
Abstract
DNA methyltransferase 1 (DNMT1) and miRNAs are both important regulators of gene expression that have been implicated in the pathogenesis of atherosclerosis. This study was designed to elucidate the potential interaction between DNMT1 and miRNAs in the context of hyperhomocysteinemia (HHcy)-related atherosclerosis. In the aorta of ApoE-/- mice fed a high methionine diet, increased expression of miR-148a/152, with decreased DNMT1 mRNA and protein levels, was detected. Similar changes were observed in cultured foam cells stimulated with homocysteine. When miR-148a/152 was overexpressed using viral vectors, DNMT1 expression was suppressed, whereas the expression of adipose differentiation-related protein (ADRP) was enhanced, and the contents of total cholesterol (TC) and cholesteryl ester (CE) were increased in cultured foam cells. Conversely, downregulation of miR-148a/152 led to elevated DNMT1 expression, reduced ADRP expression, and lowered contents of TC and CE. The luciferase reporter assay verified that DNMT1 is a target gene for miR-148a/152 and overexpression of DNMT1 can partially reverse the miR-148a/152-induced lipid accumulation in foam cells. Meanwhile, we observed that DNMT1 overexpression enhanced DNA methylation and reduced miR-148a/152 expression. Our data showed reciprocal regulation between miR-148a/152 and DNMT1 in foam cells, which likely plays a critical role in HHcy-related atherosclerosis.
Introduction
A
miRNAs are a group of small noncoding RNAs that are ∼18 to 25 nucleotides in length, and they play a critical role in the regulation of gene expression (Miao et al., 2013; Lawless et al., 2014). Accumulating data suggest that miRNAs are extensively involved in a variety of physiological and disease processes, such as development and cancer (Piletic and Kunej, 2016). The role of miRNAs in cardiovascular diseases, including atherosclerosis (Kumar et al., 2014; Novak et al., 2014), has also been highlighted. The progression of atherosclerosis can be regulated by miRNAs through their effects on endothelial cells, vascular smooth muscle cells, and macrophages (Nazari-Jahantigh et al., 2015). The expression of miRNAs is modulated by different stimuli involved in every stage of atherosclerosis (Nazari-Jahantigh et al., 2015). Generally, the expression of one gene can be repressed by multiple miRNAs and one miRNA may repress multiple target genes, resulting in the formation of complex regulatory networks (Lopez-Bertoni et al., 2015). Numerous studies have demonstrated that DNMT1 maintains DNA methylation, which serves as a key mechanism controlling gene expression in atherosclerosis (Zaina, 2014; Yu et al., 2016). Solly et al. (2016) have found that miR-126* can target the DNMT1 3′-UTR, and enhance DNMT1 expression in high-risk myelodysplastic syndrome and low blast count acute myeloid leukemia, suggesting that miRNAs regulate DNMT1 expression by targeting the 3′-UTR. Such regulation happens in Hcy-induced vascular smooth muscle cell proliferation and colorectal cancer (Chen et al., 2015; Cao et al., 2016). Conversely, DNMTs may regulate miRNA expression by controlling gene methylation (Xue et al., 2015). It remains unclear whether and how miRNAs interact with DNMT1 in the pathogenesis of Hcy-related atherosclerosis. This study therefore aimed to explore this question. The data from this study will provide new insights into the molecular mechanism of atherosclerosis development, and also provide potentially promising strategies for its diagnosis and treatment.
Materials and Methods
Animals
The Institutional Ethics Committee of the Ningxia Medical University approved all procedures involving experimental animal use. The wild-type (ApoE+/+ ) and ApoE knockout (ApoE-/- ) C57BL6J mice were generously provided by the Animal Center of Peking University (Beijing, China). Six-week-old male mice (n = 48) were equally divided into four groups and fed the following diets (KeAoXieLi, Beijing, China) for 20 weeks: (1) control group, where normal C57BL/6J mice were fed a regular diet; (2) ApoE-/- + regular group, where ApoE-/- mice were fed a regular diet; (3) ApoE-/- + Met group, where ApoE-/- mice were fed a regular diet plus 1.7% methionine (Met); and (4) ApoE-/- + Met + F + V group, where ApoE-/- mice were fed a regular diet plus 1.7% methionine (w/w), 0.006% folate, and 0.0004% vitamin B12.
Induction, culture, and evaluation of foam cells
Foam cells were induced from human monocyte leukemia cell line THP-1 as described previously (Yang et al., 2015). Briefly, treatment of THP-1 cells with 100 nmol/L phorbol-12-myristate-13-acetate (PMA) (Promega, Madison, WI) for 24 h led to the differentiation of THP-1 cells into macrophages. Transformation of macrophages into foam cells was induced by incubation with 50 μg/mL ox-LDL (Yiyuan Biotechnologies, Guangzhou, China) for 24 h. The foam cells were treated as indicated for further experiments.
Tissue preparation and evaluation of atherosclerotic lesions in ApoE-/- mice
After blood collection, murine aortas were removed when mice were anesthetized followed by snap-freezing in liquid N2 and embedded in optimum cutting temperature compound. Frozen sections from the region of the proximal aorta starting from the end of the aortic sinus to 300 mm distally were taken according to Paigen et al. (1987). Sections were stained with Oil Red O and counterstained with hematoxylin and eosin. Quantitative analysis of lipid-stained lesions was performed on sections starting just beyond the end of the aortic sinus. The lipid-stained lesions were measured by digitizing morphometry and reported as mm2 per lesion. Besides, aortic root cross-sections were stained with MOMA-2 (Abcam, Cambridge, MA), a marker for monocytes (MC) and macrophages (Mφ) (Fang et al., 2014).
Determination of Hcy and lipid concentrations in the serum of ApoE-/- mice
We collected blood from fasted mice and separated the serum by centrifugation at 3000 g for 20 min. An automatic biochemistry analyzer (SIEMENS, Germany) was used for quantification of Hcy concentrations and lipid profiles, including high-density lipoprotein, triglyceride, total cholesterol (TC), and low-density lipoprotein, in the serum.
Bioinformatics analysis
Specific miRNAs that target DNMT1 were predicted using PicTar, miRanda, and TargetScan Targets algorithms.
Quantitative reverse transcription-PCR
The small RNA and total RNA from the mouse aorta and cultured foam cells were isolated using the RNA purification kit as recommended by the manufacturer (RNAiso for small RNA, TaKaRa, Dalian, China). First-strand cDNA was synthesized by reverse transcription (RT) using the Revert Aid first strand cDNA synthesis kit (MBI, Vilnius, Lithuania). The primers used for RT and quantitative RT-PCR (qRT-PCR) amplification of mature miR-148a/152 were purchased from RiboBio Technology (
DNMT1, DNA methyltransferase 1; F, forward; M, methylation primer; O, out primer; R, reverse; U, unmethylation primer.
