
Abstract
Safety concerns of some local anesthetics, such as lidocaine, have been raised in recent years due to potential neurological impairment. Dexmedetomidine may protect humans from neurotoxicity, and miR-let-7b is activated by nerve injury; however, the roles of miR-let-7b and its target gene in lidocaine-induced cytotoxicity are not well known. Through bioinformatics and a luciferase reporter assay, COL3A1 was suggested as a direct target gene of miR-let-7b. Here, we confirmed by measuring mRNA and protein levels that miR-let-7b was downregulated and COL3A1 was upregulated in lidocaine-treated cells, an observation that was reversed by dexmedetomidine. Similar to miR-let-7b mimics or knockdown of COL3A1, dexmedetomidine treatment reduced the expression of COL3A1, suppressed cell apoptosis and cell migration/invasion ability, and induced cell cycle progression and cell proliferation in PC12 cells, effects that were reversed by the miR-let-7b inhibitor. Meanwhile, proteins involved in cell apoptosis, such as Bcl2 and caspase 3, were impacted as well. Taken together, dexmedetomidine may protect PC12 cells from lidocaine-induced cytotoxicity through miR-let-7b and COL3A1, while also increasing Bcl2 and inhibiting caspase 3. Therefore, miR-let-7b and COL3A1 might play critical roles in neuronal injury, and they are potential therapeutic targets.
Introduction
L
Dexmedetomidine, a novel sedative-analgesic agent, is considered a highly specific and selective α2-adrenoceptor agonist that provides sedation, analgesia, and sympatholysis, and it is widely used as an anesthetic in small animals and humans to provide sedation, anxiolysis, and analgesia (Szumita et al., 2007; Tang et al., 2015). By meta-analysis, long-term intrathecal dexmedetomidine application was found to induce permanent neuronal damage, especially after spinal anesthesia (Tan et al., 2015). Scientists have also rationalized combining such drugs for the improvement of postoperative or neuropathic pain treatment in a rat model of neuropathic pain for reasons including synergistic interaction and side-effects of intrathecal morphine and dexmedetomidine (Kabalak et al., 2013). Scientists have hypothesized that total intravenous anesthesia, consisting of lidocaine combined with dexmedetomidine as an opioid substitute, is a feasible technique due to the advantages of intravenous lidocaine infusion in the perioperative period (Mizrak et al., 2012; Hwang et al., 2015); however, limited data exist regarding the molecular mechanism of dexmedetomidine's protective role, warranting further exploration.
Mature microRNAs (miRNAs) function by specifically binding the 3′ UTR of mRNA to interrupt mRNA translation, cause downregulation of protein expression through mRNA cleavage, or facilitate translational repression (Deng et al., 2016; Wang et al., 2016). miRNAs have been implicated in neurogenesis, neuronal degeneration, and memory function, and they are highly enriched in the brain. Initially identified in Caenorhabditis elegans, the lethal-7 (let-7) gene is a founding member of let-7 miRNAs and is highly conserved across species. Attention has been increasingly paid to the involvement of let-7 miRNAs in carcinogenesis and nerve damage, as let-7 miRNAs regulate neuronal cell fate and affect both neurodegeneration and neuronal regeneration (Lehmann et al., 2012; Boese et al., 2016). However, it is unknown whether let-7 miRNAs mediate neuroprotection under dexmedetomidine treatment conditions after lidocaine-induced injury.
In this study, we assessed the molecular mechanism of lidocaine-induced cytotoxicity and the role of miR-let-7b in differentiated PC12 cells. We found that miR-let-7b modulates the cellular expression of COL3A1 and contributes to cell proliferation, apoptosis, cycle, migration, and invasion in response to lidocaine.
Materials and Methods
Cell culture and drug treatment
A rat pheochromocytoma cell line (PC12) was supplied from American Type Culture Collection (Manassas, VA) and maintained in RPMI 1640 (Life Technologies, Grand Island, NY) with 10% heat-inactivated fetal bovine serum (GIBCO/BRL) at 37°C in a humidified atmosphere of 5% CO2. To mimic the effects of cytotoxicity, 1 mM lidocaine was added to the medium for 24 h after diluting into single-cell suspensions and seeding in 12-well plates (1 × 105 cells/mL). Then, PC12 cells were transfected with or without dexmedetomidine (10.0 μM), miR-let-7b mimics/NC (1 nM), miR-let-7b inhibitor (1 nM), and COL3A1-siRNA/NC (1 nM) (RIB Bio, Guangzhou, China) for another 48 h, by using Lipofectamine 2000 (Invitrogen). Subsequently, the RNA and protein were harvested at the specified times. Cells treated only with lidocaine were used as the control group.
Bioinformatics methods
The potential targets of miR-let-7b were obtained by using computer-aided algorithms from TargetScan and data from the literature. The online programs PicTar (
Recombinant plasmid construction and dual-luciferase reporter
The 3′UTR sequence of COL3A1 (NCBI reference sequence NM_000090) was retrieved from the GenBank Database. A fragment encoding the COL3A1-3′UTR sequence was PCR amplified from PC12 genomic DNA by using the forward primer 5′-CCGCTCGAGTTGCTGTTTCACCCACC-3′ and the reverse primer 5′-ATTTGCGGCCGCTCCTTATGATCCCTGCTCT-3′. The PCR product was then cloned into the NotI/Xhol sites of the psiCHECK2 vector (System Biosciences, Mountain View, CA). Mutations of the miR-let-7b binding sites in the COL3A1 3′UTR sequence were performed by using the KOD-Plus-Mutagenesis Kit (Agilent Technologies, Santa Clara, CA) according to the manufacturer's protocol. All plasmids were confirmed by restriction enzyme digestion and sequencing.
