
Abstract
Recently, more and more evidences unveiled that ubiquitin-proteasome system (UPS) makes an important contribution to the occurrence and development of cancer. HERC4 is one identified Ubiqutin ligase E3, a member of UPS. Although some studies showed that HERC4 abnormally expresses in many cancer cells, till now, nothing has been reported about the function of HERC4 in the development of hepatoma carcinoma. To this end, in this study, we studied the function of HERC4 for the first time in hepatoma carcinoma cells. We detected the expression of HERC4 in tumor and normal tissues, and in hepatoma carcinoma cell lines by using qRT-PCR, Western blot, immunohistochemistry, and immunofluorescence. The data showed that tumor tissues expressed higher HERC4 than normal ones. HERC4 was expressed, although to a different extent, in hepatoma carcinoma cell lines. Colony formation assay, CCK-8 assay, EdU assay, wound healing assay, and FACS indicated that HERC4 plays a role in cell proliferative and migration ability. HERC4 overexpression increases the proliferative and migration ability and reduces apoptosis of hepatoma carcinoma cells; in contrast, knockdown of HERC4 decreases the proliferative and migration ability and increases the apoptosis rate of hepatoma carcinoma cells. Taken together, our findings showed that HERC4 has an effect on the occurrence and development of hepatoma carcinoma by promoting hepatoma carcinoma cell proliferation and migration, and by reducing cell apoptosis, further providing another therapeutic target for the intervention of related diseases.
Introduction
H
Ubiquitin-proteasome system (UPS) contributes to almost every cell live process, proliferation, differentiation, apoptosis, and homeostasis. Ubiquitination of the target protein is achieved through an enzymatic cascade involving the concerted action of E1 (ubiquitin activating), E2 (ubiquitin conjugating), and E3 (ubiquitin ligase) enzymes (Schlossarek et al., 2014). E3 ubiquitin ligase is the most various type and the most important component during the ubiquitination process. HERC family, belonging to ubiquitin ligase E3 and locating to endosome (Hochrainer et al., 2008), contains six members (e.g.,
The role of UPS in tumorigenesis has become obvious. HERC4 is one of ubiquitin ligase family members. Previous works have shown that HERC4 contributes to the occurrence and development of cancer. HERC4 plays an important role in myeloma (Zhang et al., 2016). In addition, some investigators verified that HERC4 highly expresses in lung cancer and breast cancer (Zhou et al., 2013; Zeng et al., 2015). However, the biological functions of HERC4 in cancers are not clear, especially in HCC. In this article, we took HERC4 as the investigative candidate protein and tried to figure out the pathological function of HERC4 in HCC.
Materials and Methods
Antibodies and other reagents
HERC4 and GAPDH monoclonal antibodies were purchased from Abcam company. All the cell lines were purchased from ATCC. All the secondary antibodies, Lipofectamine 2000 kit, and Crystal violet were from Thermo fisher. SYBR Green qPCR Master Mix was obtained from DBI Bestar®. CCK-8 kit was purchased from Dojindo Japan. EdU Kit was obtained from RiboBio, Guangzhou, China. FITC annexin V apoptosis detection kit I was purchased from BD.
Patient selection
Human HCC samples (n = 15) and adjacent liver tissue were freshly obtained from the patients undergoing surgical resection of primary HCC at the Second Affiliated Hospital of Zhejiang Chinese Medical University. Diagnostic confirmation of HCC was defined as having positive histology, cytology, and laboratory test/marker study, direct visualization, or positive radiology and imaging. Patients diagnosed with stomach, colon, lung, pancreatic, breast, or rectal cancers within the 5 years before the date of HCC diagnosis were excluded to further ensure the inclusion of only HCC, rather than metastatic liver cancers. The institutional review board at the Second Affiliated Hospital of Zhejiang Chinese Medical University approved the study protocol, and all patients provided written informed consent.