Western blot analysis
Cells (tissues) were lysed in radio-immunoprecipitation assay (RIPA) buffer containing proteinase inhibitors. Western blot analysis was performed as we described previously (Yang et al., 2015). Antibodies against DNMT1 (Abcam) and β-actin (Boster, China) were used, and the blots were developed using the enhanced chemiluminescence reagent (Amersham Biosciences, Piscataway, NJ).
Construction of lentiviral vectors for miR-148a/152 mimics and inhibitors, and infection of foam cells
Lentivirus for miR-148a/152 mimics and inhibitors (dsRNA oligonucleotides) and negative control mimics were designed and constructed by GenePharma (GenePharma, Shanghai, China). Foam cells, induced from THP-1 cells as described above, were infected with lentivirus vectors. The cells stably expressing miR-148a/152 mimics or inhibitors were selected with puromycin for 3 to 4 week and then used for downstream experiments.
Luciferase reporter construct containing DNMT1 3′-UTR and validation of miR-148a/152 target sites
A luciferase reporter construct containing the 3′-UTR sequence of DNMT1 mRNA was made to examine whether DNMT1 3′-UTR is a target of miR-148a/152. The primers for amplification of wild-type and mutated 3′-UTR of DNMT1 mRNA were designed based on GenBank database (NM_007489.3). The sequence of the forward primer was 5′-CGGGA
pSicoR/miR-148a/152 and pMIR-Report/DNMT1 or pMIR-Report/Mut-DNMT1 were cotransfected into foam cells, and the specificity of miR-148a/152 targeting DNMT1 was examined by the relative activity of firefly luciferase unit in 48 h after transfection using a dual-luciferase reporter assay kit (Promega). To normalize the efficiency of each transfection, a Renilla luciferase expressing plasmid pRL-TK (Promega) was included in the transfection. All luciferase assays were repeated thrice.
Measurement of intracellular TC, cholesteryl ester, and free cholesterol content
To determine intracellular TC, cholesteryl ester (CE), and free cholesterol (FC) content, fully differentiated foam cells were seeded in 24-well plates. Cellular lipids were extract by 1 mL hexane:isopropanol (3:2) mixture after washing twice with phosphate-buffered saline (PBS). TC and FC contents were determined using a TC/FC assay kit (Infinity, Thermo Electron, Melbourne, Australia). Cells undergoing the same treatment were lysed in RIPA buffer to quantify the protein concentration and normalize data.
Immunofluorescence staining of adipose differentiation-related protein in foam cells
Cells in slides were fixed with 4% formaldehyde freshly prepared from paraformaldehyde in PBS for 20–30 min after washing with PBS. For immunofluorescence staining, cells were washed with PBS followed by permeabilization with 0.05% Triton X-100 and 10% goat serum in PBS for 5 min, and incubation with anti-adipose differentiation-related protein (ADRP) antibody (Abcam) overnight at 4°C. After washing with PBS, cells were incubated with a fluorescein isothiocyanate-conjugated donkey anti-rabbit IgG (Abcam) for 1 h at 37°C, followed by washing with PBS thrice, and the nuclei were then counterstained with DAPI (Bioss). The fluorescence signal was detected by a confocal microscope (Olympus Fluoview 1000, Japan).
Construction and transduction of DNMT1
The human DNMT1 gene was inserted into replication-defective adenoviral shuttle vector pHBAd-CMV-IRES-GFP and adenoviral backbone plasmid pBHGloxE1, 3Cre (Stratagene, La Jolla, CA) as previously described (Han et al., 2015). The DNMT1 fragment (Kpn1-Not1) from a DNMT1-pBSKS subclone (EcoR1-Not1) was inserted into pHBAd-CMV-IRES-GFP vector and transfected into BJ5183E coli by electroporation after being linearized with Pme1. Adenovirus packaging 293T cells were transfected with linearized recombinant plasmid pAd-DNMT1 to generate viruses. The recombinant adenoviruses were purified and their titers were determined as described (Yang et al., 2015). Foam cells were infected with adenovirus at the indicated multiplicity of infection for 24 h and treated with Hcy. DNMT1 expression was detected by Western blotting. DNMT1 cDNA carrying a wild-type 3′-UTR or a 3′-UTR containing mutated seed sequence for miR-148a/152 (DNMT1 mut) were cloned into pcDNA 3.1 for “rescue” experiments.
Nested methylation-specific PCR for miRNA-148a/152
Genomic DNA of foam cells was purified by the DNA extraction kit (Qiagen, CA, USA). EZ DNA Methylation-Gold kit was used for bisulfite conversion of genomic DNA (Zymo research, CA, USA) according to the manufacturer's instruction. The methylation status of miR-148a/152 was determined by methylation-specific PCR (MSP). PCR primers for methylated or unmethylated sequences are listed in Table 1. The PCR reaction using bisulphite-modified DNA as template was performed as follows: 95°C for 5 min followed by 40 cycles of 95°C for 1 min, 56°C for 1 min, and 72°C for 2 min, and then a final extension at 72°C for 7 min. PCR products were separated by a 2.0% agarose gel. The presence of methylation was calculated using the following formula: methylation% = methylation/(methylation + unmethylation) × 100%.
Statistical analysis
t-Test was used to compare results between two groups. One-way analysis of variance and additional analysis using the Student-Newman-Keuls test were used for multiple comparisons. p-Value <0.05 was considered statistically significant.
Results
High methionine diet results in elevated Hcy and lipids in serum and induces the formation of atherosclerotic lesions in ApoE-/- mice
To confirm the atherogenic effect of hyperhomocysteinemia (HHcy), atherosclerotic lesions were examined by cross-sectional analysis of the aorta root stained with hematoxylin-eosin and Oil Red O (Fig. 1A, B). Compared to the control group, the area of the atherosclerotic lesion in the three ApoE-/- groups (ApoE-/- + regular, ApoE-/- + Met, and ApoE-/- + Met + F + V) increased significantly by 8.18-, 13.58-, and 8.23-fold, respectively. However, the size of the plaque in the “ApoE-/- + Met + F + V” group diminished significantly compared with the “ApoE-/- + Met” group (Fig. 1D). In addition, increased MC/MØ infiltration was observed in atherosclerotic lesions of mice fed Met, as shown by the MC/MØ marker MOMA-2 (Fig. 1C). Determination of serum Hcy and lipid levels verified that high methionine diet caused HHcy (Fig. 1E) and hyperlipidemia (Fig. 1F). Taken together, these data suggest that a high methionine diet elevated serum homocysteine and lipids and promoted MC/MØ infiltration and the formation of atherosclerotic lesions in ApoE-/- mice.
