
Abstract
As one of transcription factors and pluripotency factors, Klf4 plays a crucial role in regulation of cellular processes. In this article, we characterized Klf4 of large yellow croaker (Lc-Klf4), which encodes a 452-amino acid protein (Lc-Klf4) with three highly conserved C2H2 zinc fingers. Lc-Klf4 shares high conservative functions in teleosts with the closest relationship with Stegastes partitus. The spatiotemporal expression showed that Lc-Klf4 was expressed widely in adult tissues with gender difference as follows: brain>gill>eye>heart in female; heart>testis>gill>brain in male; male>female in heart, gill, and testis; and female>male in eye. During growth, the highest expression level of Lc-Klf4 was at 635 dph (days posthatching) in testis and at 270 dph and 635 dph in brain. Besides, Lc-Klf4 was widely and highly distributed in different developmental spermatids especially in spermatocytes. The expression of Lc-Klf4 in embryos was exhibited zygotically beginning from late gastrula stage with high level in closure of blastopore stage and appearance of optic vesicle stage. This expression pattern was supported by whole-mount in situ hybridization with high expression in back and head of late embryos. In conclusion, the spatiotemporal expression patterns of Lc-Klf4 illustrated that Klf4 involves in spermatogenesis, embryogenesis, and adult physiological processes.
Introduction
A
In general, the generation of iPSCs is performed by in vitro transfection of pluripotency factors (e.g., Oct4, Sox2, Klf4, cMyc, Nanog, and Lin28) to cause somatic cell reprogramming (Takahashi and Yamanaka, 2006; Cox and Rizzino, 2010). The pluripotency factors function in establishing and maintaining the pluripotent state of iPSCs by expression regulation of multiple genes (Rao et al., 2010). An ancient trait of reprogramming to pluripotency has been confirmed in vertebrate Oct4 and Pou2 proteins such as mouse (Mus musculus), human (Homo sapiens), axolotl (Ambystoma mexicanum), and medaka (Oryzias latipes) (Tapia et al., 2012). Similarly, reprogramming capacity of Oct4 and Nanog is functionally conserved in mammalians, avians, amphibians, and teleosts (Sánchez-Sánchez et al., 2011; Theunissen et al., 2011). So far, iPSCs have been successfully obtained by various methods from a variety of mammalian somatic cells (Debowski et al., 2015; Rohani et al., 2016), such as rodent and human fibroblasts (Ruggieri et al., 2014), human cord blood CD34+ cells (Meng et al., 2012), human neural progenitor cells (Tsai et al., 2015), and porcine mesenchymal cells (West et al., 2010). Moreover, some iPSCs also have been induced to differentiate into specific cells and successfully applied to clinical therapy (Grskovic et al., 2011; Gao et al., 2015; Ruggieri et al., 2014). Otherwise, other animal models have been introduced in nonmammalian species, such as chicken (Gallus gallus), Xenopus (Xenopus laevis), zebrafish (Danio rerio), and medaka (Christen et al., 2010; Hong et al., 2011; Sánchez-Sánchez et al., 2011). Therefore, iPSCs technology should be feasible in the large yellow croaker.
iPSCs technique circumvents ethical legislations on human embryonic stem cells (ESCs) and is widely applied in basic research, regenerative medicine, disease prevention, gene therapy, drug screening, and disease modeling. For example, vaccination projects in oncogenesis, infectious disease, and allergy prevention (Brouwer et al., 2016); cell replacement therapies for incurable diseases associated with degeneration of a specific cell type or a tissue by inducing pluripotent stem cells to differentiate into the desired cell type (Holditch et al., 2014). Likewise, the application of the iPSCs technique in fish emerges, such as teleost models of hematopoietic stem cells in zebrafish and clonal ginbuna carp (Carassius auratus langsdorfii) (Kobayashi et al., 2016) and fish spermatogonial stem cell for research on basic and applied reproductive biology (Panda et al., 2011; Lacerda et al., 2014).
Klf4 is one of the important pluripotency factors as well as a member of Kruppel-like factor (Klf) family, which includes Klf1 to Klf17 in mammals (Bieker, 2001). Klfs generally contain three Cys2/His2 (C2H2) of zinc fingers, which are the conserved DNA binding domains at the carboxyl terminus (C-terminus) (Turner and Crossley, 1999; Dang et al., 2000). The amino terminal (N-terminal) regions are diverse in different Klfs, which are involved in transcription regulation as activators, repressors, or both (Rowland and Peeper, 2006; McConnell and Yang, 2010; Moore et al., 2011). Klfs have been proved to regulate diverse biological processes, such as cell growth, differentiation, embryogenesis, and tumorigenesis (Dang et al., 2000; Yamazaki et al., 2008; Lin et al., 2010; Kotkamp et al., 2014; Gao et al., 2015), and different Klfs function to the differentiation of different cell types (Carlson et al., 2006; Kajimura et al., 2007; Swamynathan et al., 2007; Alder et al., 2008). Aberrant expression of Klfs can result in various diseases (Cao et al., 2010; McConnell and Yang, 2010; Tetreault et al., 2013).