qRT-PCR analysis
Total RNA was extracted from cells and tissues using Trizol (Invitrogen) following the manufacturer's instructions. The RNAs were reverse transcribed using the qPCR RT Kit in a 20 μL reaction. Primers were designed on exon junctions to prevent coamplification of genomic cDNA, with β-actin as reference. The sequences were as follows: β-actin Forward (5′-3′): ATCGTGCGTGACATTAAGGAGAAG; β-actin Reverse (5′-3′): AGGAAGGAAGGCTGGAAGAGTG and HERC4 Forward (5′-3′): CCTATAATGGGCAGTGTCTACCA; HERC4 Reverse (5′-3′): CACAGTTCTGGGGACTAGAGT.
Cell culture and reagents
HepG2, Bel-7402, Hep3b, SMMC7721, and LM3 cells were cultured in DMEM, supplemented with 10% fetal bovine serum (FBS) and 1% antibiotics in 5% CO2 at 37°C. The siRNA targeting HERC4 and eukaryotic expression vector pcDNA3.0-HERC4 were purchased from Genepharma Company (Shanghai, China). Both plasmids and siRNAs were transfected with Lipofectamine 2000 according to the manufacturer's instructions. The plasmids and siRNAs were transfected for 24 and 72 h, respectively, followed by the experiment.
Western blot
Whole cells and tissues were lysed in SDS-loading buffer, boiled for 10 min, and then subjected to10% polyacrylamide gel electrophoresis. The separated proteins were transferred to PVDF membranes. After blocking with tris-buffered saline with 0.1%Tween 20 (TBST) containing 5% skim milk, the membranes were incubated with primary antibodies overnight at 4°C, followed by washing with TBST thrice; the membranes were incubated with secondary antibodies for 1 h at room temperature. Proteins were detected in the X-ray film using the Image LabTM software (Bio-Rad, Hercules, CA).
Immunohistochemistry
After fixing the tissues in formalin and embedding them in paraffin, 4-μm sections of the 15 HCCs from both tumor and the adjacent nontumorous liver tissues were deparaffinized in xylene, rehydrated in an alcohol series, and washed in distilled water. Before antigen retrieval, the sections were treated with sodium citrate buffer (pH6.0) at 100°C for 20 min; activities of endogenous hydrogen peroxidase were inhibited by 0.3% H2O2. After treatment by microwave antigen retrieval, the sections were incubated with 5% FBS for 10 min at room temperature to inhibit nonspecific staining. Then, the slides were incubated with anti-HERC4 antibody in a moist chamber overnight at 4°C. The HERC4 expression score was based on the staining intensity.
Wound healing assay
Cells (4 × 105) were seeded on six-well plates and grown to 90% confluency. Then, the cells were gently scratched with pipette tip to create a mechanical wound. Images were taken at 24 h using a phase-contrast microscope. The distance of wound closure was calculated, and the p-value was calculated.
Colony assay
Cells as indicated were seeded into 10 cm plates. Ten days postseeding, the cells were fixed with 5% glutaraldehyde in phosphate-buffered saline (PBS) and stained with 0.5% Crystal violet solution. The colonies were visualized using ChampGel-1000 (Sage Creation) and counted. Each experiment was done in triplicate (Deng et al., 2010).
CCK-8
Cells were incubated in 96-well plates with 10% CCK-8 (Dojindo, Kumamoto, Japan) that was diluted in the normal culture medium at 37°C until the visual color conversion occurred. The absorbance was measured at 450 nm by a microplate reader.
Proliferation assay (EdU)
Cell proliferation was assessed by using the Cell-Light EdU DNA cell proliferation kit according to the manufacturer's instructions. 1 × 105 cells were plated into a coverslip and subjected to the EdU assay. Proliferation index was calculated as the percentage of EdU-positive cells relative to the total number cells.