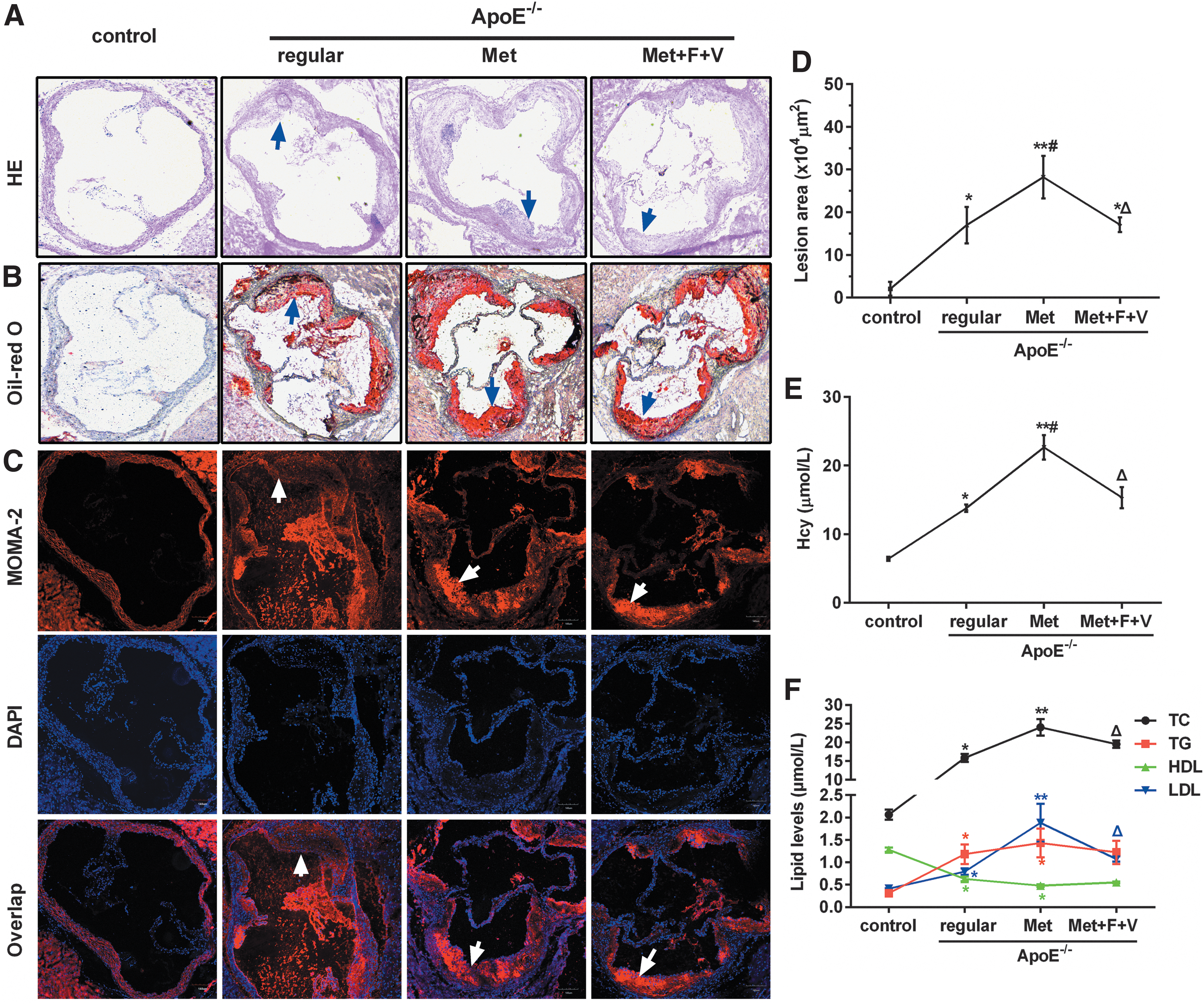
HHcy induced by Met increases atherosclerotic lesion size and accumulation of MC and MØ in ApoE-/-
mouse aortas. Normal C57BL/6J mice fed regular diet served as control. Six-week-old ApoE-/-
mice were randomly divided into three groups, fed either regular diet, or Met (1.7% w/w), or Met plus folate (0.006% w/w) plus vitamin B12 (0.0004% w/w) (Met + F + V), for 20 weeks. The cryostat sections (10 μm) of proximal aortas were stained with
Hcy suppresses the expression of DNMT1 mRNA and protein in ApoE-/- mice and cultured foam cells
Hcy, a thiol-containing amino acid formed during methionine metabolism, is involved in a one-carbon transfer reaction, which is important for DNA synthesis and methylation. DNMT catalyzes the transfer of methyl groups to DNA from S-adenosylmethionine. In mammals, DNMT1 is the maintenance methyltransferase, which preferentially methylates hemimethylated double-stranded DNA (Sharif et al., 2016). In this study, we examined the effect of Hcy on DNMT1 expression. Our results showed that DNMT1 mRNA and protein levels in the “ApoE-/- + Met” group greatly reduced compared with the control group (Fig. 2A, B). Similar results were observed for cultured foam cells (Fig. 2C, D). Such reduction of DNMT1 expression induced by Hcy was significantly attenuated by “folate plus vitamin B12,” an established treatment reducing Hcy levels, and by azacytidine (AZC), a DNA methylation inhibitor. These results suggest that DNMT1 played a crucial role in ApoE-/- mice fed Met and foam cells treated with Hcy.
