
Abstract
P-bodies are cytoplasmic foci composed of mRNAs and enzymes involved in mRNA degradation. P-bodies have been found to link to RNA interference and RNA decay mediated by microRNAs (miRNAs) and translational repression. Here, we aim to investigate different effects of overexpressed Dcp1a or GW182 on cytoplasmic aggregates formation and influence on miRNA pathway. Small RNAs were recruited into endogenous foci of P-bodies and aggregates formed by Dcpa1 and GW182 overexpression. However, only overexpressed Dcp1a but not GW182 was colocalized with DDX6, another component of P-bodies and suppressed protein translation. In addition, we investigated the relationship between stress granules and miRNA pathway and found that granules induced by G3BP1 overexpression could recruit small RNAs into the granules and repressed protein translation. As Ago2 is a key component of RNA-induced silencing complex, we also investigated the localization of endogenous Ago2 (edo-Ago2) after Dcp1a and GW182 overexpression, and found that endo-Ago2 did not colocalize with the aggregates induced by overexpression of Dcpla, GW182, and G3BP1. Notably, the ability of miRNA to regulate its target was enhanced by the granules induced by Dcp1a and G3BP1 expression. Our results suggest that overexpressed Dcp1a and GW182 can form different cytoplasmic aggregates and play distinct biological roles in the miRNA pathway.
Introduction
P-
Dcp1a is a functional partner of Dcp2 involved in decapping of 5-methylguanosine from eukaryotic mRNA and is commonly used as a marker for analysis of cellular P-body (Parker and Sheth, 2007). The 5-methylguanosine of mRNA is removed through the action of Dcp1a and Dcp2. Previous studies have showed the roles for Dcp1a in P-body formation, maintenance, and regulation. Dcp1a in P-bodies constantly and rapidly exchanges with Dcp1a in the cytoplasmic pool (Chan et al., 2013). Phosphorylation at Ser-315 suppresses the recruitment of Dcp1a into P-bodies and regulates P-body formation during translational stress (Kulkarni et al., 2010).
GW182 plays a pivotal role in siRNA- and microRNAs (miRNA)-mediated gene silencing. In both pathways, GW182, a protein with multiple glycine/tryptophan repeats, interacts with Ago2 through multiple domains to exhibit silencing activity. In siRNA-mediated gene silencing, knockdown of either GW182 or Ago2 causes loss of silencing activity with concordant P-body disassembly (Yao et al., 2013). GW182 contains two nonoverlapping repression domains that can trigger translational repression with mild effect on mRNA decay (Yao et al., 2013). Collectively, GW182 plays a critical role in miRNA-mediated gene silencing.
miRNAs are small, 21- to 23-nucleotide noncoding RNAs that target specific mRNAs for either degradation or translation repression by the RNAi pathway. A small RNA duplex generated by Dicer is subsequently loaded onto an AGO protein to form an effector complex called RNA-induced silencing complex (RISC) (Krol et al., 2010; Ayache et al., 2015). Many proteins associated with RISC and involved in miRNA-mediated suppression are concentrated into P-bodies, including four Ago proteins, Mov10, and DDX6. In addition, miRNAs are correlated with P-body formation because cellular P-bodies dispersed after knockdown of key factor of miRNA biogenesis pathway (Pauley et al., 2006).
In this study, we demonstrated that overexpression of either Dcp1a or GW182 led to the formation of different aggregates and the recruitment of miRNA mimics into these aggregates. Moreover, we investigated the correlation between stress granule (SG) and miRNA pathway, and found that miRNA also can be recruited into SGs. We demonstrated that overexpression of both Dcp1a and G3BP1 but not GW182 can suppress cellular protein synthesis. Furthermore, we showed that miRNA pathway was enhanced after miRNA was recruited into granules formed by Dcp1a and G3BP1 overexpression.
Materials and Methods
Cell culture and antibodies
HeLa and 293T cells were obtained from ATCC, and maintained in DMEM containing high glucose supplemented with 10% fetal bovine serum at 37°C in 5% CO2. Mouse monoclonal antibodies against G3BP1 and Ago2 were purchased from Abcam (Cambridge, United Kingdom), antibody against Dcp1a was purchased from Santa Cruz, rabbit polyclonal antibodies for TDP-43 and HuR were purchased from Proteintech, antibody for detecting eIF4G was purchased from Millipore, antibody against DDX6 was purchased from Bethyl Laboratories, and antibody against p38 MAKP was obtained from Cell Signaling Technology.