Immunofluorescence assays
Cells seeded on cover glasses were fixed with 4% paraformaldehyde for 20 min at room temperature, permeabilized with 0.1% Triton X-100 for 5 min, and blocked with 10% BSA in PBS for 1 h at room temperature. Samples were incubated with anti-HERC4 at 4°C overnight. After washing with TBS thrice, sections were incubated with secondary antibody for 30 min at 37°C, followed by washing with TBS thrice; Hoechst33342 was used to stain cell nuclei (blue). Images were observed by fluorescence microscope (Nikon, 90i).
Flow cytometry assay
Cells were harvested and washed twice with cold PBS, followed by resuspending in 1 × binding buffer at a concentration of 1 × 106 cells/mL. Cells were stained with 5 μL of FITC annexin V and 5 μL PI (eBioscience) for 15 min at room temperature (25°C) in the dark, followed by analysis by flow cytometry. The experiment was repeated thrice.
Statistical analysis
Student's t-test was used to analyze the relative expression of HERC4 in human HCC and normal tissues, the relative expression of HERC4 in HepG2, SMMC7721, Hep3b, LM3, and BEL-7402 cell lines, cell proliferation and migration, and apoptosis. The differences were considered significant when p < 0.05. Graphics were analyzed with GraphPad 5.0.
Results
HERC4 highly expresses in hepatoma carcinoma
To determine the expression status of HERC4 in hepatoma carcinoma, we collected 15 primary HCC samples and tested the mRNA and protein level of HERC4 in tumor tissue and normal tissue. qRT-PCR was first performed to examine the expression pattern of HERC4 in 15 pairs of HCC specimens and adjacent nontumorous liver. Interestingly, the mRNA expression of HERC4 in tumor tissue is higher than in the adjacent nontumorous tissue (p < 0.001, Fig. 1A). Western blot analysis and immunohistochemistry results are corresponding to the qPCR results (Fig. 1B, C). In addition, we also tested HERC4 expression profiles in HCC cell lines from mRNA and protein level. The results showed that all the five HCC cell lines HepG2, SMMC7721, Hep3b, Bel-7402, and LM3 express HERC4 whether in mRNA or protein level (Fig. 1D, E). The mRNA level of HERC4 in normal human liver cell line HL7702 was significantly lower than that in HCC cell lines (Supplementary Fig. S1; Supplementary Data are available online at

HERC4 expression in HCC tissues and HCC cell lines. Fifteen HCC samples and the adjacent nontumorous tissues were isolated from the fresh tissues.
Protein detection of HERC4 knockdown cell lines
To verify if HERC4 plays a role in HCC cell proliferation and migration, we first established two HERC4 knockdown cell lines SMMC7721 and Hep3b. Western blot assay detected the efficiency of knockdown (Fig. 2A). Immunofluorescence microscopy analysis showed that the fluorescence intensity of two HERC4 knockdown cell lines SMMC7721 and Hep3b stained with anti-HERC4 was weaker compared with control cells (NC) (Fig. 2B). The results showed that we successfully established the two HERC4 knockdown cell lines.