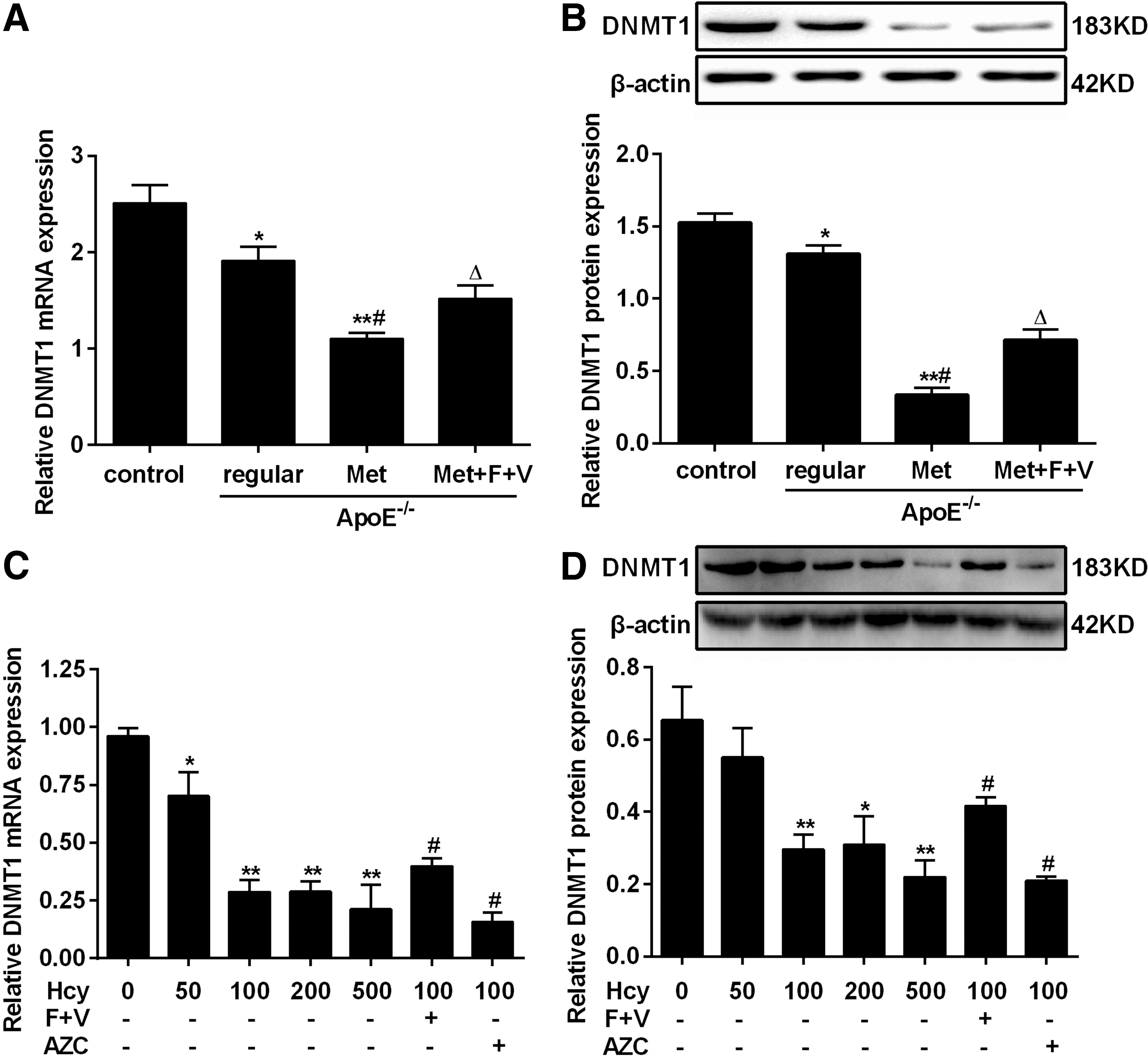
Homocysteine reduces DNMT1 expression in ApoE-/-
mouse aorta and foam cells.
The expression of miR-148a and miR-152 increases in high Met diet-fed ApoE-/- mice and Hcy-treated foam cells
miRNAs bind to imperfect sequence homology sites of mRNA and recruit the RNA-induced silencing complex, causing either degradation of mRNA or inhibition of protein translation (Novak et al., 2014; Nazari-Jahantigh et al., 2015). We hypothesized that miRNAs are involved in DNMT1 downregulation in the aorta of ApoE-/- mice and foam cells. To verify this hypothesis, we first analyzed the expression levels of miR-148a and miR-152 in the aorta of ApoE-/- mice. The results showed that miR-148a and miR-152 were dramatically upregulated in the “ApoE-/- + Met” group compared with the “ApoE-/- + regular” group (Fig. 3A, B). We further analyzed the expression levels of miR-148a and miR-152 in cultured foam cells treated with different concentrations of Hcy, either individually or with indicated treatments. Consistent with the results in ApoE-/- mice, the expression levels of miR-148a and miR-152 in foam cells increased significantly compared with the control (Fig. 3C, D). Demethylation treatment by AZC dramatically restored miR-148a/152 expression in foam cells (Fig. 3C, D), indicating that hypomethylation is responsible for the increase of miR-148a and miR-152 expression.
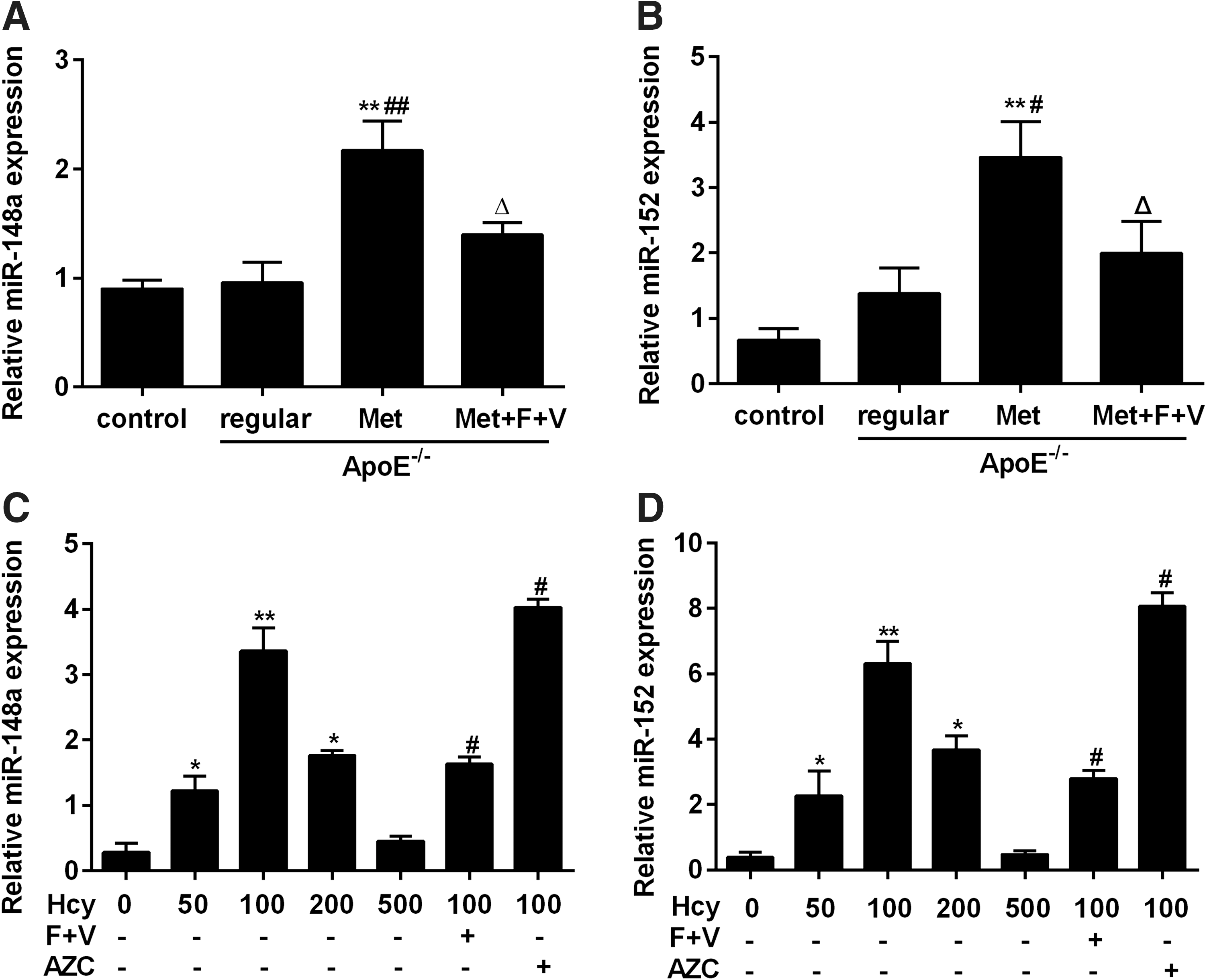
Homocysteine induces miR-148a/152 expression in ApoE-/-
mouse aorta and foam cells.