PC12 cells were then transfected with luciferase reporter vector and miRNA by using Lipofectamine 2000. Firefly and Renilla luciferase activities were measured by using the Dual-Luciferase Reporter Assay System (Promega). Results were expressed as the firefly luciferase activity normalized to Renilla luciferase activity.
RNA isolation and real-time PCR
Total RNA from PC12 cells was extracted by using TRIZOL (Invitrogen) according to the manufacturer's instructions and quantified by spectrophotometry. To measure the mRNA expression levels of miR-let-7b and COL3A1, the following gene-specific primers were used: COL3A1 Forward: 5′-CTTCCGACACTCTTCCTTCG-3′ Reverse: 5′-ATAGCACCATCAGGGGTGAG-3′; GAPDH (as the appropriate endogenous controls) Forward: 5′-GGCTACACTGAGGACCAGGTT-3′ Reverse: 5′-TGCTGTAGCCGTATTCATTGTC-3′; miR-let-7b Forward: 5′-UGAGGUAGUAGGUUGUGUGGUU-3′ Reverse: 5′-CTCAACTGGTGTCGTGGA-3′; and U6 Forward: 5′-CTCGCTTCGGCAGCACA-3′ Reverse: 5′-AACGCTTCACGAATTTGCGT-3′.
Mature miR-let-7b expression was measured by quantitative real time-PCR using the TaqMan® MicroRNA Reverse Transcription Kit and the TaqMan MicroRNA Assays (Applied Biosystems, Foster City, CA) and normalized to U6 according to the manufacturer's instructions.
Protein isolation and Western blotting analysis
PC12 cells (2 × l06) were lysed, separated by gel electrophoresis, and analyzed by traditional Western blotting procedures. Antibodies incubated were anti-rat polyclonal COL3A1 antibody (1:20,000; Sigma-Aldrich), anti-rat polyclonal Bcl2 antibody (1:1000; Abcam), anti-rat polyclonal caspase 3 antibody (1:10,000; Abcam), and anti-rat polyclonal GAPDH antibody (1:1000; Santa Cruz Biotechnology). Blots were visualized by using a chemiluminescence reagent (Millipore, Temecula, CA) in a LAS4000 Luminescent Image Analyzer (GE Healthcare Japan, Japan).
Colony formation assay
Cell proliferation assays were performed according to a published procedure. Forty-eight hours after transfection, a sample of 1 × 104 PC12 cells was harvested and added to 2 mL of RPMI 1640 containing 10% fetal bovine serum, 0.3% agarose (Difco, Detroit, ML), which was overlaid onto 0.6% bottom agar (Agarose type VII; Sigma-Aldrich) and an appropriate growth factor. The suspension was allowed to harden over a 2-mL bed of previously hardened 0.3% agarose in culture medium in a 60-mm dish. The samples were incubated for the next 7 days, and the number of colonies containing >20 cells was determined. Cells were fed every 4–5 days. After 11 days, cells were stained with crystal violet (Invitrogen) overnight.
5-Ethynyl-2′-deoxyuridine assay
The proliferation of treated PC12 cells was also tested by 5-ethynyl-2′-deoxyuridine (EdU) assay. PC12 cells (1 × 105/well) were planted into six-well plates and allowed to adhere for 24 h. Using the EdU DNA Proliferation in Detection Kit (RiboBio, Guangzhou, China) according to the manufacturer's instructions, PC12 cells were incubated with EdU for 4 h before fluorescence detection. Cells were fixed with 2% paraformaldehyde for 15 min and stained with the EdU kit according to the manufacturer's instructions. Finally, cells were analyzed with an FACScalibur (BD Biosciences, San Jose, CA) flow cytometer.
Hoechst 33258 staining assay
PC12 cells at the logarithmic-growth phase were seeded into 96-well plates (1 × 104/well). To detect morphological evidence of apoptosis, cell nuclei were visualized by DNA staining with the fluorescent dye Hoechst 33258 (Beyotime, China). The cells were cultured in normal medium (control group) or with increasing concentrations of rosiglitazone (5 and 25 μM) for 24 h. Then, PC12 cells were stained with Hoechst 33258 (1 μg/mL) for 15 min in the dark. Results were tested by visual observation of nuclear morphology through the 80i fluorescence microscope (Nikon, Tokyo, Japan) that was equipped with a UV filter (Nikon).
Flow cytometer assay
To determine the fraction of apoptotic cells after transfection, cells (1 × 103/well) were re-suspended in the culture medium and analyzed by Sulphorhodamine Apoptosis Detection Kit Caspase Assay (Immunochemistry Technologies LLC, Bloomington, MN) according to the manufacturer's instructions to determine the fluorescent inhibition of caspase activity. To determine the cell cycle change after transfection, cells (1 × 103/well) were re-suspended in 250 μL binding buffer and then fixed at 4°C overnight. After treatment, the cells were collected again and incubated with 200 μL propidium iodide (20 μg/mL) by using a cell cycle staining kit (Multiscience, Hangzhou, China).