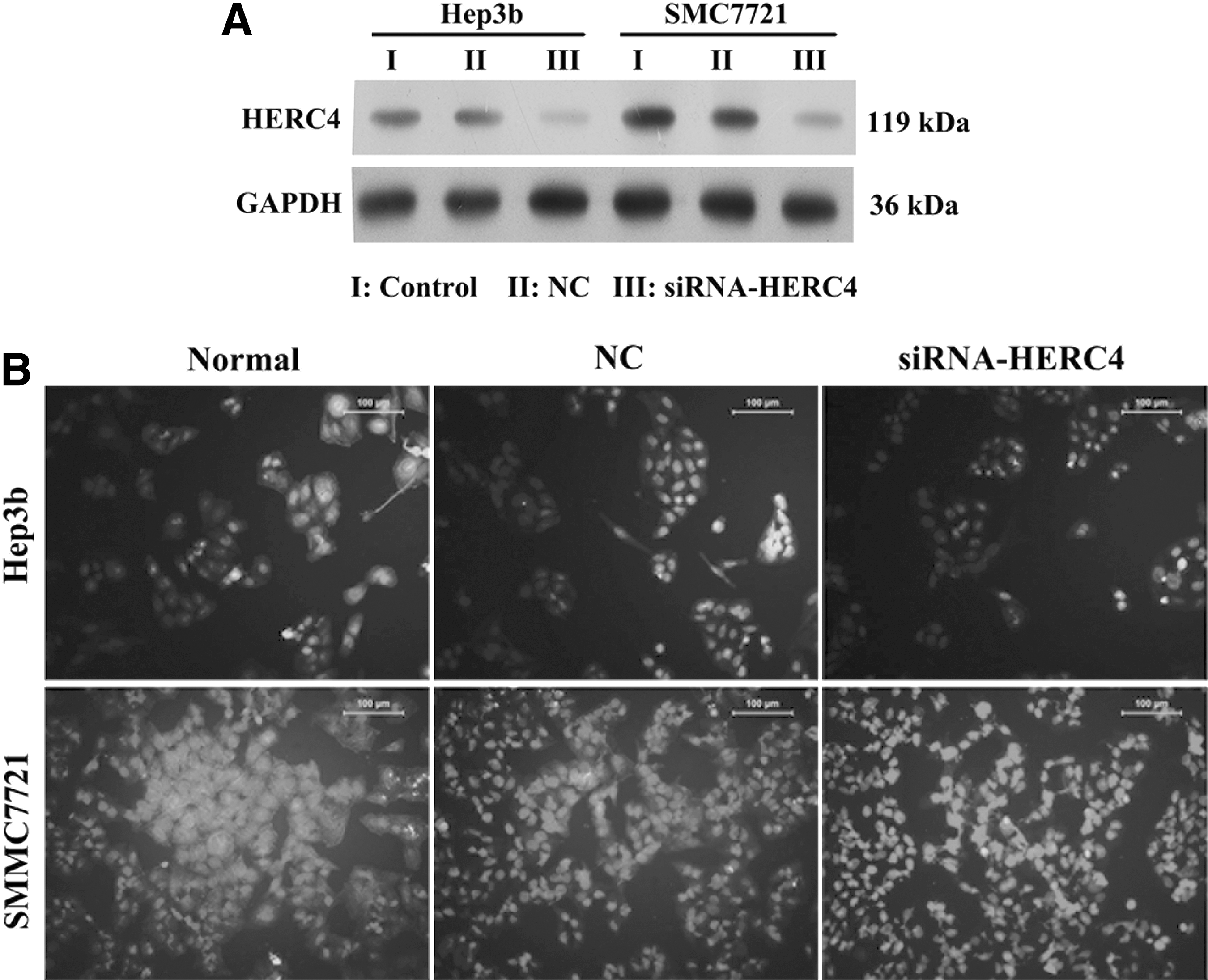
The expression of HERC4 in Hep3b and SMMC-7721 cells transfected with siRNA-HERC4 or negative control plasmid.
HERC4 knockdown inhibits HCC proliferation and migration, and induces apoptosis
To detected the ability of cell proliferation and migration in the two HERC4 knockdown cell lines SMMC7721 and Hep3b, we next did the colony formation assay, the results showed that the clone formation efficiency of both cell lines transfected with siRNA-HERC4 was dramatically reduced compared with that of cells transfected with negative control plasmid or without transfection (Fig. 3A). In addition, the CCK-8 assay testified that the proliferation ability of SMMC7721 and Hep3b cell lines was inhibited in siRNA-HERC4 cells (Fig. 3B). The wound healing assay unveiled the other function of HERC4 that the migration ability of HERC4 knockdown cells was decreased. When p < 0.001, there is a dramatically significant difference between the control group and siRNA-HERC4 group (Fig. 3C). For the EdU assay, the results showed that the EdU staining of control cells (NC) was significantly more intense compared with HERC4 knockdown cells. After transfection of si-HERC4 plasmid, the proliferation index was reduced approximately by 52% in SMMC7721 cells and 68% in Hep3b cells, respectively (Fig. 3D), suggesting that proliferative ability was blocked as a result of HERC4 knockdown. To test the effect of HERC4 on cell apoptosis, we next performed flow cytometry. From the results, we suggested that HERC4 knockdown increases cell apoptosis (Fig. 3E).