DNMT1 is a direct target of miR-148a/152
DNMT1 is critical for regulating DNA methylation in cardiovascular diseases, and is predicted as one of the high-scoring candidate genes of miR-148a/152 targets by four algorithms (TargetScan, PicTar, miRanda, and miRbase Target). As shown in Figure 4, DNMT1-encoded mRNA contained a 3′-UTR element ranging from dinucleotide 295–301 bp, which was stringently complementary to seed sequence of miR-148a/152, indicating that miR-148a/152 may directly target this site. To validate the predicted target, we cloned the wild-type 3′-UTR of DNMT1 with the seed sequence to the reporter plasmid. In parallel, we also constructed another reporter plasmid carrying a mutant DNMT1 3′-UTR, specifically mutated in seven point substitutions in the “seed region,” which did not target any gene, as predicted by the TargetScan, PicTar, miRanda, and miRbase Target (Fig. 4A). Transient transfection of the wild reporter plasmid into miR-148a/152-overexpressing foam cells resulted in a significantly reduced (p < 0.05) luciferase reporter gene activity compared with the control (miR-neg) (Fig. 4B). However, the activity of the mutant luciferase reporter gene was not reduced in miR-148a/152-overexpressing foam cells, indicating that the 3′-UTR of DNMT1 is a functional target site for miR-148a/152-induced silencing of DNMT1.
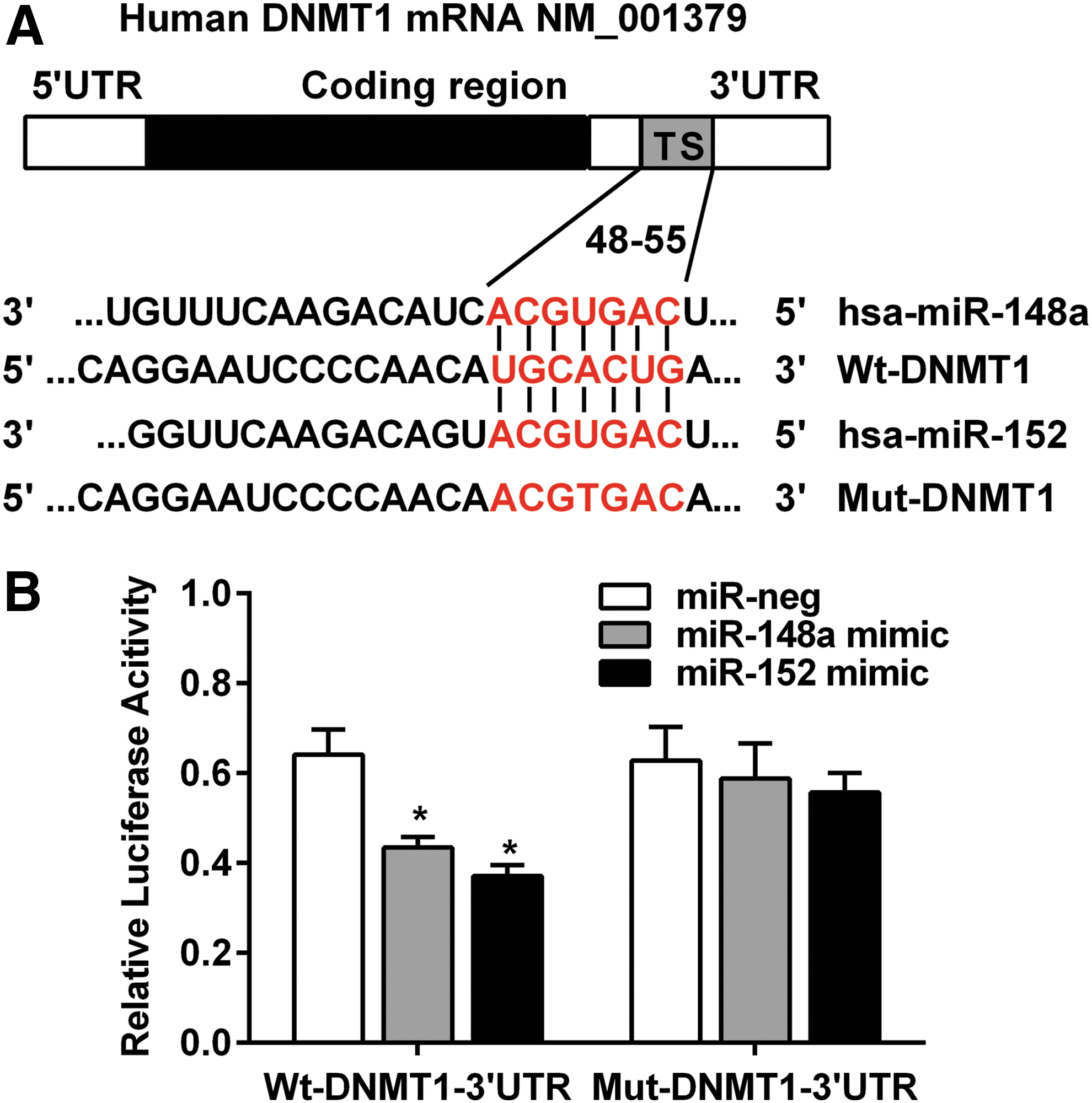
DNMT1 3′-UTR is a target of miR-148a/152.
miR-148a/152 inhibits DNMT1 expression in foam cells
To further validate the above results, we investigated whether miR-148a/152 affected the protein and/or mRNA levels of DNMT1, using foam cells derived from THP-1 cells receiving PMA and ox-LDL (Fig. 5). 100 μmol/L Hcy was added 48 h after the transfection of miR-148a/152 mimic or inhibitor vectors. The results showed that, either in the presence or absence of Hcy, DNMT1 expression reduced both at the mRNA and protein levels when miR-148a/152 expression was enforced. On the contrary, inhibition of miR-148a/152 increased DNMT1 expression. The data indicate that miR-148a/152 reversely regulates DNMT1 expression in foam cells.
