
Abstract
Increasing evidence has suggested that long noncoding RNAs (lncRNAs) play critical roles in cancer development. Nasopharyngeal carcinoma (NPC) is a disease with high incidence. Although remarkable progress has been made in understanding the molecular mechanism and therapy strategies in NPC, the potential involvement of lncRNAs in NPC remains largely unknown. In this study, we identified that lncRNA cancer susceptibility candidate 9 (CASC9) is highly expressed in NPC tissues, which facilitates cell growth and is correlated with a poor prognosis of cancer patients. The underlying molecular mechanism revealed that CASC9 interacts with HIF1α and enhances the stabilization of HIF1α. Activation of HIF1α by overexpressed CASC9 promotes the glycolysis and tumorigenesis of NPC cells. Downregulation of CASC9 significantly inhibits NPC cancer cell growth. Collectively, our results illustrated the oncogenic role of CASC9 in promoting the progression of NPC through regulating HIF1α, which imply that modulation of CASC9 expression may be a promising target in cancer therapy.
Introduction
N
Recently, long noncoding RNAs (lncRNAs), which were defined as RNAs longer than 200 nucleotides that are not translated into protein, were reported as key regulators in embryonic growth, cell proliferation, and differentiation by regulating transcriptional, posttranscriptional, and epigenetic mechanisms (Ponting et al., 2009; Wang et al., 2014; Khorkova et al., 2015; Yuan et al., 2016). Accumulating evidence has suggested that dysregulation of lncRNAs play an important role in a variety of human cancers (Orom et al., 2010; Geisler et al., 2012; Shi et al., 2015; Schmitt and Chang, 2016). Among these known lncRNAs, H19 is identified as a tumor suppressor that possesses the function of gene imprinting and is highly expressed in breast, endometrial, and bladder cancers (Lottin et al., 2002; Zhang et al., 2016). Ewing sarcoma-associated transcript 1 (EWSAT1), a kind of lncRNA located on chromosome 15, has oncogenic potential (Song and Yin, 2016; Sun et al., 2016). Upregulated expression of EWSAT1 was observed in esophageal adenocarcinoma and NPC (Marques Howarth et al., 2014; Song and Yin, 2016). Depletion of EWSAT1 inhibits malignant cancer cell proliferation, cell cycle distribution, and invasion (Song and Yin, 2016). These results indicated the importance of lncRNA species in cancer research.
Cancer susceptibility candidate 9 (CASC9) was reported as an esophageal squamous cell carcinoma (ESCC)-associated lncRNA and a higher level of CASC9 expression was observed in ESCC patients (Pan et al., 2016). Knockdown of CASC9 inhibits cell migration and invasion of ESCC cells, suggesting the oncogenic potential of CASC9 in esophageal carcinogenesis (Pan et al., 2016). However, the role of CASC9 in other type of cancers and the detailed molecular mechanism of CASC9 in cancer progression remain largely unknown.
In this study, we found that CASC9 is highly expressed in human NPC tissues. Higher expression level of CASC9 facilitates cell growth and correlates with a poor prognosis of cancer patients. We also found that CASC9 functions as an oncogenic regulator via activation of HIF1α, which finally reprograms the glycolysis metabolism and promotes the cancer cell growth. Downregulation of CASC9 inhibits tumorigenesis suggesting the application of CASC9 in cancer diagnosis.
Materials and Methods
Cell culture and clinical samples
Human NPC cell lines were cultured in RPMI-1640 (Gibco, Life Technologies, Carlsbad, CA) supplemented with 10% FBS (Gibco) at 37°C in a 5% CO2 humidified atmosphere. Human NPC tissue samples were kindly provided by the Cangzhou People's Hospital. Written informed consent was obtained from all the patients before biopsy. The investigation was approved by the Ethics Committee of Cangzhou Central Hospital.
Western blot analysis
Total proteins were extracted from cultured cells with the NP-40 buffer (50 mM Tris-HCl, pH 8.0, 150 mM NaCl, 1% NP-40, 1 mM EDTA). Proteins were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to the nitrocellulose filter membrane (Millipore, Billerica, MA). The membrane was first blocked with 5% BSA for 1 h and then sequentially incubated with the primary antibody for 2 h and secondary antibody for 1 h at room temperature. The signal was visualized with KeyGEN Enhanced ECL detection kit according to the manufacturer's instructions (KeyGEN, Nanjing, China).