The samples were immediately analyzed by using FACScalibur (BD Biosciences). Data acquisition and analysis were performed by using CellQuest software.
Cell migration/invasion ability analysis
In brief, the lower compartments were filled with RPMI1640 that was pre-coated with 10 mg/mL collagen for 1 h at 37°C. The cell suspension (0.5 mL) was placed on the upper chambers of transwell inserts (8-μm pore size; Corning) for 4 h at 37°C before the migration assay was conducted. The upper chambers were coated with Matrigel (354234; BD Bioscience) before inoculation of the PC12 cells with dexmedetomidine, miR-let-7b, or COL3A1-siRNA in the invasion assay. The cells that migrated to the lower layer and attached to the membrane were stained with crystal violet and were counted in five fields per well under a microscope.
Data analysis
All experiments were run in triplicate and carried out three independent times. Quantitative data were expressed as the mean ± standard deviation and were evaluated by using two-tailed t tests. Data were considered significant at a value of p < 0.05. Statistical analysis was performed by using the SPSS/Win13.0 software (SPSS, Inc., Chicago, IL).
Results
Effect of miR-let-7b on COL3A1 expression in lidocaine-treated PC12 cells
To detect changes in endogenous miR-let-7b and COL3A1, we conducted real-time PCR to investigate miR-let-7b levels and COL3A1 mRNA expression, with or without dexmedetomidine treatment after pretreatment with lidocaine for 24 h. As shown in Figure 1, miR-let-7b (Fig. 1A) was downregulated, and COL3A1 mRNA expression (Fig. 1B) was increased in cells treated with lidocaine only. Levels of miR-let-7b (Fig. 1C) were increased, and COL3A1 mRNA expression (Fig. 1D) was decreased in cells treated with both lidocaine and dexmedetomidine, compared with the control group.
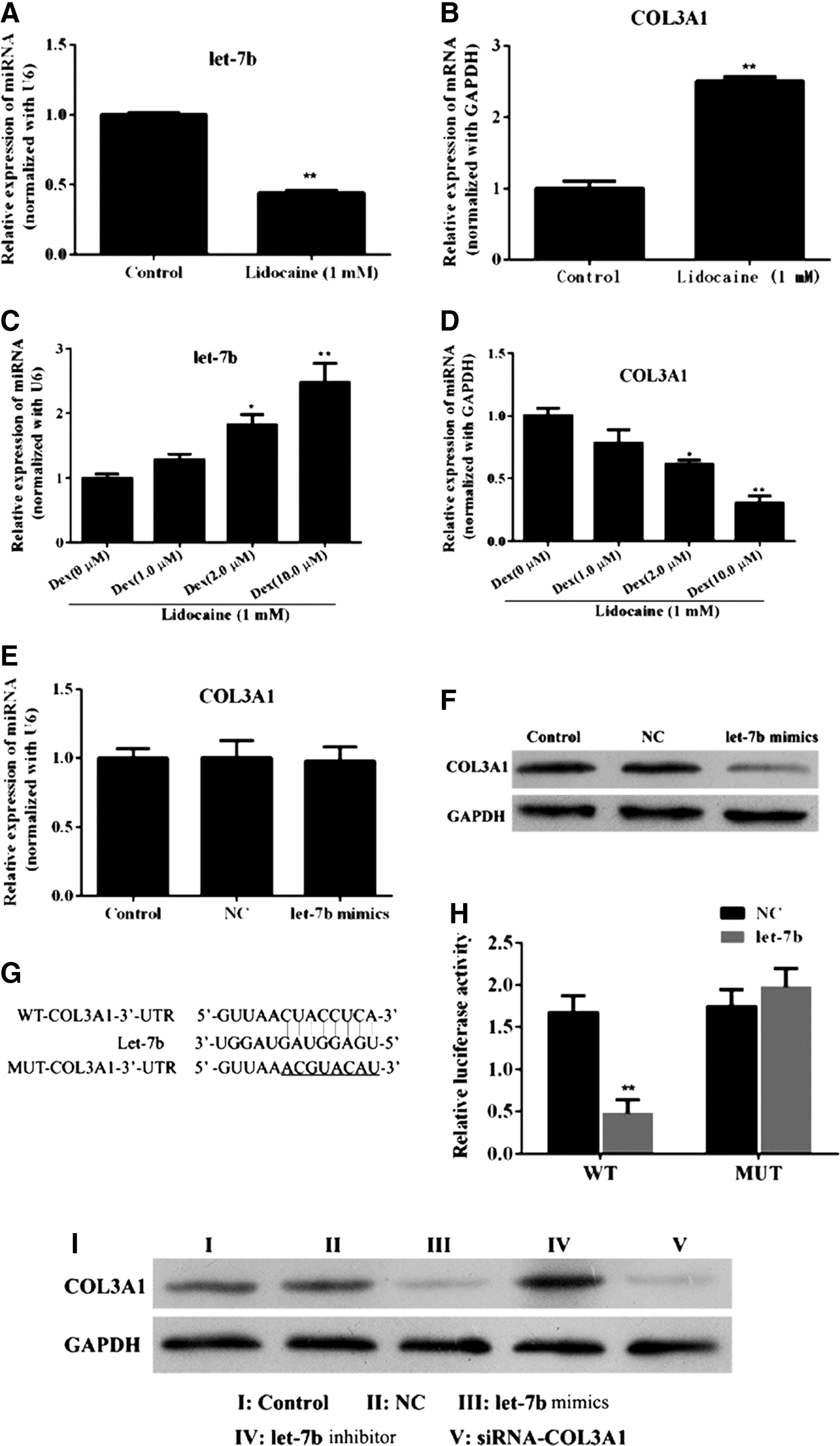
Effect of miR-let-7b on COL3A1 in PC12 cells.
