
Abstract
Parkinson's disease (PD) is a serious neurodegenerative disorder that lacks effective therapeutic methods. In this research, expressions of PPARα, RXRα, and miR-21 were evaluated in PD patients and normal controls. To investigate the effects of miR-21, docosahexaenoic acid (DHA) and aspirin (ASA) on PD, as well as the relationships between them, SH-Y5Y cells were treated with DHA, ASA, or both for 24 h. The assay showed that levels of miR-21 were increased and levels of PPARα were decreased in PD patients compared with normal controls. miR-21 was negatively correlated with PPARα in PD patients. DHA and ASA could activate RXRα and PPARα, respectively. Additionally, DHA upregulated PPARα expression by inhibiting miR-21 in SH-Y5Y cells. A combination of DHA and ASA efficiently enhanced heterodimer formations of PPARα and RXRα and increased the expression of neurotrophic factors PSD-95, brain-derived neurotrophic factor (BDNF), and glial cell-derived neurotrophic factor (GDNF), while inhibiting NFκB and COX2. These findings suggest that a combination of DHA and ASA could significantly improve the expression of PSD-95, BDNF, and GDNF by promoting heterodimerization of PPARα and RXRα, thus supplying a new therapeutic method for PD.
Introduction
P
There are many inherent difficulties in exploring the pathogenesis of PD because this disease occurs in the CNS. Therefore, research tends to focus on other methods of diagnosis and treatment. Some biological molecules, including proteins in peripheral blood lymphocytes or metabolomics in serum, are biomarkers of PD (Scherzer et al., 2007; Bogdanov et al., 2008). Many molecules are involved in PD; some are neuroprotective and others are neurodegenerative. For example, fibroblast growth factors can protect dopaminergic neurons (Walker et al., 1998). Fractalkine functions as a protective protein by inhibiting microglia activation (Pabon et al., 2011). However, ring finger protein 11 colocalized with Lewy bodies promotes the development of PD.
A class of small RNAs called microRNAs (miRNAs) is also involved in PD. Some miRNA expressions are changed in the blood or brain of PD patients. miRNA is noncoding and regulates gene expression posttranscription by targeting specific mRNA. miRNAs gradually develop into potential therapeutic targets. In PD, miR-7 and miR-153 can bind to the 3′UTR of a-synuclein mRNA and downregulate a-synuclein expression relating to Lewy bodies (Junn et al., 2009; Doxakis, 2010). miR-34 is reduced in PD patients compared with healthy controls (Minones-Moyano et al., 2011). Functional research of miR-21 focuses on tumors. miR-21 plays a key role in human tumor progression and is considered an onco-miR (Pan et al., 2010). It is reported that miR-21 enhances axon growth and regulates growth pathways following peripheral nerve injury (Strickland et al., 2011). However, its specific role in the nervous system is still largely unknown.
Docosahexaenoic acid (DHA), an n−3 long-chain polyunsaturated fatty acid (n−3 LCPUFA), plays a critical role in the preservation of the nervous system and is involved in the treatment of neurodegenerative diseases (Ikemoto et al., 2001). DHA is highly concentrated in the nervous system, but its levels are reduced in neurodegenerative diseases. Some research has found that DHA shows a neuroprotective effect in Alzheimer's disease (Chua et al., 2014), but the effects of DHA on PD are still unclear.
Some nonsteroidal anti-inflammatory drugs (NSAID) could specifically bind to and activate PPARα. Aspirin (ASA) is the most common NSAID drug used for treating pyrexia, pain, inflammation, and antiplatelet. It also possesses some newly discovered benefits, including neuroprotective and cancer-inhibiting effects (Grilli et al., 1996). ASA could upregulate and activate PPARα in macrophages (Paganini-Hill et al., 1992). However, whether ASA modulates PPARα in neurons is still being explored.
In our research, we found that miR-21 was higher and PPARα was lower in PD patients than in normal controls. We found that DHA activated RXRα and ASA activated PPARα in human neuroblastoma cell lines SH-Y5Y. Otherwise, DHA could enhance PPARα protein expression by inhibiting miR-21 expression. When SH-Y5Y cells were treated with both DHA and ASA, neurotrophic factors, postsynaptic density protein 95 (PSD-95), brain-derived neurotrophic factor (BDNF), and glial cell-derived neurotrophic factor (GDNF) were increased, whereas caspase-3, pERK1/2, and COX-2 were decreased. Combined, these results indicate that a combination of DHA and ASA could modify PD by activating RXRα and PPARα inhibiting NFκB.