Plasmids and miRNA mimics
Plasmids EGFP-HsDcp1a, peGFP-hAgo2, pmyc-GFP-GW182, and pFLAG/HA-Mov10 were obtained from Addgene. G3BP1 ORF was cut from pFLAG-G3BP1 (kindly provided by Dr. Fujii) (Matsuki et al., 2013) using endonuclease KpnI and XhoI and ligated to peGFP-N3 plasmid to construct peGFP-G3BP1. pFLAG-DDX6 was kindly provided by Dr. Winkler. miRNA mimics were synthesized and labeled by Genepharma (Shanghai, China). The transfection of plasmids and miRNA mimics was performed using Lipofectamin 2000 (Life Technologies, Carlsbad). Twelve hours after plasmids transfection, cells were transfected with Cy3-labeled miRNA mimics to detect miRNA localization, or to examine silence efficiency of miRNA mimics-targeted protein by western blotting.
Western blot analysis
Cellular total proteins were prepared by lysing cells using radio-immunoprecipitation assay lysis buffer and quantified using bicinchoninic acid protein assay kit (Pierce). Proteins were then separated on SDS-PAGE after 10 mM dithiothreitol reduction and transferred to polyvinylidene difluoride membrane (Millipore, Bedford, MA). The membrane, blocked with blocking buffer (LI-COR Biosciences, Lincoln), was stained with primary antibodies, washed, and then incubated with IRDye 800 goat-antimouse IgG, or IRDye 680 donkey-antirabbit IgG (LI-COR Biosciences) at 1:10,000 dilution for 1 h. Blots were observed by scanning in an LI-COR Odyssey Infrared Imager.
Immunofluorescence assay
Cells were seeded on glass coverslips and transfected with plasmids already mentioned. The cells were then fixed in 4% paraformaldehyde for 15 min at room temperature. After the cells were washed twice with phosphate-buffered saline, they were stained with specific primary antibodies. The secondary antibodies were either Alexa Fluor 594-labeled donkey antirabbit IgG or Alexa Fluor 488-labeled goat antimouse IgG (Molecular Probes; Life Technologies) at 1:1000 dilutions. Click-iT® Plus OPP Protein Synthesis Assay Kits (Molecular Probes, Life Technologies) were used for examination of nascent protein synthesis in cells. Images were obtained with an Olympus Fluoview FV10i laser scanning confocal microscope.
Results
Overexpression of GW182, Dcp1a, or G3BP1 triggers formation of cytoplasmic aggregates
To identify the factors involved in P-body formation, a number of P-body components, GW182, Dcp1a, DDX6, Mov10, Ago2, and Pan3, were overexpressed in Hela cells and the size and the number of the cytoplasmic foci induced were measured by immunofluorescence assay (IFA). Only the overexpression of either GW182 or Dcp1a resulted in a large number of granules with larger size, whereas the overexpression of other proteins had no significant effects on both the size and the number of the cytoplasmic foci (Fig. 1A, B). As both GW182 and Dcp1a are components of P-bodies (Kulkarni et al., 2010), we investigated the colocalization of the aggregates with endogenous DDX6 (endo-DDX6), another marker for P-bodies, which suppresses translation when tethered to 3′-UTR of mRNA, and translation suppression mediated by other mechanism also requires cooperation of DDX6 (Chu and Rana, 2006; Minshall et al., 2009; Qi et al., 2012). The results demonstrated that only aggregates induced by Dcp1a overexpression but not aggregates induced by GW182 overexpression were colocalized with endo-DDX6 (Fig. 1C, upper and middle panel). Furthermore, endo-Dcp1a was not recruited into granules induced by GW182 overexpression (Fig. 1C, lower panel).

Overexpression of GW182, Dcp1a, or G3BP1 triggers formation of cytoplasmic aggregates.
SGs are cytoplasmic aggregates induced by environmental stress (Kedersha and Anderson, 2002; Kedersha et al., 2013; Protter and Parker, 2016). In certain circumstances, SG and P-bodies become associated and exchange proteins and RNAs (Kedersha et al., 2005). After overexpression of G3BP1, a marker of SGs, we found the formation of aggregates that colocalized with both eIF4G and TDP-43, two components of SGs (Anderson and Kedersha, 2008; Dewey et al., 2011; Ma et al., 2012; Fig. 1D).