Based on recent reports from the literature, we chose COL3A1 as the candidate miR-let-7b target gene according to predicted bioinformatic results. Before prediction, mRNA and protein levels of COL3A1 were detected by using real-time PCR and Western blotting with overexpressed miR-let-7b. As shown in Figure 1E and F, miR-let-7b had no effects on the mRNA expression of COL3A1 in transcription level (Fig. 1E) but inhibited the protein expression of COL3A1 in translation level (Fig. 1F) in PC12 cells after transfection with miR-let-7b mimics, compared with the negative control (NC) group at the same time point. To explore the interaction between miR-let-7b and COL3A1, cell lysates were detected by using a dual-luciferase assay system for the potential binding region (Fig. 1G). With pretreatment using miR-let-7b mimics, we found that the psiCHECK2-COL3A1-3′UTR-WT group decreased the luciferase activity in PC12 cells (Fig. 1H) compared with the psiCHECK2-COL3A1-3′UTR-MUT group.
To further confirm the exogenous change of COL3A1, PC12 cells were transfected with miR-let-7b mimics/NC/inhibitor after treatment with lidocaine and with or without dexmedetomidine, and protein was detected by Western blot analysis. As shown in Figure 1I, COL3A1 protein levels were significantly decreased in the miR-let-7b mimic group, COL3A1-siRNA group, and lidocaine/dexmedetomidine combination group, compared with the lidocaine-only group and normalized to the miR-NC group. Among the miR-let-7b mimic, COL3A1-siRNA, and lidocaine/dexmedetomidine combination groups, there was no statistically significant difference in the amount of COL3A1 protein detected.