Materials and Methods
Ethics statement
Fifteen PD patients' blood samples were obtained from Inner Mongolia Autonomous Region People's Hospital, China. Fifteen healthy controls were volunteers in Huhhot City, Inner Mongolia, China. All experiments were approved by the Ethics Committee of Wuhan University. The investigations were carried out following the rules of the Declaration of Helsinki of 1975 revised in 2008.
Cell line and reagents
SH-Y5Y cells were cultured in a DMEM medium with
Anti-human RXRα polyclonal antibody (ab80453) and anti-human PPARα polyclonal antibody (ab8934) were purchased from Abcam (Cambridge, MA). The LanthaScreen TR-FRET Peroxisome Proliferator-Activated Receptor Alpha Coactivator Assay Kit (Thermo Fisher, PV4684), the LanthaScreen TR-FRET Retinoic X Receptor Alpha Coactivator Assay Kit (PV4797; Thermo Fisher, CA), and all of the antibodies for western blot detection were purchased from Abcam. miR-21 mimics (5′-AACAUCAGUCUGAUAAGCUAUU-3′), miR-21 inhibitors (5′-UCAACAUCA-GUCUGAUAAGCUA-3′), and negative controls (5′-CAGUACUUUUG-UGUAGUACAA-3′) were obtained from Ambion, Inc. (Austin, TX; Liu et al., 2013).
Quantitative real-time polymerase chain reaction
Peripheral blood mononuclear cells (PBMC) were isolated from experimental subjects using Ficoll-Paque Plus (Amersham Biosciences, Uppsala, Sweden). Total RNA was extracted from PBMC using TRIzol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. Subsequently, 2 μg of total RNA was transcribed using the TaqMan MicroRNA Reverse Transcription Kit (Applied Biosystems, CA). Quantitative real-time polymerase chain reaction (qRT-PCR) was executed using specific TaqMan MicroRNA analysis through Applied Biosystems StepOnePlus Real-Time PCR System. The quantity of mature miR-21 was calculated using 2−ΔΔCt method. U6 was used as an internal control gene to normalize the data in this experiment. Specific primers were used as follows. miR-21, forward: 5′-CACCCCCGTCCCCTCCAA-3′; reverse: 5′-GATCCCAGGTTGGCGGGC-3′ (Chen et al., 2013); U6, forward: CTCGCTTCGGCAGCACA; reverse: AACGCTTCACGAATTTGCGT (Song et al., 2014). RT-PCR results were calculated, statistically analyzed using GraphPad Prism 5 software. All experiments were performed at least three separate times in triplicate.
Enzyme-linked immunosorbent assay
For analysis of human PPARα and RXRα in peripheral blood, PBMC were isolated, counted, and adjusted to a concentration of 5 × 106 cells/mL. Then, 1 × 107 cells in 0.5 mL of phosphate-buffered saline were frozen and thawed three times to lyse PBMC completely. The cell lysate was centrifuged at 10,000 rpm at 4°C to remove cell precipitation. The supernatant was collected and used for enzyme-linked immunosorbent assay (ELISA) detection.
First, 100 μL of supernatant of PBMC lysis was added into an ELISA 96-well plate (Costar) at 4°C overnight. Then, 5% of BSA was used to block the ELISA plate at 37°C for 1 h. Primary antibodies of PPARα and RXRα (1:1000) were added after washing with phosphate-buffered saline tween (PBST) and incubated for 1 h at 37°C. After washing with PBST, HRP goat anti-rabbit IgG (1:4000) was added and incubated for 1 h.
PPARα and RXRα-binding assays
DHA and ASA binding to the ligand-binding domain of RXRα and PPARα was assessed, respectively, in a LanthaScreen TR-FRET Peroxisome Proliferator-Activated Receptor Alpha Coactivator Assay and LanthaScreen TR-FRET Retinoic X Receptor Alpha Coactivator Assay, according to the protocol of the manufacturer. EC50 values for the RXRα and PPARα were determined using GraphPad prism 5.0 software.