Overexpression of either Dcp1a or G3BP1 inhibits protein synthesis
Both SGs and P-bodies can sequester mRNAs and translation initiation complex to suppress protein synthesis (Kulkarni et al., 2010; Stoecklin and Kedersha, 2013). We speculate that the aggregation induced by either G3BP1 or Dcp1a overexpression may inhibit protein translation. Cells were transfected with either Dcp1a or G3BP1 expression plasmids, and nascent protein synthesis was monitored 24 h post-transfection. The results showed that nascent protein synthesis was almost completely suppressed in HeLa cells transfected with Dcp1a or G3BP1 expression plasmid (Fig. 2C–E). In contrast, cells transfected with GW182 expression or empty plasmid had normal protein translation (Fig. 2A, B).
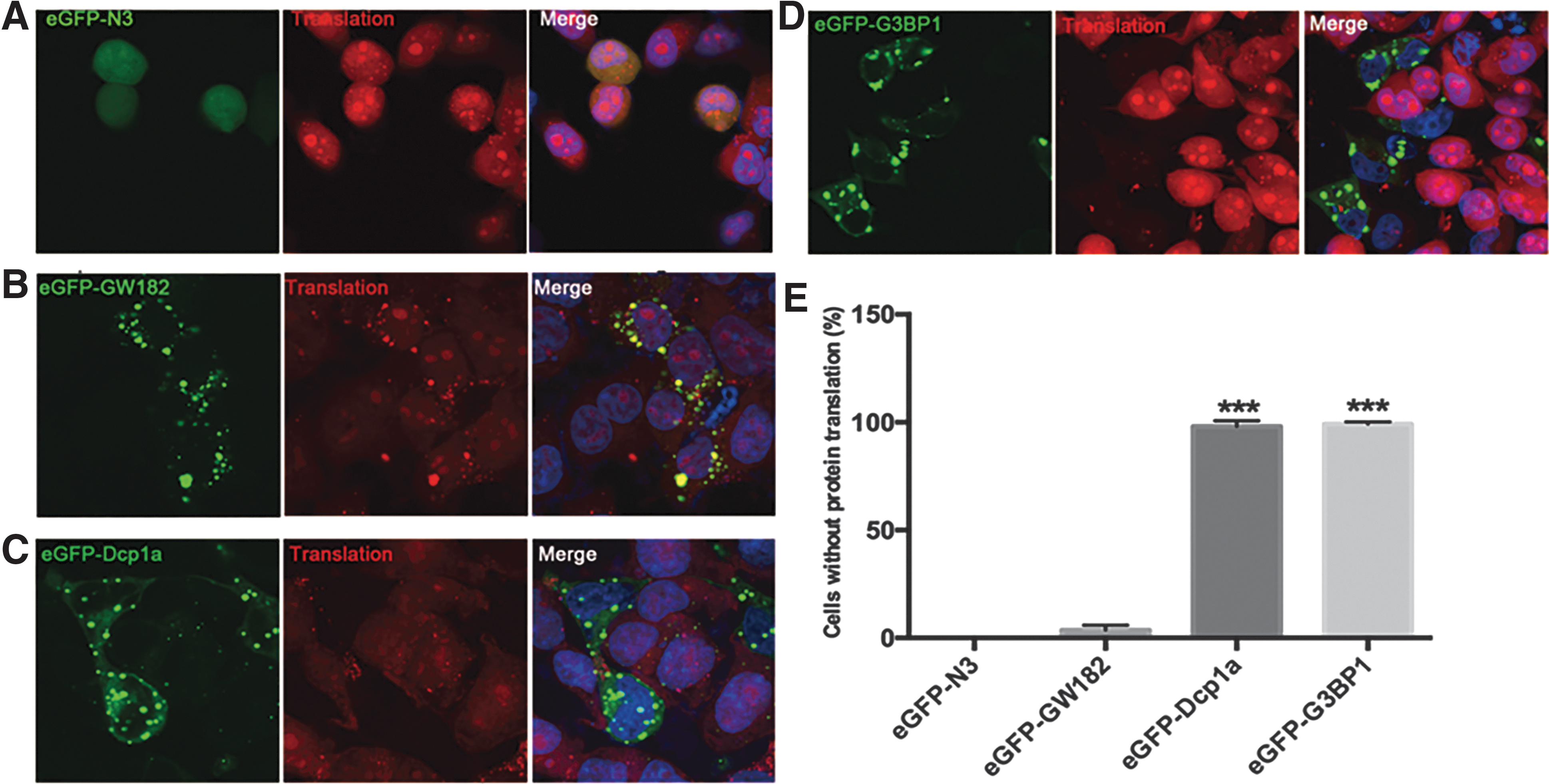
Overexpression of either Dcp1a or G3BP1 inhibits protein synthesis.