RNA extraction and quality control
Total RNA was extracted from the NPC tissue samples using TRIzol reagent (Invitrogen). The amount and quality of RNA were evaluated with the NanoDrop ND-2000 spectrophotometer (Thermo Scientific, Rockford, IL). Reverse transcription was performed with 1 μg RNA according to the manufacturer's instructions. Quantitative reverse transcription-PCR was conducted using Platinum SYBR Green Master Mix (Invitrogen, Grand Island, NY) on the Roche LightCycler® 96 (LC96) real-time PCR platform. β-actin was used as the normalization control, and the relative expression abundance of CASC9 was calculated with the 2−ΔΔCt method. Primer sequence of CASC9 and β-actin were as follows: CASC9, 5′-AGATGAAGCCGGTACCTCAGAT and 5′-TCACTTTAAAGAGGGAGAGGAG; β-actin, 5′-AAGTGTGACGTGGACATCCGC and 5′-CCGGACTCGTCATACTCCTGCT.
Cell proliferation assay
CNE-1 cells transfected with CASC9 were plated in 96-well plates with 1000 cells per well. After 1, 2, 3, 4, 5, and 6 day, 0.05 mg mL−1 MTS (3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxypheyl)-2-(4-sulphophenyl)-2H-tetrazolium, inner salt (Sigma) regent (Promega) was added to the wells. The plates were incubated at 37°C for 3 h, after which the abundance of the solution in each well was measured at 492 nm using an ELX800 spectrophotometric plate reader (Bio-Tek, Winooski, VT).
Colony formation analysis
Soft agar colony formation assay was performed with CNE-1 cells stably expressing CASC9. Cells were harvested and pipetted well to become single-cell suspension in complete culture media with the concentration at 1 × 106/mL. Ten percent FBS DMEM containing 0.75% agar were put into 60 mm culture dish in the flow hood. After the agar solidified, the top layer containing 3 × 104 cells in 3 mL of 10% FBS DMEM were added to the well. Mark the dish and incubate it at 37°C for 3 weeks, and then the photographs of the colonies were taken.
Cell cycle and apoptosis assay
For the cell cycle arrest analysis, cells were harvested by trypsinization and fixed in 75% ethanol at 4°C overnight. And then cells were incubated with 1 mg mL−1 RNase A at 37°C for 30 min, stained with 50 μg mL−1 propidium iodide, and exposed to DNA content analysis by FACS. Cell apoptosis assay was performed with the AnnexinV-FITC Apoptosis Detection kit (Life Technology) according to the manufacturer's instructions.
Measurement of glucose uptake
Cells were plated in six-well plate with a concentration of 3 × 105 cells/mL at 24 h before the experiment. Cells were washed firstly with precold PBS, and the glucose uptake was performed by incubating cells in 2 mL of Krebs-Ringer-HEPES (KRH) buffer (25 mM HEPES, pH 7.4, 5 mM KCl, 1.2 mM MgSO4, 120 mM NaCl, 1.3 mM CaCl2, and 1.3 mM KH2PO4) containing 1 μCi of [3H]-2-deoxyglucose (PerkinElmer Life Sciences) for 30 min. Uptake was stopped by washing the cells with ice-cold KRH buffer. Cells were harvested and dissolved in 300 μL lysis buffer (10 mM Tris-HCl, pH 8.0, 0.2% SDS). The radioactivity level of each aliquot was determined by the liquid scintillation spectrometry. Disintegrations per minute value was used to evaluate the intracellular level of [3H]-2-deoxyglucose. The experiment was performed in triplicate.
Statistical analysis
Results are shown as the mean ± SD from three independent replicates. The Student's t-test was used to determine the significance between two groups. Survival curves were obtained from Kaplan–Meier estimates and validated with the log-rank test. p-values <0.05 was considered as statistically significant.