Luciferase activity assay
A total of 1 × 105 of SH-Y5Y cells were seeded in six-well plate and cultured for 24 h. Then, 2 μg of PPRE (PPARα response element)-luc plasmid and pcDNA3.1-PPARα were cotransfected into the cells with Lipofectamine 2000. After 48 h, the transfected cells were stimulated with different concentrations of DHA (0, 0.1, 1, 10, 100, and 1000 μM) for another 24 h. The cells were then lysed, and luciferase activity was determined using a Dual-Luciferase Reporter Assay System (Promega) according to the manufacturer's instructions.
The same method was used to detect the effect of ASA on RXR. SH-Y5Y cells were maintained in a six-well plate. A luciferase reporter plasmid containing RXRα response element (RXRE) and pcDNA3.1-RXRα was used to transfect SH-Y5Y cells. Then, transfected cells were treated with ASA (0, 0.1, 1, 10, 100, 1000 μM) for 24 h. The Dual-Luciferase Reporter Assay was performed. The luciferase activity expression was normalized to Renilla activity. At least three independent experiments were carried out in triplicate. The value was expressed as the mean ± standard deviation.
According to the manufacturer's instructions, the LanthaScreen TR-FRET Peroxisome Proliferator-Activated Receptor Alpha Coactivator Assay Kit and the TR-FRET Retinoic X Receptor Alpha Coactivator Assay Kit were used to test whether DHA and ASA were agonists of RXRα and PPARα.
For the pGL3-PPARα-3′UTR luciferase assay, SH-Y5Y cells were cultured in 12-well plates for 24 h. The cells were cotransfected with pGL3-PPARα-3′UTR plasmid and miR-21 mimics or miR-21 inhibitors. After 24 h, a Dual-Luciferase Reporter Assay was performed.
Western blot analysis
SH-Y5Y cells were cultured in 12-well plates and treated with DHA (100 μM) or ASA (100 μM) for 48 h. Cells were harvested and lysed using cell lysis buffer (20 mM Tris, 150 mM NaCl, 0.5% Triton X-100, 1% NP-40, 1% protease inhibitor). Then, 20 μg of protein was used for western blotting. Anti-human RXRα antibody (1:1000) and anti-human PPARα (1:1000) antibody were used as primary antibodies. HRP goat anti-rabbit IgG (1:3000) was used as a secondary antibody. Lamin B (Cell Signaling Technology, Boston, MA) was used as a loading control for the detection of the transcription factors.
For detecting proteins involved in signaling pathways, SH-Y5Y cells were treated with DHA, ASA, or both for 48 h. Additionally, SH-Y5Y cells were transfected with plasmid pcDNA3.1-PPARα. The cells were then lysed and used for detecting PTEN, pAKT, Caspase-3, p65, COX-2, pERK1/2, PSD-95, BDNF, and GDNF by western blotting. The western blot signals were analyzed using MCID Elite software. The baseline density of each band was subtracted from band density peak. The signal intensity was estimated by mean density value of the sampled peak.
Coimmunoprecipitation assays
Coimmunoprecipitation (Co-IP) assays were employed using the CoImmunoprecipitation Kit (Thermo Scientific, MA) according to the manufacturer's protocol. SH-Y5Y cells were cultured in a six-well plate with 2 × 105 cells per well. The cells were transfected with miR-21 mimics, miR-21 inhibitors, and negative controls, or stimulated with DHA and ASA. After 48 h, nuclear extractions from the SH-Y5Y cells were immunoprecipitated with anti-RXRα antibody. PPARα was analyzed using the immunoblotting procedures described above.
Results
PPARα is negatively correlated with miR-21
Expressions of miR-21, PPARα, and RXRα were examined in PD patients using qRT-PCR and ELISA. miRNA levels of miR-21 were higher in PD patients than in normal controls (Fig. 1A). ELISA results showed that expression of PPARα was much lower in PD patients than in normal controls (Fig. 1C), but no significant difference in the expression of RXRα was observed (Fig. 1B). Correlation analysis was performed to analyze the relationships between miR-21 and PPARα, miR-21 and RXRα. miR-21 is negatively correlated with PPARα (R2 = 0.7406), but has no significant relationship with RXRα in patients with PD (Fig. 1D, E).