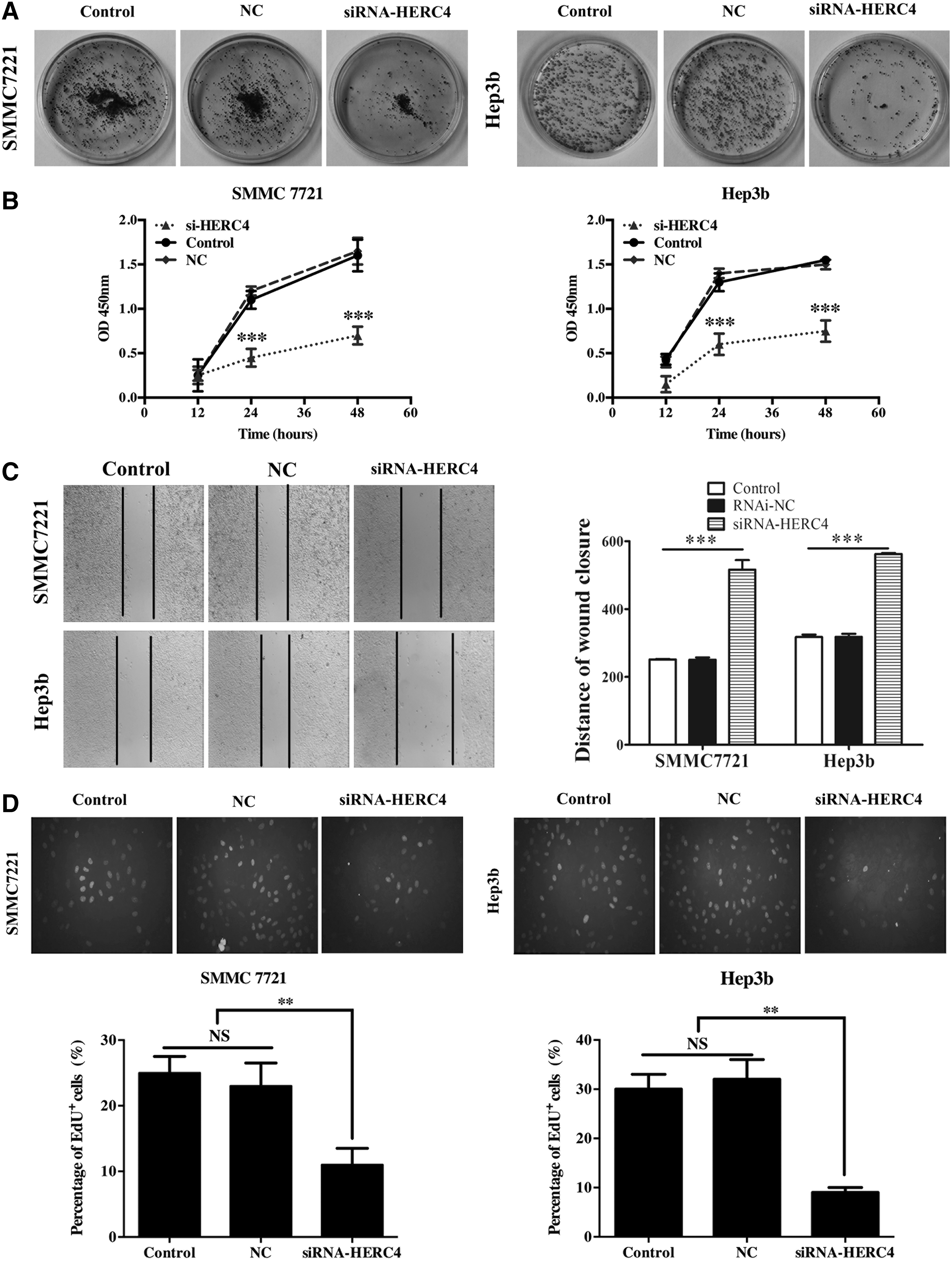
Effects of siRNA-HERC4 on cellular biological function.