Results
CASC9 is highly expressed in human NPC
To investigate the expression of CASC9 in NPC, we measured the expression abundance of CASC9 in a panel of human NPC cell lines. CASC9 showed a higher expression in NPC cancer cell line CNE-2, SUNE-1, HNE-1, HONE-1, and CNE-1 compared with that of in normal cell line NP-69 (Fig. 1A). To examine the mRNA level of CASC9 in cancer tissues, reverse transcription-quantitative PCR assay was performed with 20 paired NPC tissues and adjacent normal tissues. The results showed that CASC9 mRNA expression level was significantly upregulated in cancer tissues in comparison with that of normal counterparts (Fig. 1B). To elevate the clinical significance of CASC9, we investigated the percentage of NPC patients with highly expressed CASC9 (>2.5-fold). As shown in Figure 1C, significantly increased CASC9 expression was observed in 75% of NPC patients. And more interestingly, Kaplan–Meier analysis showed that higher CASC9 expression was significantly associated with a poor overall survival in NPC patients (Fig. 1D). These results suggested that high level of CASC9 has considerable clinical significance in the NPC diagnosis, and overexpression of CASC9 may be important for NPC progression.
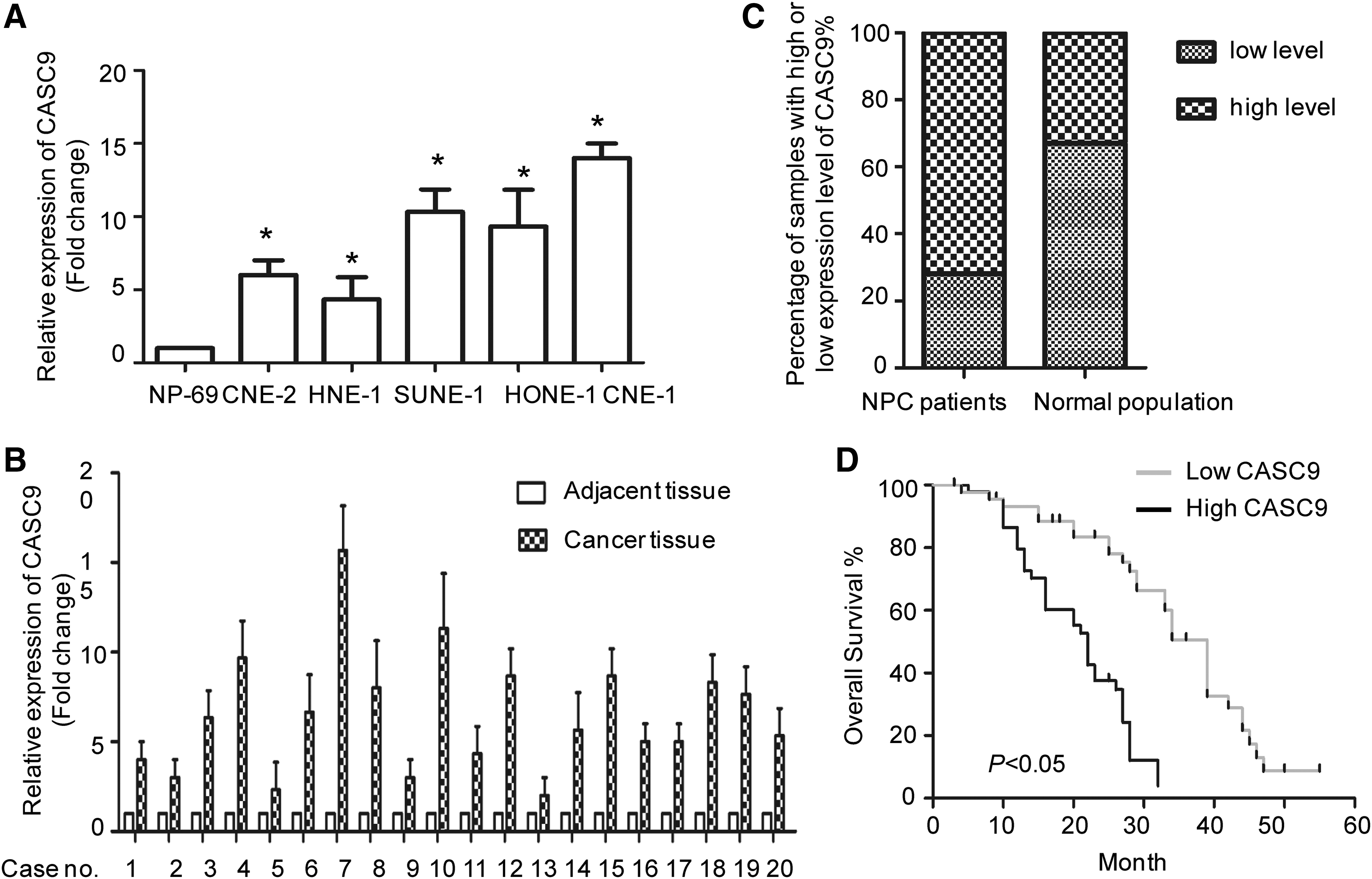
Relative expression of CASC9 in human NPC cell line and tissues.