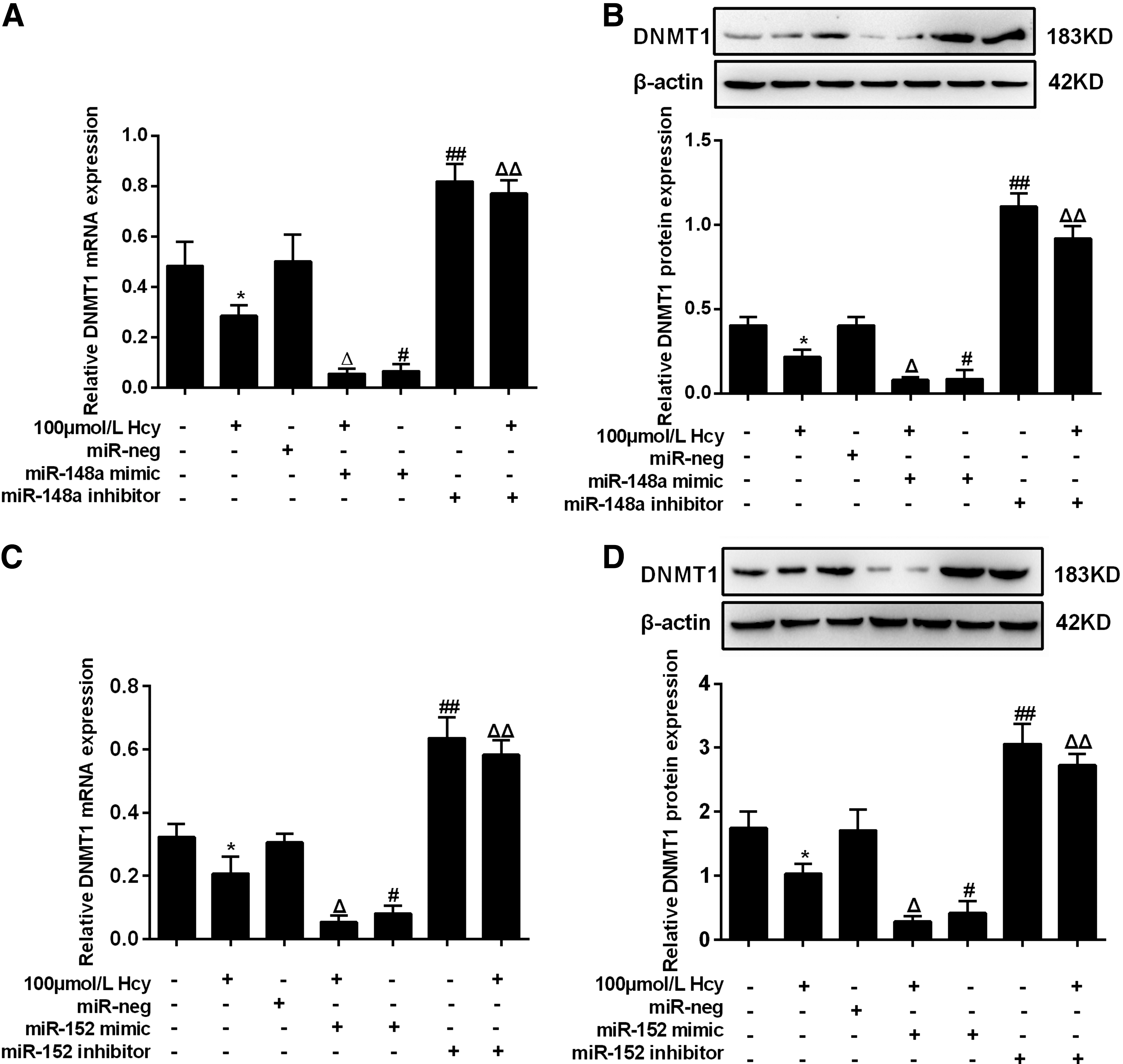
miR-148a/152 reversely regulates DNMT1 expression. Data are DNMT1 mRNA and protein levels in foam cells transfected with lentivirus vectors harboring miR-148a mimics or inhibitors
miR-148a/152 positively regulates the lipid content in foam cells
We further investigated the potential role of miR-148a/152 in lipid metabolism of foam cells. As shown in Figure 6A and B, overexpression of miR-148a or miR-152 mimics significantly increased the content of TC and CE induced by Hcy in foam cells, whereas it had no significant effect on the FC content. Consistently, inhibition of miR-148a or miR-152 reduced the content of Hcy-induced cellular lipids in foam cells. Meanwhile, Oil Red O staining showed lipid accumulation in foam cells (Fig. 6C). The results suggested that Hcy promoted lipid accumulation in foam cells.
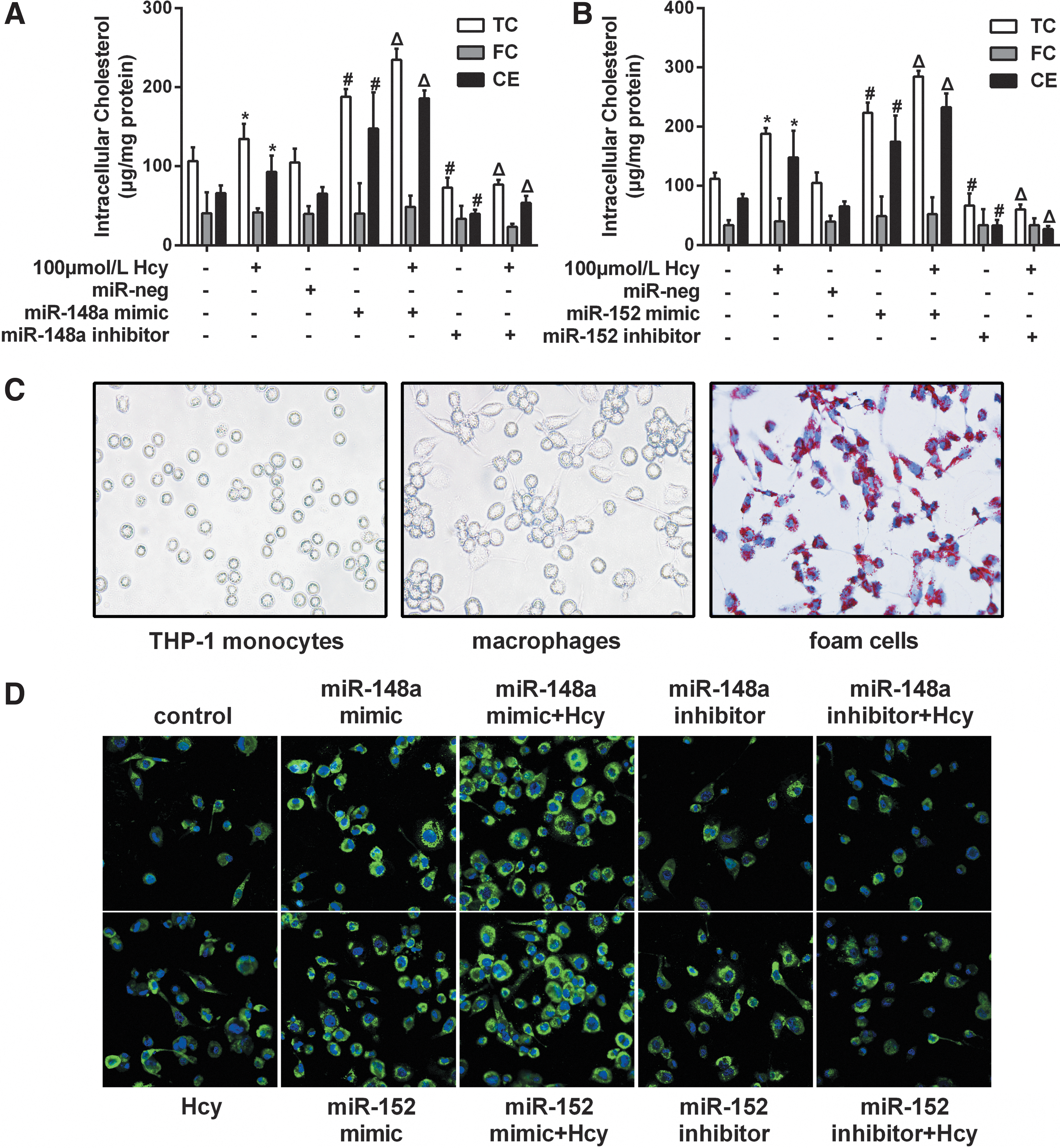
Analysis of lipid metabolism in foam cells transfected with mimics or inhibitors of miR-148a/152.