Effect of overexpressed miR-let-7b on PC12 cell proliferation
We examined the effects of miR-let-7b on cell proliferation, as shown in Figure 2. First, we counted the number of colonies containing >20 cells in eight randomly chosen fields for each cell culture condition. Since colony formations have transforming activity, the anchorage-independent multiplication of PC12 cells embedded in soft agar (Fig. 2A) was induced in the miR-let-7b mimic group, COL3A1-siRNA group, and lidocaine/dexmedetomidine combination group, compared with the lidocaine only group and normalized to the miR-NC group. Among the miR-let-7b mimic, COL3A1-siRNA, and lidocaine/dexmedetomidine combination groups, there was no statistically significant difference in multiplication.
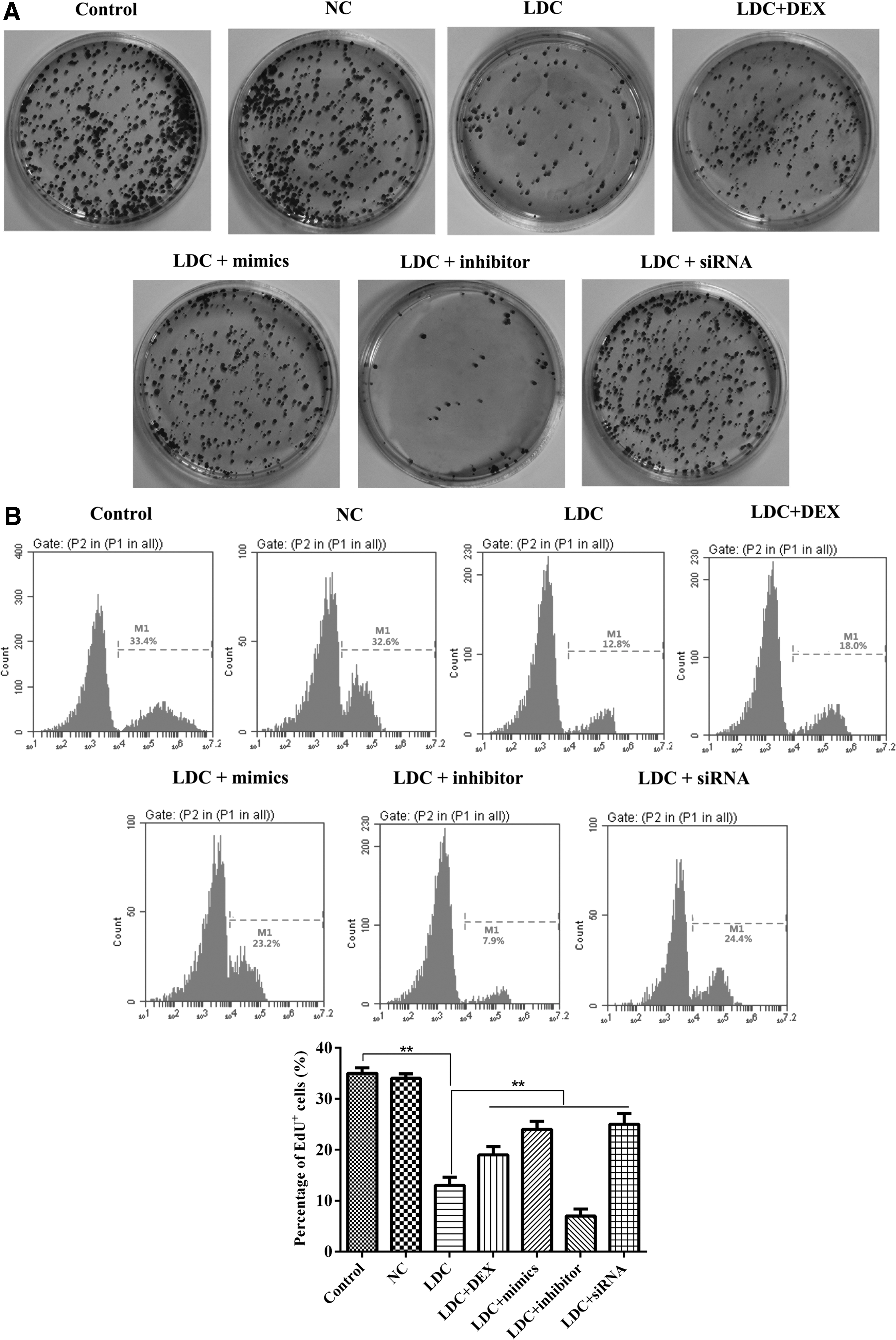
Effect of miR-let-7b on PC12 cell proliferation.
Next, we used another assay to detect cell proliferation that suggested a result similar to that of the colony formation assay. PC12 cells were transfected with miR-let-7b (for over-expression) or COL3A1-siRNA (for knockdown) followed by EdU assay. As illustrated in Figure 2B, the ratio of EdU-positive cells to total cells was promoted in the miR-let-7b mimic group, COL3A1-siRNA group, and lidocaine/dexmedetomidine combination group.
Effect of overexpressed miR-let-7b on PC12 cell apoptosis