miRNAs are recruited into granules induced by overexpression of Dcp1a, GW182, and G3BP1
RISC can localize to P-bodies and SG (Liu et al., 2005; Sen and Blau, 2005; Leung et al., 2006). To investigate whether miRNAs are associated with the aggregates induced by the overexpression of Dcp1a, GW182, and G3BP1, we transfected HeLa cells with Dcp1a, GW182, or G3BP1 expression plasmids then Cy3-labeled miRNA-141, and the cells were analyzed 4 h later by IFA. Cy3-miRNA-141 was detected within the granules induced by the overexpression of either proteins (Fig. 3A). Furthermore, we cotransfected with plasmid pEGFP-HsDcp1a and another Cy3-labeled miRNA mimics, miRNA-30a, and found that miRNA-30a was recruited into granules formed by the overexpressed Dcp1a (Fig. 3B). To further confirm that Cy3-miRNA-141 can also be recruited into endo-P-bodies, we analyzed the P-bodies using a specific antibody for Dcp1a and found that miRNA was localized in the endo-P-bodies soon after being transfected into the cells (Fig. 3C). Ago2 is the key component of RISC (Ghildiyal and Zamore, 2009; Filipowicz et al., 2010), and overexpressed Ago2 was colocalized with TIA1, another marker of SG (Sen and Blau, 2005; Leung et al., 2006). We determined whether endo-Ago2 can also be recruited into granules formed by these overexpressed proteins. Different from overexpressed Ago2, endo-Ago2 did not colocalize with Dcp1a, GW182, and G3BP1 (Fig. 3D).
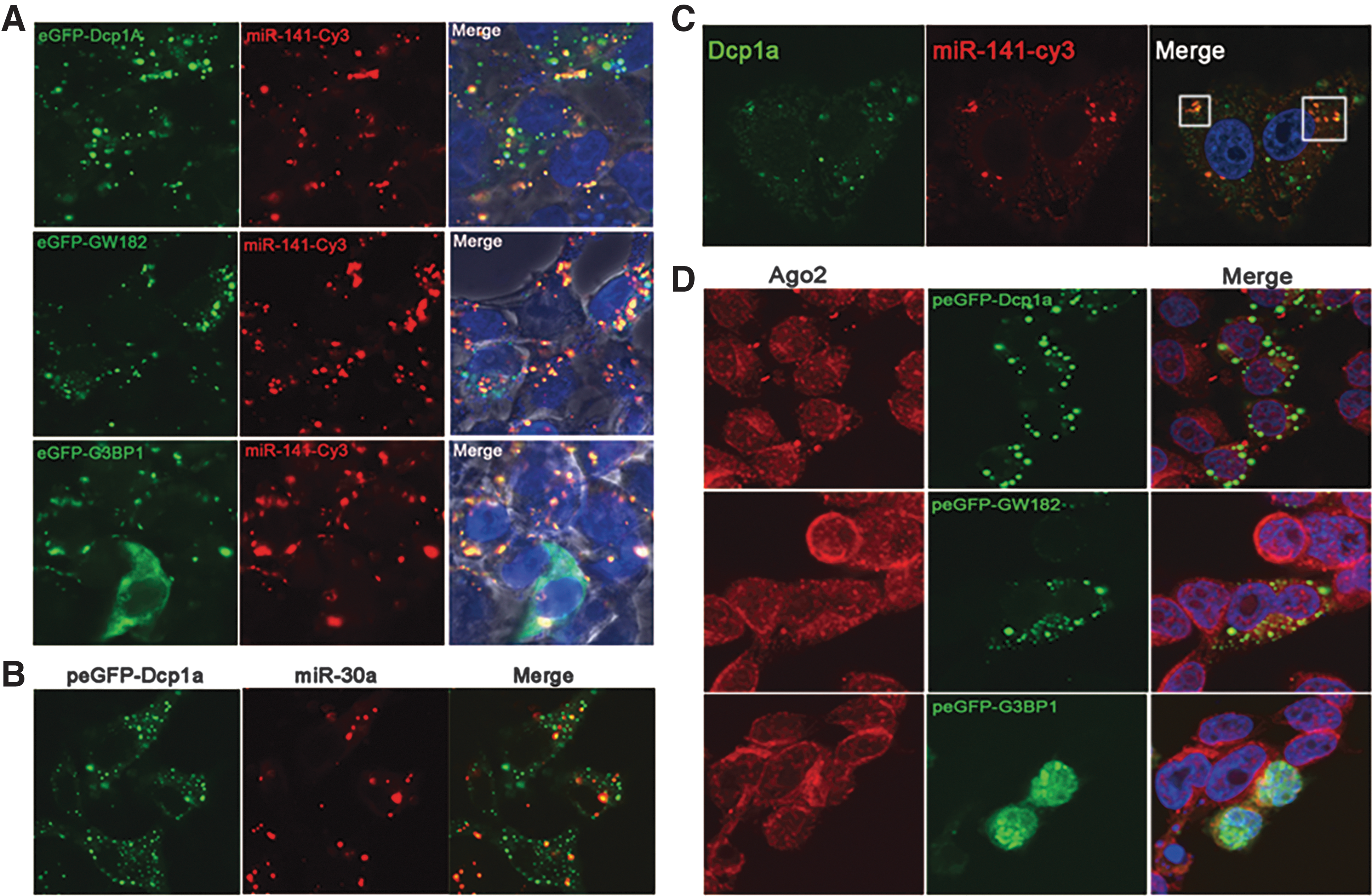
miRNAs are recruited into granules induced by overexpression of Dcp1a, GW182, and G3BP1.