Klf4 was identified first as a downstream target of LIF/STAT3 signaling in ESCs of mice with characteristic three C2H2 zinc fingers (Li et al., 2005). Klf4 plays a vital role in mammalian embryonic and organ development (Chiambaretta et al., 2004; Swamynathan et al., 2007; Moore et al., 2011; Aksoy et al., 2014; Apara and Goldberg, 2014; Fang et al., 2016). As mammalian Klf4 orthologs, teleostean Klf4 also participates in embryogenesis and organogenesis. For example, zebrafish Klf4 involves in anterior mesendoderm/prepolster differentiation, hatching, and embryonic erythropoiesis (Gardiner et al., 2005, 2007). Zebrafish Klf4a is essential for differentiation of goblet cells and enterocytes, as well as repression of intestinal cell proliferation (Li et al., 2011). Besides, zebrafish Klf4 also involves in optic nerve regeneration (Nishitani et al., 2010). Similar to zebrafish Klf4, the maternal inheritance and early embryonic expression are detected in medaka Klf4, which perhaps regulates medaka ESCs culture (Wang et al., 2011).
By far, no data of Klf4 in large yellow croaker were found, hence we reported the structural characteristic, evolutionary conservation, and temporal–spatial expression of Lc-Klf4 in various tissues and development stages of embryos and gonads. Our data will contribute to further research on Lc-Klf4 function and provide tools for efficient generation of iPSCs in large yellow croaker.
Materials and Methods
Sample collection
The samples of adult tissues and embryos were obtained from healthy large yellow croakers cultured in the Fishery Technology Extension Station of Ningde, Fujian, China. Eleven types of adult tissues (gonad, head kidney, muscle, liver, spleen, brain, gill, stomach, eye, intestine, and heart) were dissected from both sexes (female 508 ± 88 g, male 423 ± 72 g, at least 16, respectively) and treated by being stored promptly in RNAlater overnight at 4°C, then kept at −20°C for quantitative real-time PCR (qRT-PCR). In addition, different developmental gonads (juvenile fish: 270 days posthatching (dph), female 55 ± 15 g, male 57 ± 16 g; subadult fish: 635 dph, female 311 ± 53 g, male 289 ± 63 g; and adult fish: 1000 dph, female 445 ± 67 g, male 399 ± 69 g) were dissected and treated in two ways: one way for qRT-PCR as described above; another way for in situ hybridization by being fixed in 4% paraformaldehyde (PFA)–phosphate-buffered saline (PBS) fixation buffer overnight at 4°C, then rinsed with 1× PBS, dehydrated in gradient dilute methanol (70%, 80%, 90%, and 100% methanol), and kept at −20°C finally.
Embryos (2-cell, 4-cell, 8-cell, 16-cell, 32-cell, 64-cell, multiple-cell, blastula, gastrula, formation of yolk plug, appearance of optic vesicle, closure of blastopore, formation of eye lens, tail bud, beginning of heart pulsation, prehatching, and 1-day posthatching stage) were gathered according to our previous description (Jiang et al., 2015). The treatment of embryos was similar to gonads: a part for qRT-PCR, another part for whole-mount in situ hybridization (WISH). Inconsistently, for WISH, embryonic chorions must be first removed manually before being fixed in 4% PFA.
Total RNA extraction and first-strand cDNA synthesis
Total RNAs were extracted from tissues (four individuals as a sample) and embryos (about 30 embryos as a sample) of large yellow croaker using TRIzol reagent (Invitrogen, Carlsbad, CA) with quality and purity detection of RNAs by spectrophotometer (Nanodrop 2000; Thermo Scientific) and 1.0% agarose gel electrophoresis. Next, the high-quality RNAs were treated by DNase I (Promega, Shanghai, China) to remove the genomic DNA. Then, 3 μg of RNAs of each tissue and embryo was used to synthesize first-strand cDNA as qRT-PCR template by reverse transcription-PCR (RT-PCR) with random hexamers and the M-MLV reverse transcriptase (Promega). In the end, the synthesized first-strand cDNAs were diluted and stored at −20°C.
cDNA cloning of Lc-Klf4
The gene-specific primers for full-length cDNA of Lc-Klf4 were designed with Primer 5.0 based on Lc-Klf4 mRNA sequences from GenBank database (accession number XM_010742687.1) (Table 1). The cDNAs of brain were used as the templates for head to toe PCR. The PCR products were gel-purified, inserted into the pMD19-T vector (Takara, Dalian, China), and transformed into DH5α-competent cells. The positive clones were sequenced.