ADRP is a major structural protein of lipid droplets. Increases in ADRP is an early indication of adipocyte differentiation (Takahashi et al., 2016). We examined ADRP in foam cells, using the method of immunofluorescent staining. The results showed that overexpression of miR-148a or miR-152 significantly increased ADRP expression, whereas inhibition of miR-148a or miR-152 reduced ADRP expression (Fig. 6D). These data suggest that miR-148a/152 promoted the accumulation of lipid droplets in foam cells.
Overexpression of DNMT1 partially reversed the lipid accumulation in foam cells caused by miR-148a and miR-152
To test whether DNMT1 regulation contributed to the effect of miR-148a and miR-152 on lipid accumulation in foam cells after being treated with Hcy, we cotransfected miR-148a or miR-152 (or miR-neg) and a vector pcDNA3.1-DNMT1-mut containing DNMT1 mutated in the miR-148a and miR-152 binding site in the 3′-UTR into foam cells after being treated with Hcy, and found that DNMT1 is role of suppressing lipid accumulation was not limited by miR-148a and miR-152 when the sites in DNMT1 3′-UTR targeted by miR-148a and miR-152 were mutated. Besides, overexpressing DNMT1 with either pcDNA3.1-DNMT1 or pcDNA3.1-DNMT1-mut vector decreased lipid contents of foam cells after being treated with Hcy (Fig. 7A, B).
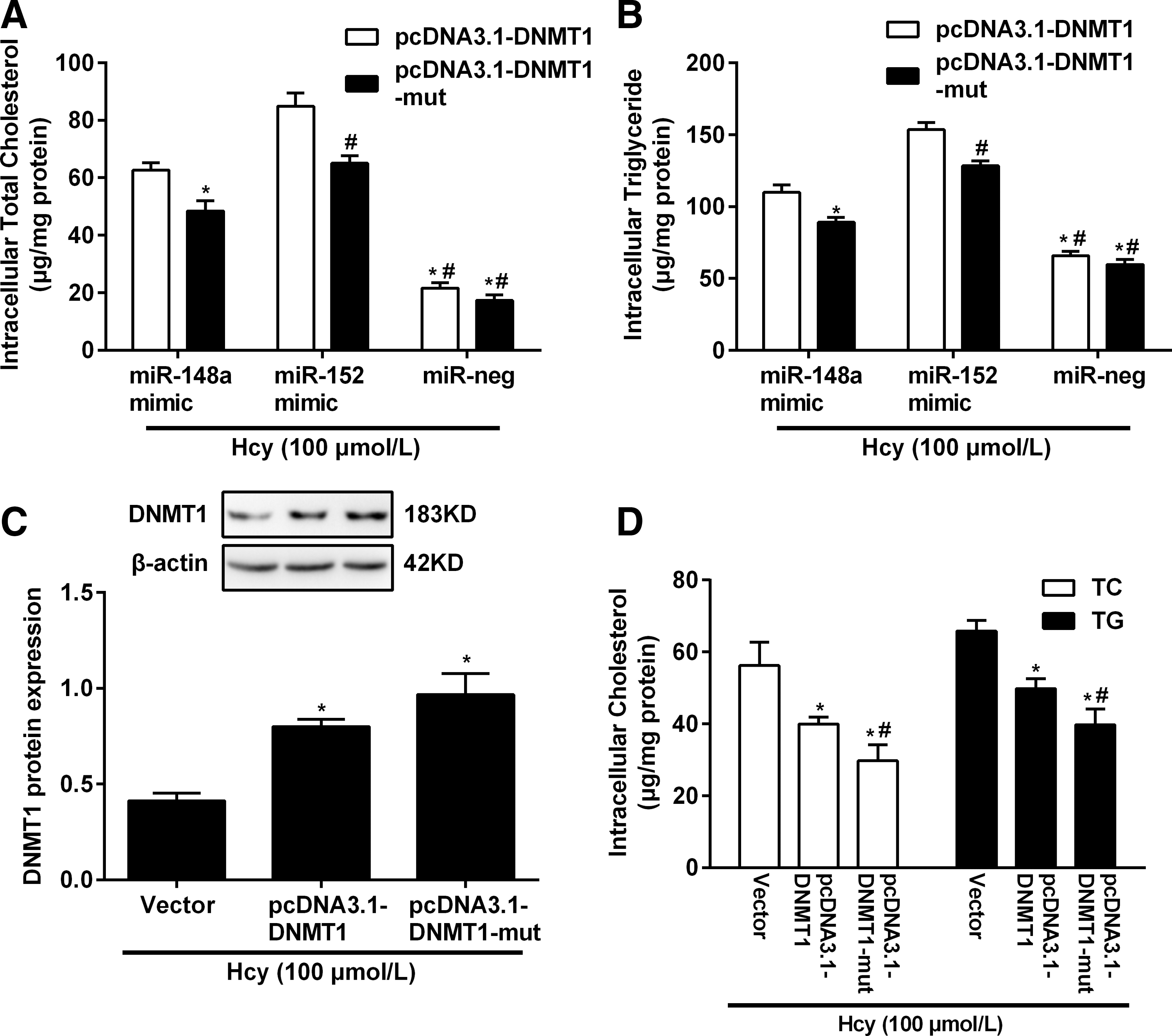
Expression of DNMT1 reversed TC and TG accumulation caused by miR-148a/152 in foam cells treated with Hcy. The TC
We then testified the function of DNMT1-expressing vectors that included a 3′-UTR mutated gene (pcDNA3.1-DNMT1-mut) in foam cells treated with Hcy. Western blot analysis showed that the level of the DNMT1 protein expressed from the construct mutating the DNMT1 3′-UTR was the highest compared to the cells transfected with pcDNA3.1-DNMT1 containing the full-length DNMT1 cDNA or the negative control (Vector) (Fig. 7C). Lipid accumulation assay in foam cells after being treated with Hcy showed that overexpression of DNMT1 reversed the effect of miR-148a and miR-152 on lipid accumulation by at least 28% (Fig. 7D). However, in the foam cells after being treated with Hcy, which express high levels of endogenous miR-148a and miR-152, overexpression of DNMT1 protein by the pcDNA3.1-DNMT1 was decreased compared to the amount expressed by pcDNA3.1-DNMT1-mut, which carries the DNMT1 3′-UTR mutation, and its effect on suppressing lipid accumulation was also less strong. Since miR-148a and miR-152 is expressed endogenously in foam cells, it is likely that endogenous miR-148a and miR-152 inhibit DNMT1 overexpression by targeting its 3′-UTR.