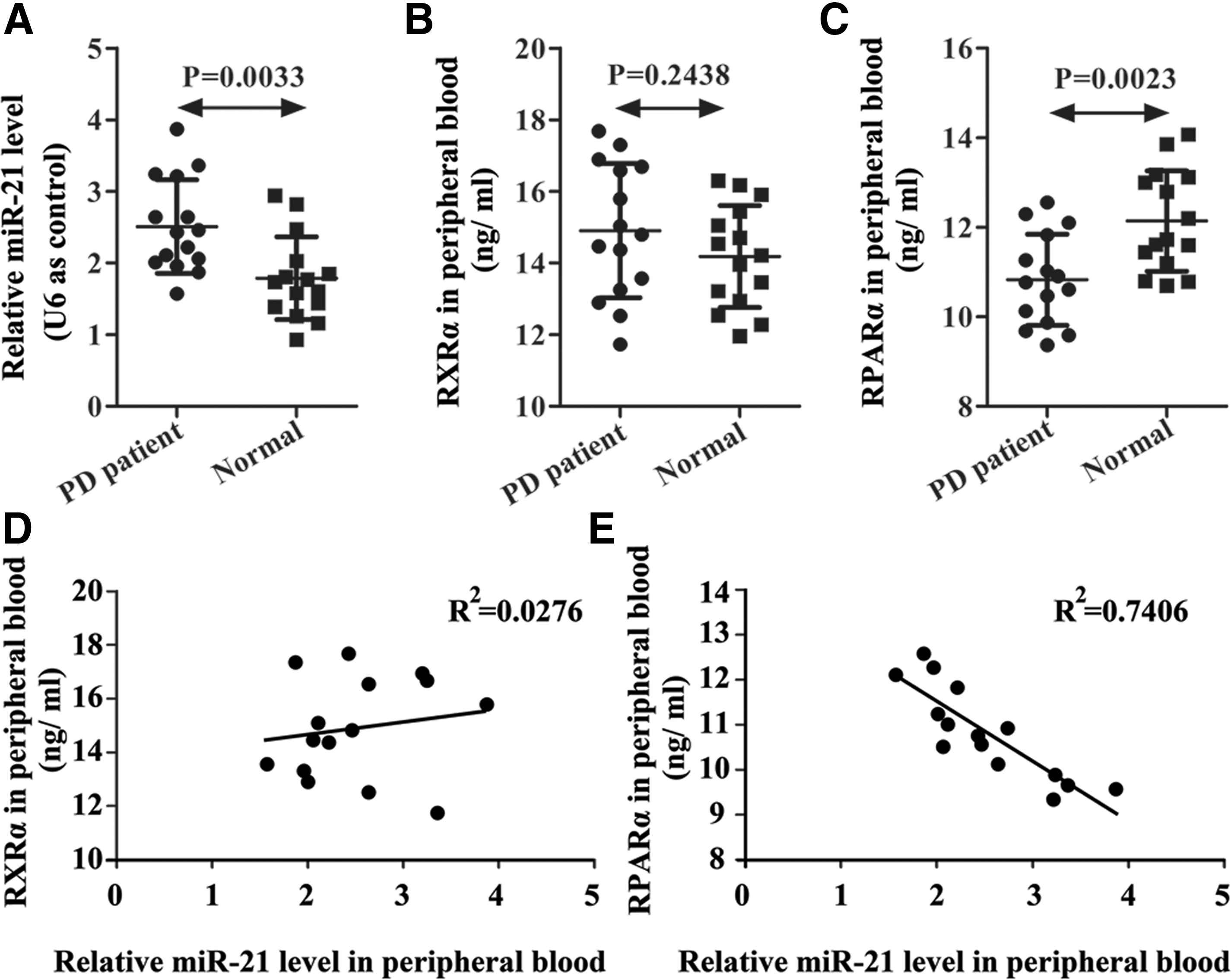
Expression of miR-21, RXRα, and PPARα in PD patients and normal healthy controls.
