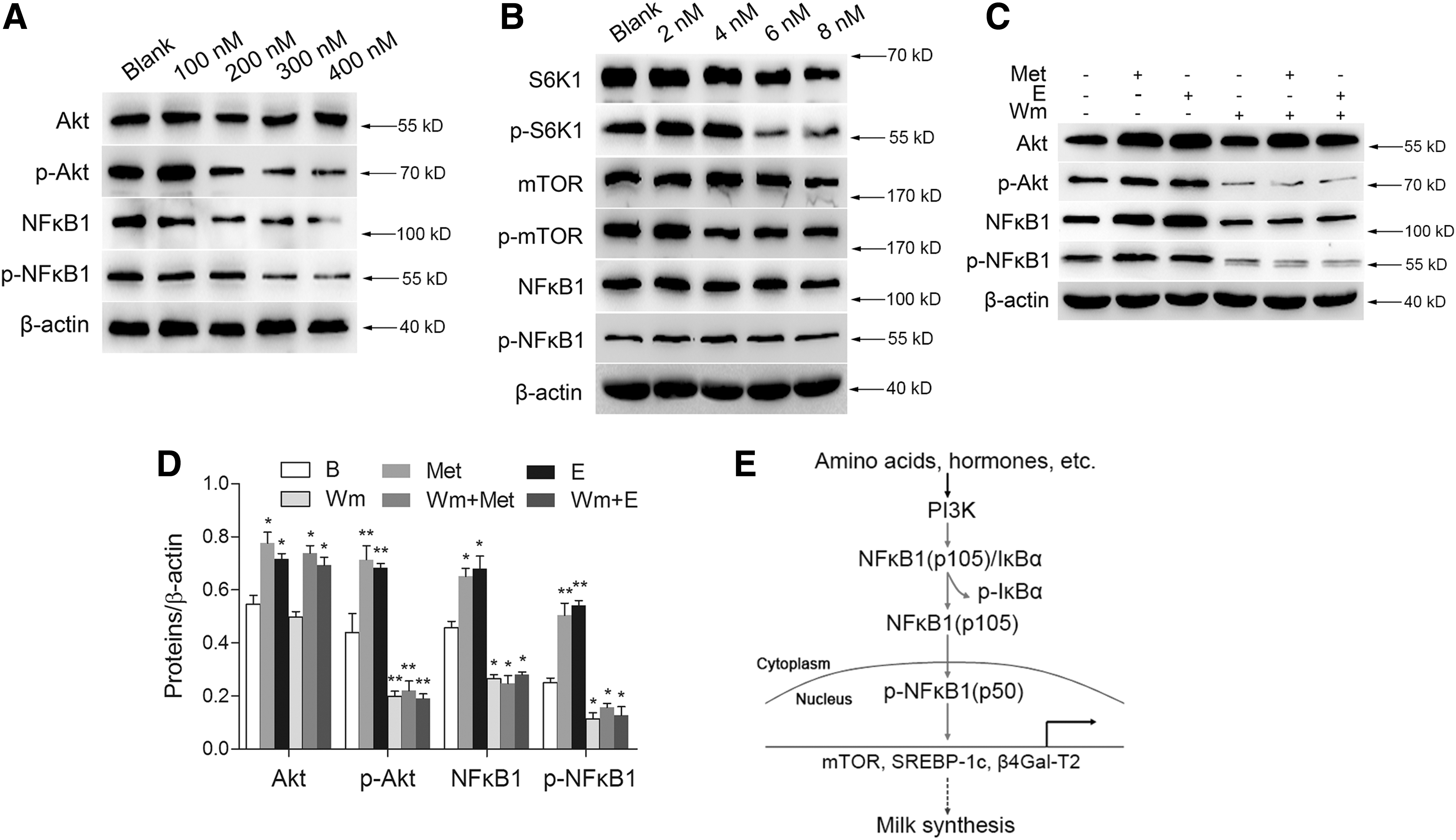
The stimulation of Met and E on NFκB1 phosphorylation is dependent on PI3K but not mTOR signaling pathway.
Discussion
The aim of this study is to reveal whether NFκB1 is a critical mediator for the transcriptional activation of milk synthesis stimulated by amino acids and hormones in BMECs. The regulation of gene expression for milk synthesis depends on nutritional and hormonal regulation (Qian and Zhao, 2014). Previous studies in our lab have showed that Met stimulation increases β-casein expression in BMECs (Yu et al., 2014). E also can stimulate the secretion of β-casein and TG in BMECs (Khudhair et al., 2015). In this study, we found an obvious increasing expression and nuclear translocation of NFκB1 after Met and E treatments. Met and E also promote NFκB1 phosphorylation and its nuclear localization. These data suggest that NFκB1 is associated with milk synthesis induced by amino acids and hormones.
Our gene function studies showed that NFκB1 positively regulates mTOR, SREBP-1c, and β4Gal-T2 signaling pathways. We detected the mRNA levels of these signaling molecules at 24 h post transfection of NFκB1 overexpression vector or siRNA targeting NFκB1, and the protein levels of these signaling molecules were detected at 48 h post transfection. The consistent results of qPCR and western blot analysis at different time points reflected the effects of NFκB1 on milk synthesis at transcriptional and translational level. We further determined the secretion of milk fat and lactose at 48 h post transfection, and the results were consistent with western blot analysis. Our data revealed that NFκB1 positively regulated milk synthesis in BMECs.