F, forward; R, reverse; qRT-PCR, quantitative real-time polymerase chain reaction.
Bioinformatic analysis of sequence
Nucleotide sequence homology of Lc-Klf4 was analyzed by BLAST software (
Quantitative real-time polymerase chain reaction
According to the existing sequence of Lc-Klf4 in NCBI (XP_010740989.1), the gene-specific primers of Lc-Klf4 for qRT-PCR were designed by Primer 3.0 software (Table 1). The primers of reference gene β-actin (as reference gene of tissue expression) and 18S rRNA (as reference gene of embryo expression) were designed in reference to Jiang et al. (2015) (Table 1). Each sample (tissues and embryos) for qRT-PCR was properly diluted as templates and assayed in quadruplication. qRT-PCR was performed with the SYBR Green PCRMaster Mix (Applied Biosystem) and analyzed by the melting curve in LightCycler 480 system and electrophoretogram. The data were analyzed by the comparative threshold cycle method and expressed as the mean of RQ value (2−ΔΔCT) (ΔCT = CT of target gene minus CT of β-actin (or 18S rRNA), ΔΔCT = ΔCT of any sample minus calibrator sample) with arithmetic mean ± standard error of the mean. The results were analyzed with SPSS version 20.0 (SPSS, Inc., Chicago, IL) for one-way analysis of variance and independent samples T test (T-test). The statistically significant differences were shown at p < 0.05 and most significant differences were shown at p < 0.01.
In situ hybridization
Probe preparation
The digoxigenin (DIG)-labeled RNA probes needed to be prepared before in situ hybridization. The preparation process of probes is as follows: First, an Lc-Klf4 cDNA fragment was amplified by gene-specific primers for in situ hybridization using testis cDNA as the templates (Table 1). Next, the PCR product was gel-purified, ligated into PGEM-T Easy Vector (Promega, Madison), transformed into DH5α-competent cells, and identified by monoclonal colony PCR using generic primer T7 as a forward primer and target gene primers as reverse primers to confirm the insert direction of target gene. Then, the Lc-Klf4 cDNA-harboring vector was linearized and used for in vitro transcription by T7 RNA polymerase (Thermo scientific, Lithuania) and 10 × DIG-RNA Labeling Mix (Roche, Indianapolis) with T7 reverse primer of target gene for sense and T7 forward primer of target gene for antisense DIG-labeled cRNA probes as described by Chen et al. (2015). Finally, the DIG-labeled probes were stored at −80°C.
Tissue in situ hybridization
The protocol of in situ hybridization in tissues was carried out as our previous description (Jiang et al., 2015). The main processes included six steps: (1) the preparation of paraffin section: dehydration, transparentizing, paraffin embedding, paraffin section, and drying at 42°C; (2) the treatment before hybridization: dewaxing, rehydrating, PBS washing, and proteinase K (1.0 mg/mL) digestion at 37°C; (3) hybridization: prehybridization in prehybridization buffer (Hyb−, no DIG-labeled RNA probe) at 65°C for 3 h, hybridization in hybridization buffer (Hyb+, DIG-labeled RNA probe, 1.0 ng/μL) at 65°C for 16 h on a rotary shaker; (4) the treatment after hybridization: washing by Hyb− gradient, sealing by PBST-SB, and incubating with anti-DIG-AP antibody (1:5000) overnight at 4°C; (5) dyeing: washing by PBST and TMNT, staining with BCIP/NBT in the dark; and (6) observation: observation and photomicrograph by optical microscope.
Whole-mount in situ hybridization
Unlike tissue in situ hybridization, WISH was started directly from embryo rehydrating without the sectioning step. The next steps were similar to tissue in situ hybridization except the concentration of proteinase K with 10 mg/mL. In addition, observation and photomicrograph were performed by stereo microscope.
Results
Identification and characterization of Lc-Klf4
The full-length cDNA of Lc-Klf4 was confirmed by head to toe PCR. The nucleotide sequence and deduced amino acid sequence of Lc-Klf4 are displayed in Figure 1. The full length of Lc-Klf4 cDNA is 2404 bp, which contains an open reading frame encoding a 452-amino acid (aa) protein Lc-Klf4, a 255 bp 5′ untranslated region (UTR), and a 790 bp 3′UTR. The putative polyadenylation signal (AATAAA) was found at 455 bp downstream of the termination codon TGA (Fig. 1).