DHA is an agonist of RXRα, whereas ASA is an agonist of PPARα
DHA and ASA are both reported to protect neurons. To explore the targets of DHA and ASA, the LanthaScreen TR-FRET PPAR Alpha Coactivator Assay Kit and the LanthaScreen TR-FRET RXR Alpha Coactivator Assay Kit were used in this study. The results suggest that DHA and ASA were agonists of RXRα and PPARα, respectively. The EC50 for DHA binding was 10.75 μM for RXRα, and the EC50 for ASA binding was 3.04 μM for PPARα (Fig. 2A, C). To further support this result, luciferase activity was detected. DHA could enhance RXRα binding to RXRE (Fig. 2B). Similar results were obtained in ASA. ASA increased PPARα binding to PPRE in a dose-dependent manner (Fig. 2D). Additionally, PPARα protein expression could be elevated by DHA. However, ASA could not change PPARα protein levels, suggesting that ASA increased the activity of PPARα, but not the protein quantity (Fig. 2E).

DHA and ASA were agonists of RXRα and PPARα, respectively.
We next determined the mechanism of DHA for inducing PPARα expression. SH-Y5Y cells were treated with DHA for 48 h and miR-21 was examined by qRT-PCR. It was found that DHA inhibited miR-21 in a dose-dependent manner (Fig. 2F). These results suggest that DHA not only enhances PPARα activity, but also increases PPARα protein expression by inhibiting miR-21 and that ASA could stimulate RXRα activity.
DHA promotes PPARα by inhibiting miR-21
To identify the target of miR-21, miR-21 mimics or miR-21 inhibitors with PGL3-PPARα-3′UTR were cotransfected into SH-Y5Y cells. miR-21 mimics effectively decreased the luciferase activity compared with negative miRNA, whereas miR-21 inhibitors increased the luciferase activity, suggesting that miR-21 could target 3′UTR of PPARα (Fig. 3A, B). Western blot results indicated that miR-21 reduced the expression PPARα, but miR-21 inhibitor enhanced the expression PPARα (Fig. 3C). When cells were treated with DHA, with or without ASA, PPARα was significantly increased in both DHA- and ASA-treated cells. DHA induced PPARα expression enormously (Fig. 3D). Meanwhile, miR-21 mimics inhibited the heterodimer formation of PPARα and RXRα (Fig. 3E). However, DHA, but not ASA increased the heterodimer formation of PPARα and RXRα (Fig. 3F). Together, the results illustrate that DHA could enhance the expression of PPARα through the inhibition of miR-21.

Effects of miR-21 and DHA on expression of PPARα.
DHA and ASA influence diversified signaling pathways
PPARα is a critical factor in regulating proliferation and inflammation of neurons and also protects neurons by promoting neurotrophic factor expressions. SH-Y5Y cells were transfected with pcDNA3.1-PPARα, and western blotting was performed. The results suggest that PPARα was able to downregulate PTEN, pAKT, Caspase-3, NF-KB p65, pERK1/2, and COX-2 and upregulate PSD-95, BDNF, and GDNF (Fig. 4A).

Effects of DHA and ASA on neuroapoptosis, neuroinflammation, and neuroprotection as related to protein expression.
In the present research, DHA was used to stimulate SH-Y5Y cells. Proteins related to apoptosis, inflammation, and neurotrophic factors were identified. DHA downregulated Caspase-3 expression by promoting PTEN and prohibiting phosphorylation of AKT. DHA also decreased the expression of NFκB p65, phosphorylation of ERK1/2 and COX-2, which suggests that DHA could inhibit the development of inflammation. Importantly, DHA increased neurotrophic factors such as PSD-95, BDNF, and GDNF, and functioned as a neuroprotective factor (Fig. 4B).
These data reveal that the combination of DHA and ASA is a mechanism for improving PD. DHA affected PPARα by activating RXRα and promoting expression of PPARα through inhibiting miR-21, and ASA could activate PPARα. The functions of ASA and DHA significantly increased heterodimer formation of PPARα and RXRα and improved their ability to enter the nucleus (Fig. 4C). After entering the nucleus, the heterodimers exhibited regulating functions in proteins involved in apoptosis, inflammation, and neurotrophy (Fig. 4D).