CASC9 promotes NPC cell growth
As there is high expression of CASC9 in NPC patients, we determined the role of CASC9 in CNE-1 cell growth. MTT assay showed that CNE-1 cells with stably expressed CASC9 had a higher proliferation rate than that of the control cells (Fig. 2A). However, overexpression of CASC9 in normal NP-69 cells has no significant effect on cell growth (Fig. 2B). To further demonstrate the role of CASC9 in NPC cell proliferation, cell cycle analysis was performed and the result demonstrated that overexpression of CASC9 in CNE-1 cells decreased the proportion of cells in the G0/G1 phase (Fig. 2C). Consistently with these results, in vitro soft agar colony formation assay showed that CNE-1 cells harboring higher level of CASC9 generated more and larger colonies compared with that of control cells (Fig. 2D). To determine whether inhibition of CASC9 expression suppresses NPC progression, cell apoptosis assay was carried out with CNE-1 cells harboring depleted CASC9. As shown in Figure 2E, downregulation of CASC9 obviously induced the cell apoptosis rate. These results suggested that high expression of CSAC9 facilitates NPC cell growth.
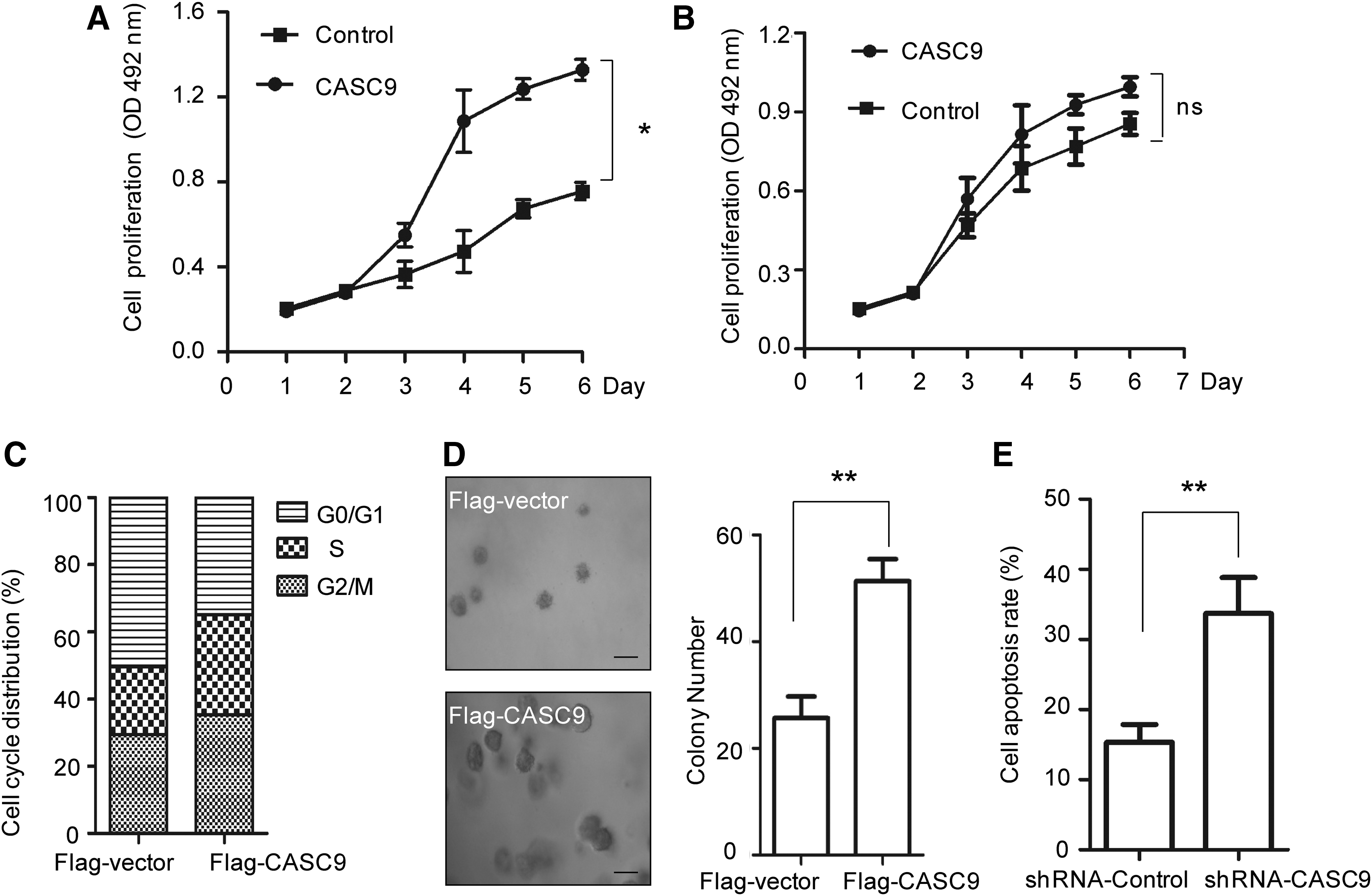
CASC9 promotes NPC cell growth.
CASC9 interacts with HIF1α and enhances its stability
To uncover the molecular mechanism of CASC9 in regulating NPC tumorigenesis, we performed an RNA pull-down assay followed by MS to identify CASC9-binding candidates. Interestingly, CASC9 specifically associated with HIF1α in the NPC cells (Fig. 3A). To confirm this result, CASC9 was conjugated to D-Biotin, and the interaction between CASC9 and HIF1α was examined using recombinant HIF1α by the in vitro streptavidin-binding assay. As shown in Figure 3B, specific association between CASC9 and HIF1α was observed. These results demonstrated that CASC9 binds to HIF1α and raise the important question of whether CASC9 is relevant in the regulation of HIF1α in the fundamental cellular conditions.