Protein detection of HERC4 overexpressing cell lines
With the consequence that HERC4 knockdown suppressed cell proliferation and migration, we hypothesized that we will get the opposite results in HERC4 overexpression of HCC cells. First, we established two HERC4 overexpression cell lines Bel-7402 and LM3. Western blot assay detected the efficiency of overexpression (Fig. 4A). Immunofluorescence microscopy analysis showed that the fluorescence intensity of two HERC4 overexpressing cell lines Bel-7402 and LM3 stained with anti-HERC4 were stronger compared with control cells (NC) (Fig. 4B). Both the results above confirmed that two HERC4 overexpression cell lines were generated.

The expression of HERC4 in Bel-7402 and LM3 cells transfected with HERC4 plasmid or negative control plasmid.
HERC4 overexpression promotes proliferation and migration, and remain cell survival
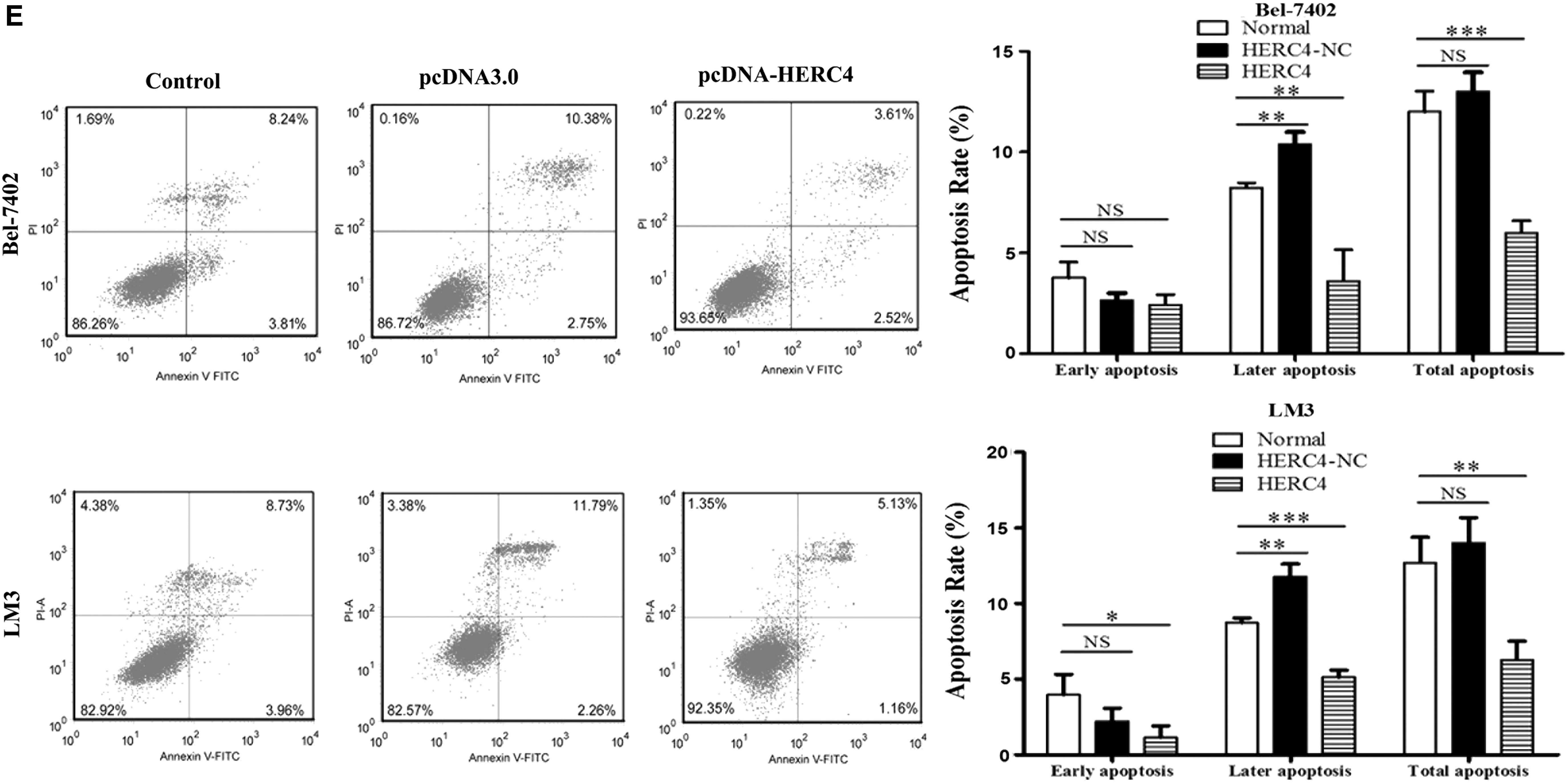
Similarly, we tested the ability of cell proliferation and migration in the two HERC4 overexpression cell lines established above. As shown in the images of colony formation assay, the colony formation efficiency of both cell lines transfected with pcDNA3.0-HERC4 was dramatically higher compared with that of control ones (Fig. 5A). In addition, the CCK-8 assay testified that the proliferation ability was promoted in HERC4 overexpressing cells (Fig. 5B). The wound healing assay unveiled the other function of HERC4 that the migration ability of HERC4 overexpressing cells was increased. When p-value is less than 0.001, there is a dramatically significant difference between the control group and the HERC4 overexpressing group (Fig. 5C). For the EdU assay, the results showed that the EdU staining of HERC4 overexpressing cells was significantly more intense compared with control cells (NC). After transfection of pcDNA-HERC4 plasmid, the proliferation index was increased approximately by 34% in Bel-7402 cells and 50% in LM3 