The induction of apoptosis was employed to evaluate the protective effect of dexmedetomidine and miR-let-7b. Morphologic changes were first examined by Hoechst 33258 staining (Fig. 3A). After PC12, cells were treated with miR-let-7b mimics/NC/inhibitor, followed by treatment with dexmedetomidine and lidocaine; more apparent apoptosis-induced morphologic changes were observed as compared with the NC. In the control group, the nuclei of PC12 cells exhibited evident characteristics of apoptosis, including cell shrinkage, deformation or loss of membrane integrity, nuclear fragmentation, and chromatin compaction with late apoptotic appearance; whereas PC12 cells were round and homogeneously stained, with fewer cells exhibiting characteristics of apoptosis.
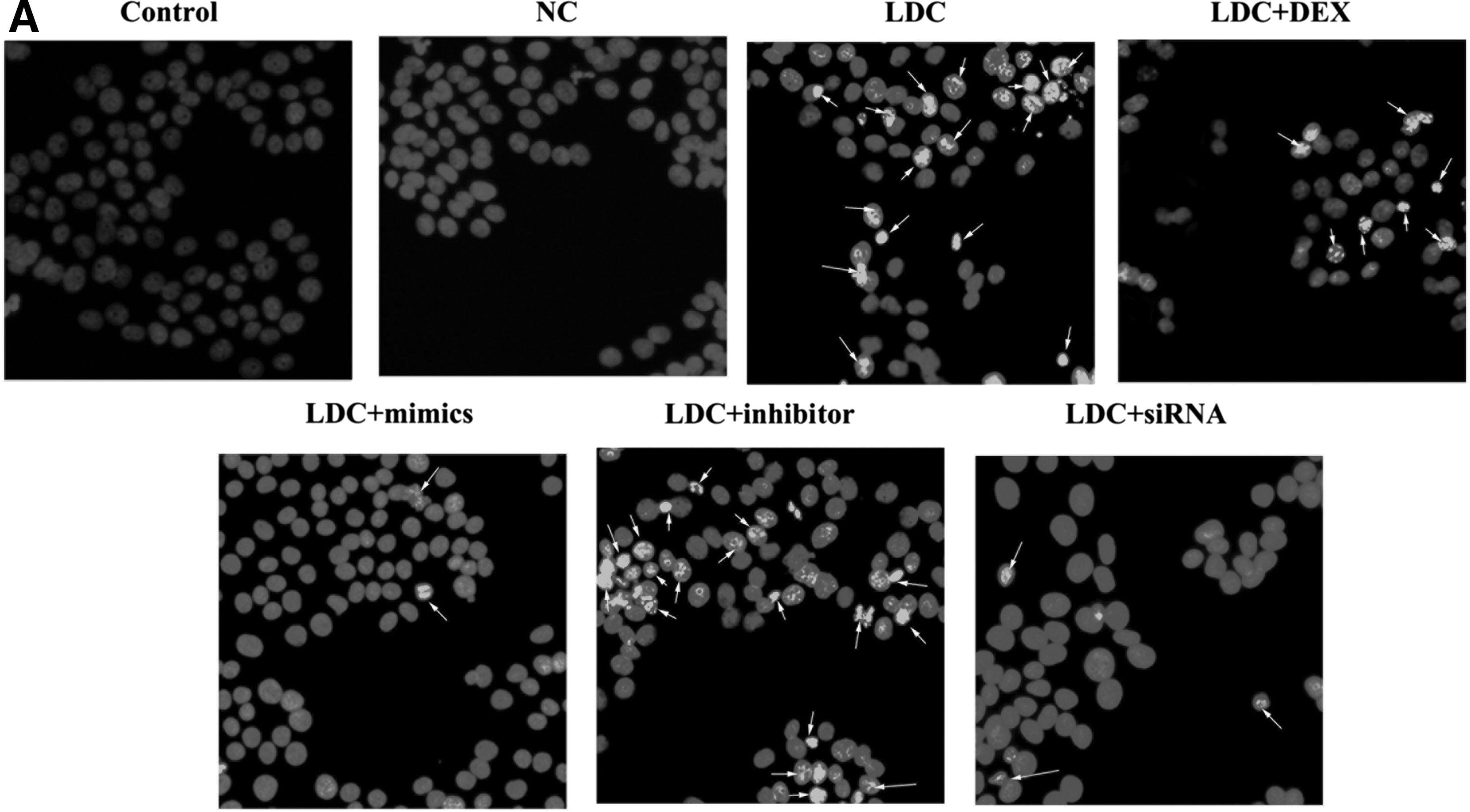
Morphologic changes and apoptosis of PC12 cells in vitro.

The apoptosis inhibitory activity of miR-let-7b and dexmedetomidine in PC12 cells was then analyzed by flow cytometry assay. Our data showed a significant decrease in the quantity of apoptotic cells after exposure to miR-let-7b or dexmedetomidine for 48 h (Fig. 3B). In the miR-let-7b mimic group, COL3A1-siRNA group, and lidocaine/dexmedetomidine combination group, there were only a few apoptotic cells, whereas the percentage of apoptotic cells was higher in the miR-NC group and lidocaine-only group.