CASC9 interacts with HIF1α and increases the protein abundance of HIF1α.
As the binding partner of CASC9, to detect whether CASC9 affects the stability of HIF1α, CNE-1 cells were transfected with CASC9 and the protein level of HIF1α was examined with the indicated antibodies. The result showed that overexpression of CASC9 enhanced the protein abundance of HIF1α (Fig. 3C). qPCR assay was also preformed with CNE-1 cells expressing CASC9. The result indicated that overexpression of CASC9 has no significant effect on the mRNA level of HIF1α (Fig. 3D). Previous studies have found that stabilization of HIF1α triggers the transcription of HIF1α target genes, including GLUT1, EPO, VEGF, and LDHA. To detect the regulation of CASC9 on HIF1α, RT-PCR assay was performed to evaluate the expression level of HIF1α targeting genes. As shown in Figure 3E, significantly increased HIF1α-dependent gene expression was observed with CASC9 overexpression. These results demonstrated that CASC9 is a novel upstream activator of HIF1α.
CASC9 promotes glycolysis metabolism
HIF1α adapted to cellular response to low oxygen condition by activating target gene expression controlling glycolysis metabolism of cancer cells (Depoix et al., 2016; Lin et al., 2016; Slemc and Kunej, 2016). As CASC9 stimulates the activity of HIF1α, the reprogramming of glycolysis by CASC9 was further examined by in vitro glucose uptake assay. The result showed that cells harboring overexpressed CASC9 represented a significantly increased glucose uptake compared with that of control cells (Fig. 4A). Consistently, the lactate production was also dramatically upregulated in CNE-1 cells with highly expressed CASC9 (Fig. 4B). These results indicated that CASC9 activated HIF1α and promoted cancer cell glycolysis metabolism.