Granules formed by overexpressed Dcp1a and G3BP1 regulate the targeting of miRNAs to their targets
To investigate whether granules induced by the overexpressed Dcp1a, GW82, and G3BP1 influence the miRNA pathway, we first transfected 293T cells with the respective overexpression plasmids, and then with miR-30a and miR-141 mimics. Forty-eight hours later we detected the expression of Beclin1, one of the targets of miR-30a (Zhu et al., 2009), and p38 MAPK, which is proven to be a miR-141 target (Mateescu et al., 2011; Karginov and Hannon, 2013). Western blot analysis demonstrated that overexpression of either G3BP1 or Dcp1a but not GW182 enhances miRNAs' function to suppress the targets (Fig. 4A). To further confirm the specificity, we also transfected scrambled miRNA after plasmid transfection and found that the expression of both Beclin1 and p38 MAPK was not influenced by the Dcp1a, GW182, and G3BP1 overexpression (Fig. 4B).
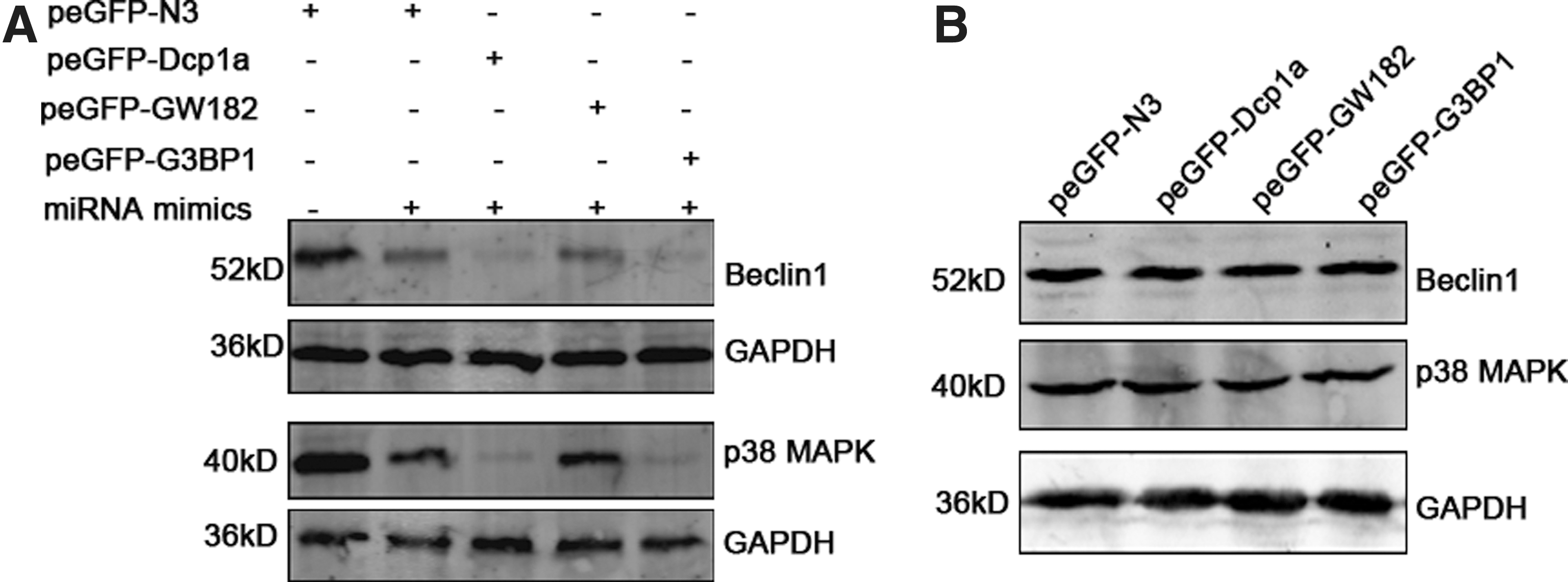
Granules formed by overexpressed Dcp1a and G3BP1 regulate the targeting of miRNAs to their targets
Discussion
P-bodies contain a large number of different proteins, some of which are essential whereas some of which are dispensable for P-body formation (Kulkarni et al., 2010). It is also shown that there is another type of aggregates called GW-bodies existing in the cytoplasm, which contains GW182 in addition to other decay factors shared with P-bodies (Yang et al., 2004). Evidence has demonstrated that P-bodies and GW-bodies have different dynamic response to cycloheximide treatment and the cell cycle (Yang et al., 2004; Aizer et al., 2008). Also, it is found that the two aggregates showed different functions in that P-bodies mainly function as RNA decay and translational suppression, whereas GW-bodies play roles as repositories for translationally silencing RNAs, but not as active translational silencing (Patel et al., 2016). In this study, we overexpressed six P-body components and found that they have different aggregation properties. Overexpressed Dcp1a and GW182 aggregated to form granules with larger number and size, whereas other overexpressed proteins only formed a few and small aggregates. DDX6, also named RCK/p54, was reported to be pivotal for P-body formation, and P-body formation was disrupted after DDX6 deletion by the RNAi method (Chu and Rana, 2006; Perez-Ortin et al., 2013; Ayache et al., 2015). However, our results demonstrated that overexpressed DDX6 did not induce granule formation, suggesting that unlike Dcp1a and GW182, excessive DDX6 in the cytoplasm cannot induce cytoplasmic aggregates formation.