DNMT1 downregulation leads to hypomethylation of miR-148a and miR-152 gene promoters
Gene promoter methylation often occurs in cardiovascular disease, resulting in the gene expression inhibition of the methylated gene (Muka et al., 2016). We tested whether Hcy treatment-induced DNA methylation leads to upregulation of miR-148a/152 expression in foam cells. To verify this hypothesis, we analyzed the sequence of genomic DNA spanning of miR-148a and miR-152 genes and found large amount of CpG-rich regions (CpG islands) in the promoter regions of both genes. Subsequently, MSP was run to detect the methylation status of the promoter regions of miR-148a and miR-152. We found hypomethylation of CpG islands in miR-148a and miR-152 promoters in Hcy-treated foam cells. Furthermore, the methylation can be inhibited by DNA methylation inhibitor AZC (Fig. 8A, B). To further know the role of DNMT1 in the regulation of miR-148a and miR-152 expression, adenovirus vectors of DNMT1 were constructed and infected into the foam cells to overexpress de novo DNMT1 (Fig. 8C, D). The results revealed that DNMT1 upregulation increased the methylation of miR-148a and miR-152 genes (Fig. 8E, F) and downregulates miR-148a/152 expression (Fig. 8G, H). Collectively, DNMT1 overexpression results in the hypermethylation of miR-148a and miR-152 genes, leading to the downregulation of miR-148a and miR-152 levels.

DNMT1 overexpression enhances miR-148a/152 promotor methylation and reduces their expression.
Discussion
Tremendous progress has been made in recent years in understanding the biology of atherosclerosis, which aids in the prevention and treatment of this condition (Rustempasic et al., 2014). Hcy is a thiol-containing nonprotein amino acid that has been recognized as an independent predictor of atherosclerosis (McCully, 2015). In addition, the molecular mechanism underlying the significance of elevated Hcy as an independent risk factor for atherosclerosis can be explained by its direct effect on DNA methylation (Zhang et al., 2013; Eren et al., 2014).
DNA methylation is an epigenetic change that results in the addition of a methyl group at carbon-5 of cytosine residues. This process is mediated by DNMTs, a family of enzymes that includes DNMT1, DNMT3a, and DNMT3b (Li et al., 2014). DNMTs are ubiquitously expressed in normal human tissues, but overexpressed in various tumors, including leukemia, colorectal, ovarian, and prostate cancers (Berg et al., 2004; Sui et al., 2013; Flis et al., 2014). DNMT1, the principal DNA methyltransferase in mammalian cells, is a large and highly dynamic enzyme with multiple regulatory features that control DNA methylation (Shaffer et al., 2015). We previously reported (Yang et al., 2015) that DNMT1 functioned as a biomarker of gene methylation status in HHcy-associated atherosclerosis. However, the regulation mechanism on DNMT1 remains elusive.
MiRNAs have been shown to target the mRNA through 5′-seed sequences interacting with miRNA regulatory elements mostly located in the 3′-UTR of the target mRNA (Pi et al., 2015). Our bioinformation analyses showed that miR-148a/152 potentially modulates DNMT1 expression. Previous literature reported the inhibition of DNMT1 expression by miR-148a/152 in tumor cells (Xu et al., 2013). In accordance, this study observed that miR-148a/152 reversely regulates DNMT1 gene expression in the context of Hcy-accelerated atherosclerosis, and ectopic miR-148a/152 expression promoted lipid accumulation in foam cells by silencing DNMT1. Overexpression of DNMT1 without 3′-UTR targeting by miR-148a/152 led to decreased lipid accumulation in foam cells treated with Hcy. Besides, we also observed that DNMT1 controls methylation of miR-148a/152 gene promoter region.
Previous studies have shown that DNMT1 overexpression contributes to gene promoter hypermethylation and is associated with the malignant potential and poor prognosis of human cancer (Lee et al., 2016). DNA methylation is maintained by DNMT1 and serves as a key mechanism that controls gene expression in a variety of chronic diseases such as atherosclerosis. It has been suggested that DNA methylation can silence or activate gene expression by hypermethylation or hypomethylation, respectively (Li et al., 2015). Recent studies (Valastyan, 2012) revealed that miRNAs play pivotal roles in normal physiology and disease, and investigation of miRNA expression for therapeutic targeting is currently underway. miRNA upregulation is mainly achieved by administration of synthetic miRNAs or miRNA-expressing vectors; whereas, miRNA is downregulated upon administration of antisense nucleotides, often chemically modified to ensure stability and specificity. There are multiple potential limitations associated with the development and testing of miRNA-based therapeutics. Like protein-coding genes, DNA sequences encoding miRNAs may undergo aberrant DNA methylation, leading to miRNA upregulation (through DNA hypomethylation) or downregulation (through DNA hypermethylation) in human cancers. We upregulated DNMT1 expression in foam cells by employing adenovirus vectors and observed the enhancement of miR-148a/152 gene promotor methylation, with reduced levels of miR-148a/152. This result is in line with documented role of DNMT1 (Yu et al., 2016).
Conclusions
In summary, this study has defined the reciprocal regulation between miR-148a/152 and DNMT1 in Hcy-accelerated foam cell differentiation and atherosclerotic lesion. It offers novel insights for understanding the mechanism underlying aberrant methylation and gene expression in Hcy-accelerated atherosclerosis, and potentially provides novel strategies for intervening atherosclerosis.
Footnotes
Acknowledgments
This study was funded by the National Natural Science Foundation of China (Grant Nos. 81260105, 81360027, 81360052, 81360053) and Natural Science Foundation of Ningxia Province (Grant No. NZ16207).
Disclosure Statement
No competing financial interests exist.