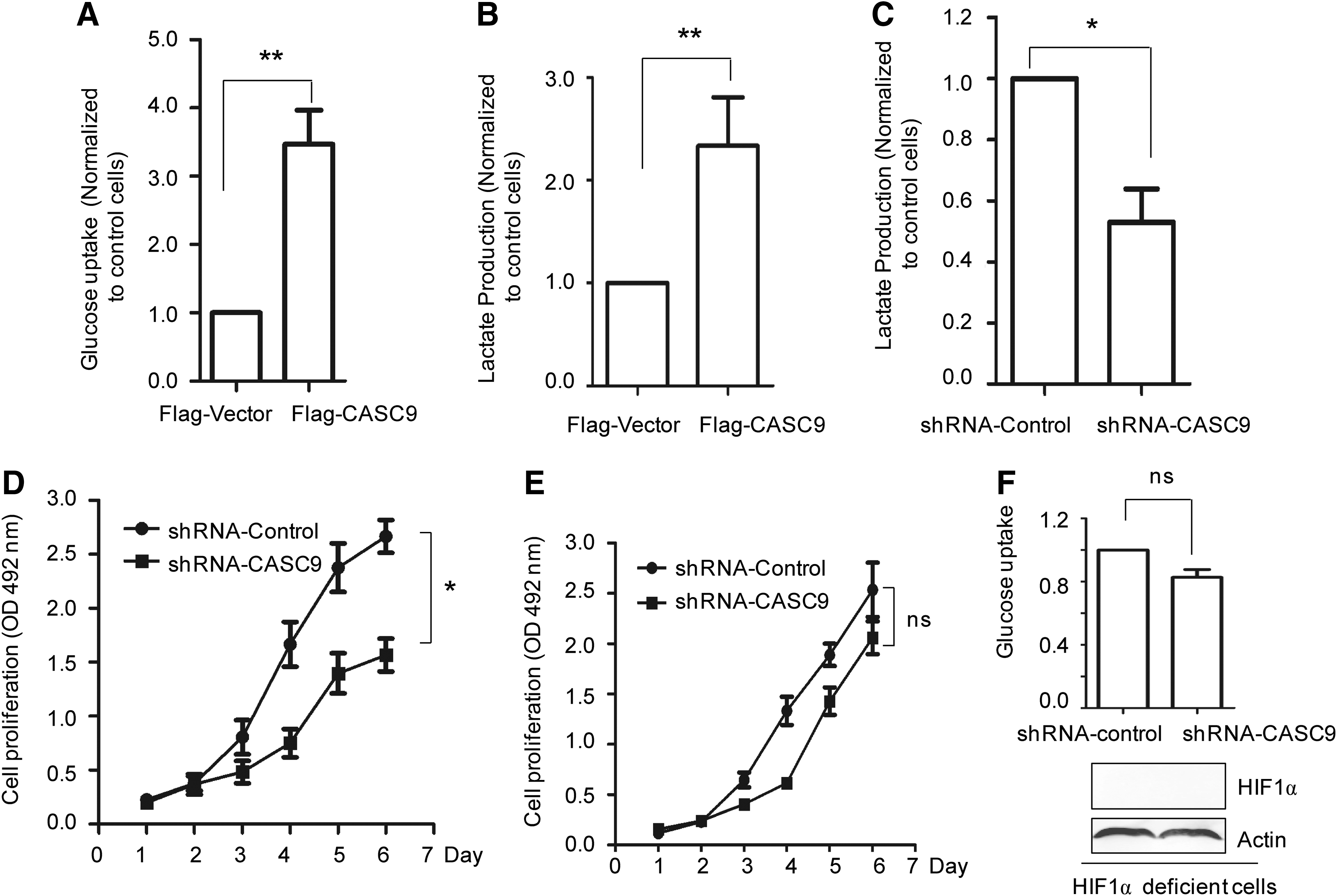
CASC9 enhances the glucose metabolism of NPC cell.