β-1,4-galactosyltransferase (β4GalT) is involved in lactose synthesis. β4GalT and α-lactalbumin assemble the lactose synthetase, which catalyzes to transfer galactose to N-acetylglucosamine and results in the generation of β4-N-acetyllactosamine (Galβ4GlcNAc) (Turkington and Hill, 1969; Charron et al., 1998). Previous report found that both β4Gal-T1 and -T2 show similar α-lactalbumin sensitive lactose synthetase activity (Amado et al., 1999; Shahbazkia et al., 2012). In this research, we observed an obvious increase of mRNA expression of β4GalT-2 in BMECs treated with Met or E, and overexpression and silencing of NFκB1 also influence the mRNA expression of β4GalT-2, consistent with the change of lactose contents. Our experimental data provide a new evidence that β4Gal-T2 is involved in lactose synthesis and regulated by NFκB1.
Our study showed that both NFκB1 overexpression and nutrient treatments upregulated IκBα expression by promoting its transcription, however, p-IκBα level was reduced. Reversely, silencing of NFκB1 markedly repressed IκBα expression, which resulted in an increasing level of p-IκBα. A previous study has demonstrated that a κB binding site exists in the promoter of IκBα. This study also showed a mutual regulation of NFκB and IκBα. This feedback loop ensures the maintenance of cellular homeostasis upon environmental stimuli (Brown et al., 1993). We speculate that the protein level of IκBα is likely to be sufficient to combine with NFκB1, and IκBα proteins free from NFκB1 might be phosphorylated for degradation.
In this study, we also identified the target genes of NFκB1 for milk protein synthesis. Previous study in our lab has confirmed that Met and E trigger NFκB1 to bind to the Tudor staphylococcal nuclease to promote milk protein and fat synthesis in BMECs (Ao et al., 2015). In our study, we identified the interaction between p-NFκB1 and its target gene promoters of mTOR, SREBP-1c, and β4Gal-T2. ChIP-qPCR analysis showed an obviously increasing binding of p-NFκB1 to these promoters by the treatment of Met or E. The results of ChIP assays showed an obvious increasing enriched binding of NFκB1 target genes after the treatment with Met and E, which was consistent with qPCR analysis. It appears that there is a general increase of gene expression by environmental stimuli, rather than specific to NFκB1 target genes. In our study, we first predicted the κB sites in gene promoters of mTOR, SREBP-1c, and β4Gal-T2. The predicted κB sites in these gene promoters showed high similarity with the consensus κB binding sites (GGGRNNYYCC), which indicate that these genes are specific to NFκB1. Our predictions were verified by ChIP-PCR. Furthermore, Met and E triggered NFκB1 gene expression and activation, whereas NFκB1 knock down significantly reduced its target gene expression and impaired corresponding signaling pathways, revealing that environmental stimuli on milk synthesis of BMECs is largely via the NFκB1 pathway. These data reveal that NFκB1 directly controls the gene transcription of mTOR, SREBP-1c, and β4Gal-T2 for milk synthesis in BMECs.
The PI3K pathway regulates numerous biological processes through its ability to regulate cell growth, apoptosis, and responses to nutrients (Vivanco and Sawyers, 2002). As a serine/threonine kinase, Akt acts downstream of PI3K and has a wide range of substrates (Ma and Blenis, 2009). Previous study has demonstrated that mTOR downstream from Akt functions through IKKα phosphorylation to control phosphorylation of RelA (p65) of NFκB (Dan et al., 2008). In this study, we detected the mechanism by which NFκB1 senses Met and E. Western blot analysis showed that the expression of NFκB1 and p-NFκB1 were markedly decreased when BMECs were treated with wortmannin, whereas rapamycin exhibited no obvious inhibitory effect on NFκB1. We further showed that the stimulation on NFκB1 expression and phosphorylation by Met and E were almost abolished when wortmannin was added. These results showed that Met and E trigger NFκB1 phosphorylation via PI3K but not mTOR signaling pathway, revealing the different mechanism of NFκB1 activation from RelA (p65).
Conclusions
Taken together, our study confirms that NFκB1 is a critical regulator for milk synthesis in BMECs. Furthermore, we reveal the molecular mechanism that Met and E activate NFκB1 via PI3K but not mTOR signaling pathway, and p-NFκB1 promotes milk synthesis by targeting mTOR, SREBP-1c, and β4Gal-T2 (Fig. 4E).
Footnotes
Acknowledgments
This study was financially supported by the National Natural Science Foundation of China (No. 31472162), and High Technology Project of the Ministry of Science and Technology of China (863 Project, No. 2013AA102504-03).
Authors' Contributions
In this study, X.H. performed BMEC culture, cell treatments, ChIP assays, data analysis, and co-drafting of the article; Y.Z. achieved immunofluorescence experimental assays and western blot; M.Z. performed qRT-PCR and determination of TG and lactose contents immunofluorescence experimental assays; X.Y. and M.L. carried out western blot and data analysis; X.G. developed the conception and design of this research, supervised the project, and wrote the article.
Disclosure Statement
No competing financial interests exist.