To explore the effect of miR-let-7b on several cell cycle relevant proteins, we harvested PC12 cells after treatment with miR-let-7b mimics for 48 h, and then detected the expression of Bcl2 and caspase 3 protein (Fig. 3C). The expression of Bcl2 was increased and that of caspase 3 was significantly downregulated in PC12 cells in the miR-let-7b mimic group, COL3A1-siRNA group, and lidocaine/dexmedetomidine combination group, compared with the lidocaine-only group and normalized to the miR-NC group, which was reversed after miR-let-7b inhibitor transfection.
Effect of overexpressed miR-let-7b on the PC12 cell cycle
Cell cycle distribution was analyzed by using flow cytometry after transfection with miR-let-7b mimics/NC/inhibitor for 48 h. Compared with the NC group, the ratio of G2/G1 was reduced and S cell cycle arrest was decreased in transfected cells (Fig. 4A, B).
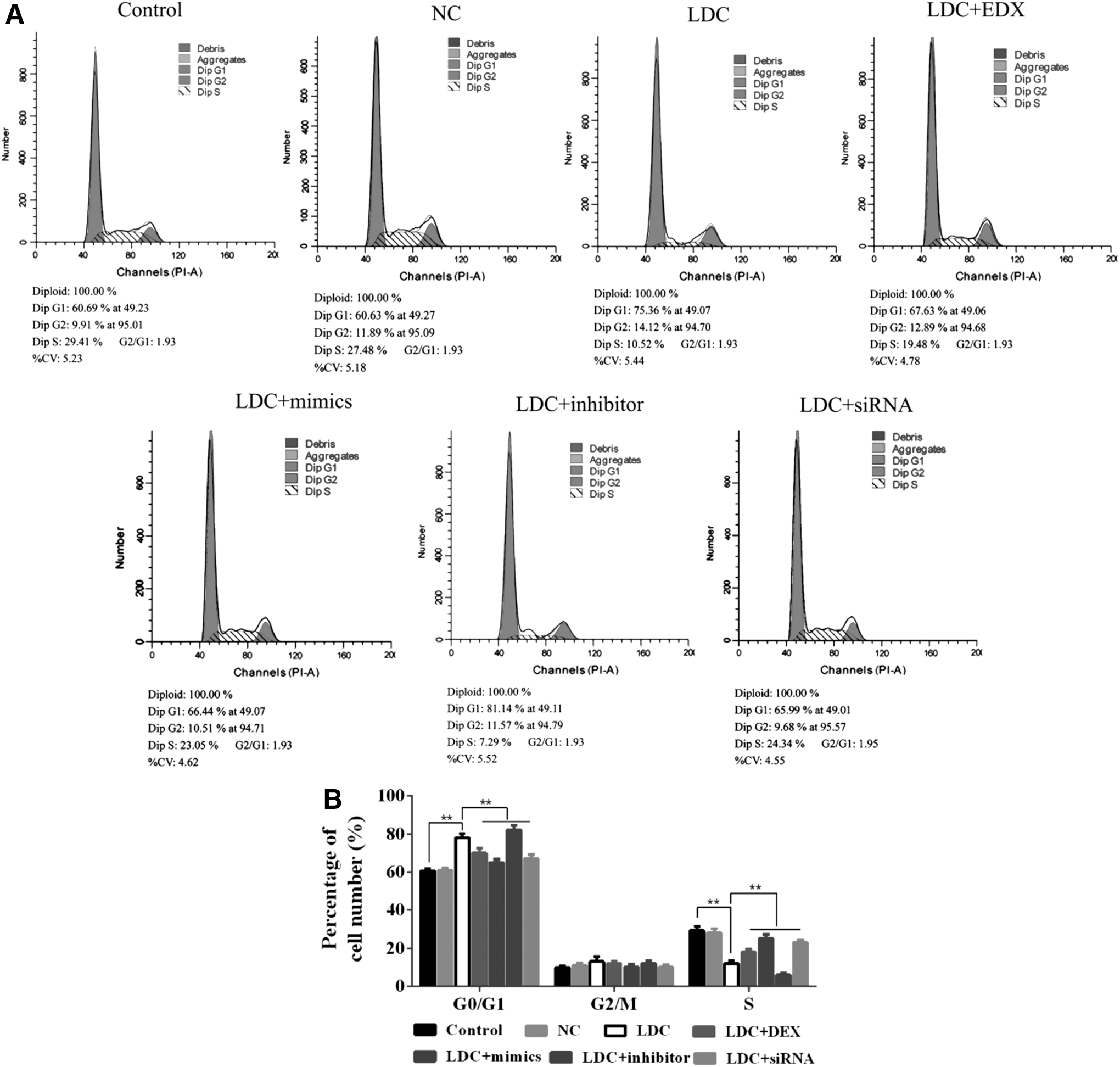
Effect of miR-let-7b on PC12 cell cycle change.
Effect of overexpressed miR-let-7b on PC12 cell migration/invasion ability
As shown in Figure 5A, the invasion number of PC12 cells in the miR-let-7b mimic group, COL3A1-siRNA group, and lidocaine/dexmedetomidine combination group was significantly higher than that in the lidocaine-only group, but lower (although, not significantly) than that of the control groups. In addition, the migration potential of PC12 cells in the miR-let-7b mimic group, COL3A1-siRNA group, and lidocaine/dexmedetomidine combination group was significantly higher than that of the lidocaine-only group, but lower (although, not significantly) than that of the control groups (Fig. 5B).
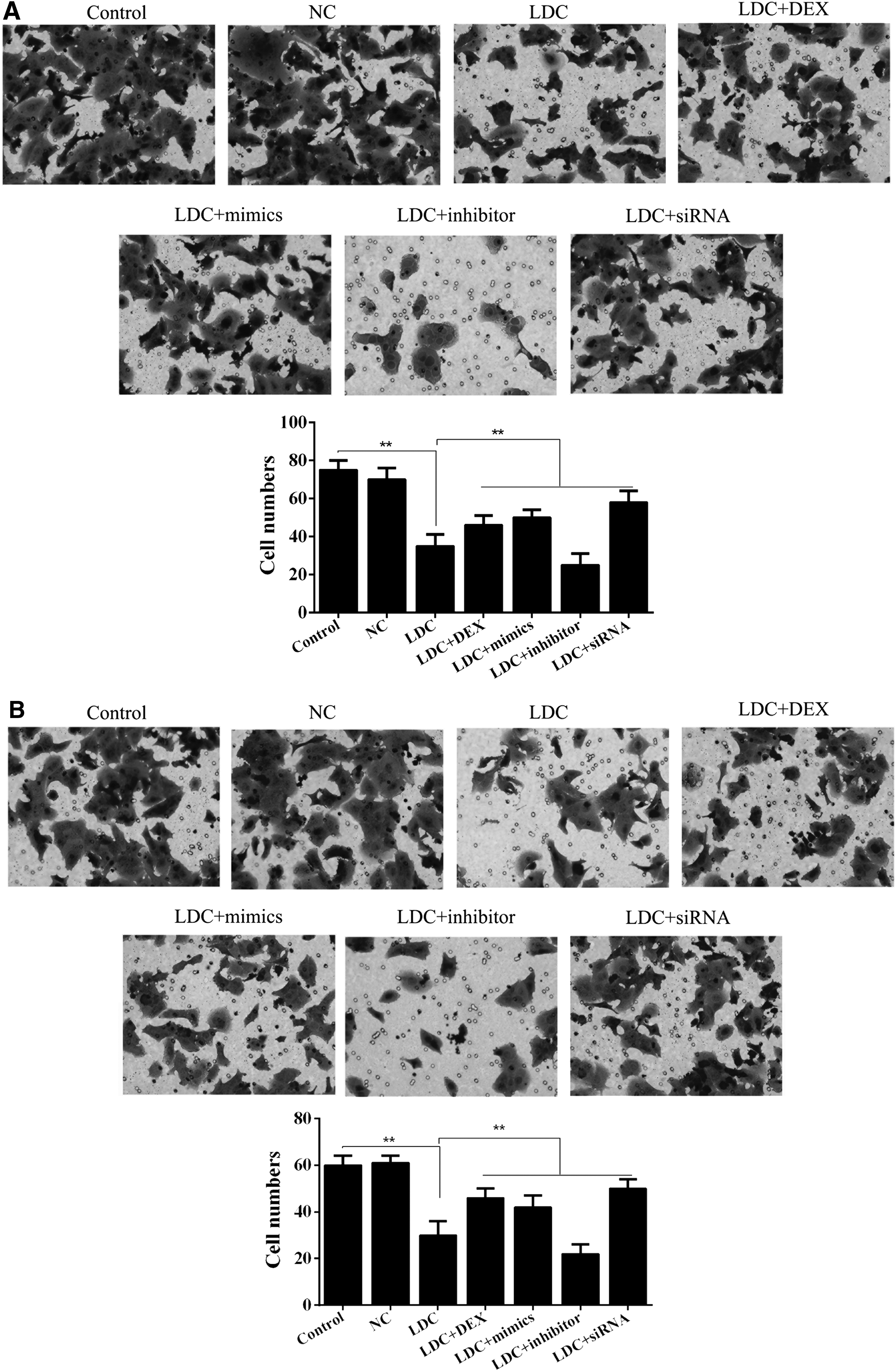
Effect of miR-let-7b on PC12 cell migration and invasion ability.