To further demonstrate the essential role of CASC9 in promoting glycolysis and cancer cell growth, the glucose uptake of CNE-1 cells with depleted CASC9 was estimated. As shown in Figure 4C, deficiency of CASC9 obviously decreased the lactate production. MTT assay also suggested that downregulation of CASC9 inhibits the CNE-1 cell proliferation (Fig. 4D). To demonstrate the role of CASC9 in regulating cell growth and glucose metabolism was HIF1α-dependent, CASC9 was depleted in HIF1α deficient CNE-1 cells. The results showed that downregulation of CASC9 has no significant effect on CNE-1 cell growth and glucose uptake in the absence of HIF1α (Fig. 4E, F). The result indicated that the regulation of CACS9 in CNE-1 cell growth and glucose metabolism was tightly associated with HIF1α. Taken together, these results demonstrated that CASC9 activates HIF1α, which leads to the reprogramming of glycolysis metabolism and facilitates cancer cell growth.
Discussion
Increasing evidence has provided insights into the important function of lncRNAs in regulating tumorigenesis. CASC9 is a lncRNA identified by a next-generation sequencing analysis in esophageal cancer, which has been demonstrated overexpressed in ESCC through bioinformatics analyses (Pan et al., 2016), however, the roles and mechanisms of CASC9 in cancers have not been well established. Our present study demonstrated that CASC9 is significantly overexpressed in human NPC tissues and facilitates tumor progression. The oncogenic function of CASC9 was partially attributed to its ability to enhance HIF1α stability, which subsequently activated the target gene transcription of HIF1α and reprogrammed cellular glucose metabolism. Our study revealed the molecular mechanism of CASC9 in regulating NPC carcinogenesis and contributes to an increasing evidence supporting the involvement of lncRNAs in cancer progression.
The high expression level of CASC9 in both ESCC and NPC highlights the significance to determine the underlying mechanism of CASC9 in cancers. Cancer cells utilized higher levels of glucose in the presence of oxygen with an associated increase in lactate production (Nishida et al., 2015). This phenomenon of aerobic glycolysis, termed the Warburg effect, has been observed in a variety of cancers, which provides critical biosynthetic intermediates required for cell growth and division (Pavlova and Thompson, 2016). There was an increase in support that HIF1α activation under tumor hypoxia confers adaptive advantages to cancer cell by inducing the expression of genes encoding glycolytic enzyme and glucose transporters (Iyer et al., 1998; Nakazawa et al., 2016). Therefore, the regulation of HIF1α plays an important role in cancer progression. It is well established that hydroxylation of HIF1α mediated by the proline hydroxylase domain protein induces HIF1α ubiquitination and degradation under normoxic conditions (Lee et al., 2016). Our study identified CASC9 as a novel regulator of HIF1α, which extended the involvement of CASC9 in cytoplasmic signal transduction.
In summary, our study identified CASC9 as an oncogenic regulator in NPC progression through activating HIF1α, which finally enhances the glucose metabolism of cancer cells. The higher expression of CASC9 in NPC patients highlights the clinical significance of CASC9 in cancer prognosis. Our study also identified the strong correlation of CASC9 expression level and NPC therapeutic outcome. The findings indicated the potential application of CASC9 as a useful biomarker in the identification of patients who are at a high risk of NPC progression. Further study is encouraged to explore the upstream regulator that facilitates the overexpression of CASC9 in cancer cells. The remarkably inhibitory effect of CASC9 depletion on the glucose metabolism and tumor growth suggested that downregulation of CASC9 in cancer cell may be a promising target for cancer therapy.
Conclusion
Our study demonstrated that CASC9 regulating NPC progression through functioning as a novel regulator of HIF1α. These findings provide insights into the critical role of CASC9 in cancers and highlight the potential application of CASC9 in NPC diagnosis and treatment.
Footnotes
Acknowledgment
We thank the Cangzhou Central Hospital for providing the clinical samples used in this study.
Disclosure Statement
No competing financial interests exist.