cells, respectively (Fig. 5D), suggesting that proliferative ability was promoted owing to HERC4 overexpression. Similarly, we used flow cytometry to detect the impact of overexpressing HERC4 on cell apoptosis. We also can make a conclusion that the apoptosis rate of overexpressing HERC4 cells was significantly reduced (Fig. 5E), further remain cell survival, which is so important for tumor cells.

Effects of HERC4 overexpression on cellular biological function.
Discussion
Previous work has shown that under physiological status, HERC4 is most highly expressed in the testis, and has effects on the development of spermatozoon, which further influences fertility, implying that HERC4 is required for proper maturation and removal of the cytoplasmic droplet for the spermatozoon to become fully functional (Rodriguez and Stewart, 2007). In addition, HERC4 also functions in tumor tissues. HERC4 highly expresses both in lung cancer (Sanchez-Tena et al., 2016) and breast cancer (Deng et al., 2010), and is closely related to clinic pathological parameters, suggesting that HERC4 is likely to be a diagnostic biomarker for lung cancer and breast cancer. Besides, HERC4 interacts with transcription factor c-Maf and catalyzed its polyubiquitination and subsequent proteasome-mediated degradation, leading to inhibition of multiple myeloma proliferation (Hochrainer et al., 2008). Taken together, we know that HERC4 works in physiology and pathology. From our findings, HERC4 expresses highly in HCC and promotes HCC cell proliferation and migration in vitro, and plays a role in cell apoptosis, which are so important evidence for further studying liver cancer cell reproduction and metastasis in vivo. Our study provided a novel clue to design small molecular drugs targeting HERC4 to treat HCC. Next, we can do a deeper research to figure out the molecular mechanism of HERC4 in HCC cell proliferation and migration, and verify the biofunction of HERC4 in vitro.