Discussion
In recent years, the regional use of local anesthetics such as lidocaine has raised concerns due to the side effects of neurotoxicity and other tissue damage. The combined use of analgesics and anesthetics has been applied in the clinic. Dexmedetomidine has been considered a good candidate for decreasing the damage caused by local anesthetics, but prior research in this area is limited. Our study revealed the cytoprotective effect of miR-let-7b- and COL3A1-mediated α2-adrenergic receptor activation during lidocaine exposure in PC12 cells. We found that miR-let-7b was downregulated in lidocaine-treated PC12 cells with morphological infarction, and that overexpressed miR-let-7b could suppress the expression of COL3A1 in PC12 cells.
miRNAs have been shown to be important gene regulators in many organs and tissues, and they have been implicated in a growing number of diseases. The aberrant expression of miRNAs in different damage stages suggests that they may have a critical role in coordinating the regulation of transcriptional and post-transcriptional phases of target mRNA. It has been demonstrated that the expression of miR-let-7b is altered in various tissues and cell types after nerve injury, and our results are consistent with this observation. In malignant neurofibromatosis type 1-associated nerve sheath tumors, miR-let-7b expression is downregulated during mesenchymal transition (Masliah-Planchon et al., 2013). Inhibition of let-7 miRNAs regenerates peripheral nerves by increasing the secretion of nerve growth factor by primary cultured Schwann cells and enhances axonal outgrowth from a co-culture of primary Schwann cells and dorsal root ganglion neurons (Li et al., 2015). Exogenous miR-let-7b was reported to protect human mesenchymal stem cells implanted into the infarcted myocardium from apoptosis and autophagy by directly targeting caspase-3 (Ham et al., 2015). However, miR-let-7b is induced in cerebrospinal fluid from individuals with Alzheimer's disease as a signaling molecule to identify Toll-like receptor 7, which contributes to the spread of central nervous system damage (Mun et al., 2013; Boese et al., 2016). In our study, we provided evidence that miR-let-7b targets the 3′UTR of COL3A1, which plays an important role in lidocaine-induced toxicity. We further detected the effect of miR-let-7b on PC12 cell apoptosis, proliferation, and cell cycle progression.
To explore the neuroprotective effects of dexmedetomidine, we established a cellular injury model of lidocaine preconditioning. We found that overexpressed miR-let-7b or its silencing target COL3A1 may have a protective effect, as evidenced by significantly enhanced cell survival, decreased cell apoptosis, and improved cell migration/invasion. We confirmed that miR-let-7b inhibits the translation of COL3A1 through targeting the 3′UTR of COL3A1, located on chromosome 2q31, which produces the pro-α1 chains of type III collagen, a fibrillar collagen found in extensible connective tissue (Chan et al., 2010; Loeser et al., 2013). COL3A1 has an important role in adjusting the strength, flexibility of tissues, and modulating the wound response process. Its expression is acutely upregulated in the early phases of wound healing and maintains high levels of expression for several weeks after injury (Dyment et al., 2013; Jiang et al., 2015). Interestingly, in lung cancer, COL3A1 has been reported as the target of the miR-29 family, with increased invasiveness and subsequent downregulation of the miRNA (Chen et al., 2015; Zanotti et al., 2015; Zhu et al., 2015). Another study revealed that COL3A1 may be a direct target of let-7d, with an inverse correlation between RCC growth and metastasis (Su et al., 2014). Our research also demonstrated that COL3A1 may play an important role during the protective effect of dexmedetomidine on lidocaine-induced cytotoxicity.
Taken together, our data reveal that miR-let-7b is a vital regulator in suppressing COL3A1 expression to inhibit apoptosis through activating Bcl2 and inhibiting Caspase 3. These results provide support for the contribution of miR-let-7b in the defense of impaired local anesthetics, which may provide strong theoretical support for future clinical treatment.
Footnotes
Acknowledgment
This work was supported by Guangzhou Women and Children's Medical Center and the Science and Technology Planning Project of Guangdong Province, China (No. 2013B022000024).
Disclosure Statement
No competing financial interests exist.