As Dcp1a and GW182 are both proven components of P-bodies (Kulkarni et al., 2010), we also investigated whether the aggregates formed by Dcp1a and GW182 colocalize with other endocomponents of P-bodies. The results demonstrate that only granules induced by overexpressed Dcpla colocalize with endo-DDX6, whereas granules induced by overexpressed GW182 did not colocalize with endo-DDX6 or endo-Dcp1a, indicating that the composition of these aggregates induced by Dcp1a and GW182 overexpression is distinct.
It has been reported that protein synthesis can be suppressed after RNA granule formation, including P-bodies and SG (Anderson and Kedersha, 2009). We investigated protein translation upon the formation of granules and found that only aggregates induced by Dcp1a and G3BP1 could inhibit nascent protein synthesis, whereas granules induced by GW182 could not. These data suggest that the aggregates induced by overexpressed Dcp1a and GW182 have different properties.
Evidence has shown that miRNA-related RISC and siRNA-related RISC are localized to RNA granules, such as Ago2, the key protein of miRNA pathway, miRNAs, and its target mRNA (Liu et al., 2005; Sen and Blau, 2005; Leung et al., 2006; Detzer et al., 2011; Ma et al., 2012). We, therefore, investigated the correlation between the granules induced by overexpressed Dcp1a, GW182, and G3BP1, and miRNA pathway-related protein Ago2 and miRNAs. The results demonstrated that miRNA mimics can be recruited into the granules induced by each of the three overexpressed proteins. However, different from the previous observation (Liu et al., 2005; Sen and Blau, 2005; Leung et al., 2006; Detzer et al., 2011), we found that endo-Ago2 does not localize into these granules. In addition, we also showed that granules induced by the overexpressed Dcp1a and G3BP1 facilitated miRNA to suppress its target.
Conclusion
In conclusion, this study further confirmed previous observations that P-bodies and GW-bodies represent two different aggregates of nontranslation RNAs, and have different influences on cellular bioactivities, such as miRNA pathways. Future characterizations of the RNAs and protein contents of these aggregates will provide valuable insights into the functional role of these structures.
Footnotes
Acknowledgments
This work was supported by The National Key Research and Development Program of China (2016YFC1201000), the Major Research and Development Project from the Ministry of Health of China (2016ZX10001005003), and Fundamental Research for the Central Universities (021414380213).
Disclosure Statement
The authors have nothing to disclose.