Discussion
In the current research, we studied a relationship between DHA and PPARα/RXRα, ASA and PPARα, and the function of DHA and ASA combination on SH-Y5Y cells. We identified that DHA reduction of miR-21 promotes PPARα binding with RXRα. DHA and ASA could effectively enhance the activities of RXRα and PPARα, respectively. When DHA and ASA were combined to treat SH-Y5Y cells, it was found that heterodimers of PPARα and RXRα promoted PTEN and neurotrophic factors and inhibited inflammation factors (as summarized in Fig. 4). miR-21 acts as an epigenetic mediator for protecting neuron cells SH-Y5Y in response to DHA.
PPARα showed a protective function in PD (Chaturvedi and Beal, 2008). Furthermore, PPAR agonists activating PPAR could inhibit inflammation and apoptosis by reducing expression of COX and NFκB (Chaturvedi and Beal, 2008), a finding corroborated by our results.
Usually, miR-21 is regarded as an onco-miR that regulates tumor growth and carcinogenesis in response to chemotherapy (Jiang et al., 2015). In this study, we found that miR-21 was implicated in PD. miR-21 in PD was higher than in normal controls, and it was negatively correlated with PPARα. We predicted that the target of miR-21 using software Targetscan Human 6.2. PPARα was regulated by miR-21 targeting regions 898–904 and 1039–1045 3′UTR. miR-21 targeting PPARα has been previously reported in the work by Shu Chien (Zhou et al., 2011). miR-21 mimics efficiently reduced PPARα expression and inhibited heterodimers with retinoid-X receptors. PPARα and RXRα heterodimers regulate the gene for cell growth and differentiation.
DHA is essential for brain maturation and promotes the activity of RXRα (de Urquiza et al., 2000). RXRα was activated by its ligand, which can increase DA neuron's chances of survival and proliferation by formation of Nurr1-RXR heterodimers (Wallen-Mackenzie et al., 2003). In this study, we introduce a TR-FRET system to study the relationship between DHA and RXRα. It has been identified that DHA is an activator of RXRα. Subsequently, when DHA is used to stimulate SH-Y5Y cells transfected with pcDNA3.1-RXRα and RXRE reporter plasmid, it can promote luciferase activity, suggesting that DHA increases the RXRE-dependent transcription. We also found that DHA can enhance PPARα expression through inhibiting miR-21 in SH-Y5Y cells. Therefore, DHA is able to promote PPARα/RXRα by both activating RXRα and elevating PPARα.
NSAID are believed to reduce the risk of neurodegenerative disorders (Hirohata et al., 2005; Rosenberg, 2005). Indomethacin, ibuprofen, naproxen, and fenoprofen can specifically bind to and activate PPARα and/or PPARγ (Sastre et al., 2006; Heneka et al., 2007). Pioglitazone protects animals from neuronal cell death in a 1-methyl-4- phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced PD model (Wahner et al., 2007). ASA is a common NSAID. It is reported that ASA can upregulate PPARα/γ (Yiqin et al., 2009). In our research, ASA activates PPARα in SH-Y5Y cells, but does not affect protein expression. This result is consistent with research by Joan (Planaguma et al., 2002). A combination of DHA and ASA further increases dimerization of PPARα and RXRα. Activation of PPARα and RXRα promotes the entering of nucleus and the formation of heterodimers. PPARα and RXRα heterodimers play important roles in regulating neuron apoptosis, inflammation, and neuroprotection. They can inhibit inflammation by prohibiting NFκB, pERK1/2, and COX-2 and promote neurotrophic factors PSD-95, BDNF, and GDNF.
Conclusions
Our results suggest that DHA and ASA could protect neurons through their functions on PPARα and RXRα, thus indicating therapeutic potential for targeting PPARα and RXRα and improving PD.
Footnotes
Acknowledgments
This study was supported by the Key Project of the National Natural Science Foundation of China (81230031/H18) and Hubei Province's Outstanding Medical Academic Leader Program. The authors thanked Hong Chang, Department of Neurology, Inner Mongolia Autonomous Region People's Hospital, for the patients' sample collection.
Disclosure Statement
No competing financial interests exist.