In the last decade, we all know that the UPS has played a more and more important role in all regulatory processes through various signaling pathways in the cell. It not only takes the responsibility for regulating most cellular processes, including recognition and degradation of misfolded, damaged, and tightly regulated proteins, but also plays crucial roles in DNA repair, cell cycle regulation, cell migration, and the immune response (Malhi, 2015). Thus, UPS is involved in normal cellular functions and homeostasis. However, the alteration of proteasomal activity contributes to the pathological states of several clinical disorders, including inflammation, neurodegeneration, and cancer (Chen and Dou, 2010). In our study, we found that HERC4 expression in HCC tissues and HCC cell lines has an impact on cell proliferation and migration, suggesting that alterative expression of HERC4 changed cancer cell life processes. In addition, previous works unveiled that the inhibition of proteasomal activity induces G2-M cell cycle arrest and caspase-mediated apoptosis, suggesting that UPS stress sensitizes ovarian cancer cells to apoptosis (Bazzaro et al., 2006). The ubiquitin-proteasome pathway can also rapidly degrade antiapoptotic proteins, such as MCL-1 and IkB (Yang et al., 1996; Perkins, 2007), and finally produce unlimited proliferative cells. Our findings showed that the overexpression of HERC4 decreases apoptosis rate in HCC, indicating that a reduced apoptosis may play a role in remaining HCC cell survival.
The human HERC family has six members that contain a HECT domain and one or more RCC1-like domain (RLD) within their amino acid sequence. HERC2 interacts with the coiled-coil domain of USP16 through its C-terminal HECT domain (Sanchez-Tena et al., 2016). In response to DNA damage, HERC2 knockdown affects the levels of ubiquitinated H2A through the action of USP16 (Zhang et al., 2014). HERC2 can promote the recruitment of BRCA1 at DNA damage sites and has the potential to improve the BRCA1 predictive model (Bonanno et al., 2016). HERC2 can promote the translesion DNA synthesis (Mohiuddin et al., 2016) and also regulate DNA damage checkpoint by dissociating with deubiquitination enzyme USP20 (Yuan et al., 2014; Zhu et al., 2014). Therefore, HERC4 may have similar roles in the prediction and treatment of cancer-related disease.
Based on the mechanism of cancer-associated signaling pathway mediated by UPS, lots of data (Luo et al., 2009; Williams and Amon, 2009; Guo et al., 2013) suggest that cancer cells possess an elevated level of proteasome activity and are more sensitive to proteasome inhibitors than normal cells. Thus, specific molecular drugs targeting UPS, especially targeting E3 ubiquitin ligases, have been focusing on design (Nalepa et al., 2006). E6-AP is a HECT domain-containing p53 ubiquitin ligase (Scheffner et al., 1993); inhibitors of the HECT domain of E6 might prove useful in the treatment of cervical cancer (Harper et al., 2006; Lowy and Schiller, 2006; Mayeaux, 2006). In addition, downregulation of one E3s, SIAH2, is associated with resistance to endocrine therapy in breast cancer (Jansen et al., 2009). HERC4 containing the HECT domain belongs to a component of UPS (ubiquitin ligase E3). In this study, we proved that the knockdown of HERC4 decreases the capability of cell proliferation and migration and increases cell apoptosis rate, meaning that inhibition of HERC4 may become a kind of treatment for HCC, and specific molecular drugs targeting HERC4 may be designed.
In conclusion, the stabilization of some proteins concentration is essential for cell internal homeostasis. Imbalance of cell proliferation will lead to tumorigenesis and change of cell motility allows metastasis of cancer cells. HERC4 has effects on both proliferation and migration capability of cancer cells. In this study, we have demonstrated the following: (1) HERC4 is found as a relevantly upregulated gene in HCC tissues and HCC cell lines. (2) Proliferation and migration ability of HCC cells are influenced in vitro as a result of the changes of HERC4 expression. Thus, our findings provide the novel clues for HCC therapy.
Footnotes
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.