Full-length cDNA of nucleotide sequence and deduced amino acid sequence of Lc-Klf4 from large yellow croaker. The initiation codon (ATG) and the stop codon (TGA) are indicated in bold. The putative polyadenylation signal (AATAAA) and DNA CACCC box binding motifs (GGGTG) are marked in box. There ZnF-C2H2 domains are indicated as underline. The Glu-rich region and Ser-rich region are displayed by wavy line. The potential nuclear localization signal (KPKRGRR) is marked by dotted line. The glycosylation sites are marked in gray. The phosphorylation sites are flagged as follows: serine by circle, threonine by rectangle, and tyrosine by triangle. Color images available online at
Bioinformatic analysis showed that Lc-Klf4 is located in the nucleus (k-NN prediction, k = 23) with a potential nuclear localization signal (KPKRGRR). The estimated MW of Lc-Klf4 is 49.91 kDa, theoretical IP is 8.64, and grand average of hydropathicity is −0.846. The abundant Serine (Ser) (14.8%), proline (Pro) (9.1%), and glutamine (Gln) (8.4%) were predicted in amino acid composition of Lc-Klf4. The N-terminal signal peptide cleavage site and transmembrane helix are absent in Lc-Klf4. Forty-four phosphorylation sites (Ser: 33, Thr: 5, Tyr: 6) and three glycosylation sites were predicted in Lc-Klf4. The structural and functional domain analysis exhibited that Lc-Klf4 possesses three ZnF-C2H2 domains, which are the DNA binding domains by motifs (GGGTG) binding to DNA, and located at aa: 369–393, 399–423, and 429–451 of Lc-Klf4, respectively (Fig. 1). Moreover, a Glu-rich region (aa: 98–115) and Ser-rich region (aa: 285–289) were predicted in Lc-Klf4 (Fig. 1). Additionally, three α-helices were deduced in the tertiary structure of Lc-Klf4 (aa: 363–450) by the model based on template of Xenopus tfiiia zinc finger domain 2 (PDB code: c1tf6D) (Fig. 2).
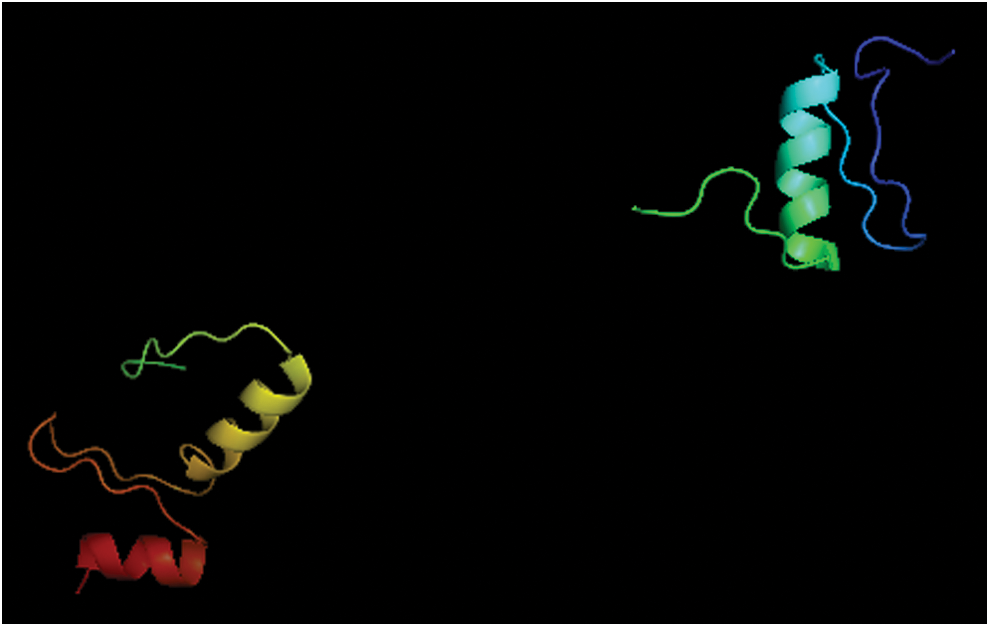
The predicted tertiary structure of Lc-Klf4 (ZnF-C2H2 domains) in large yellow croaker containing three α-helices. Color images available online at
Phylogenetic analysis of Lc-Klf4
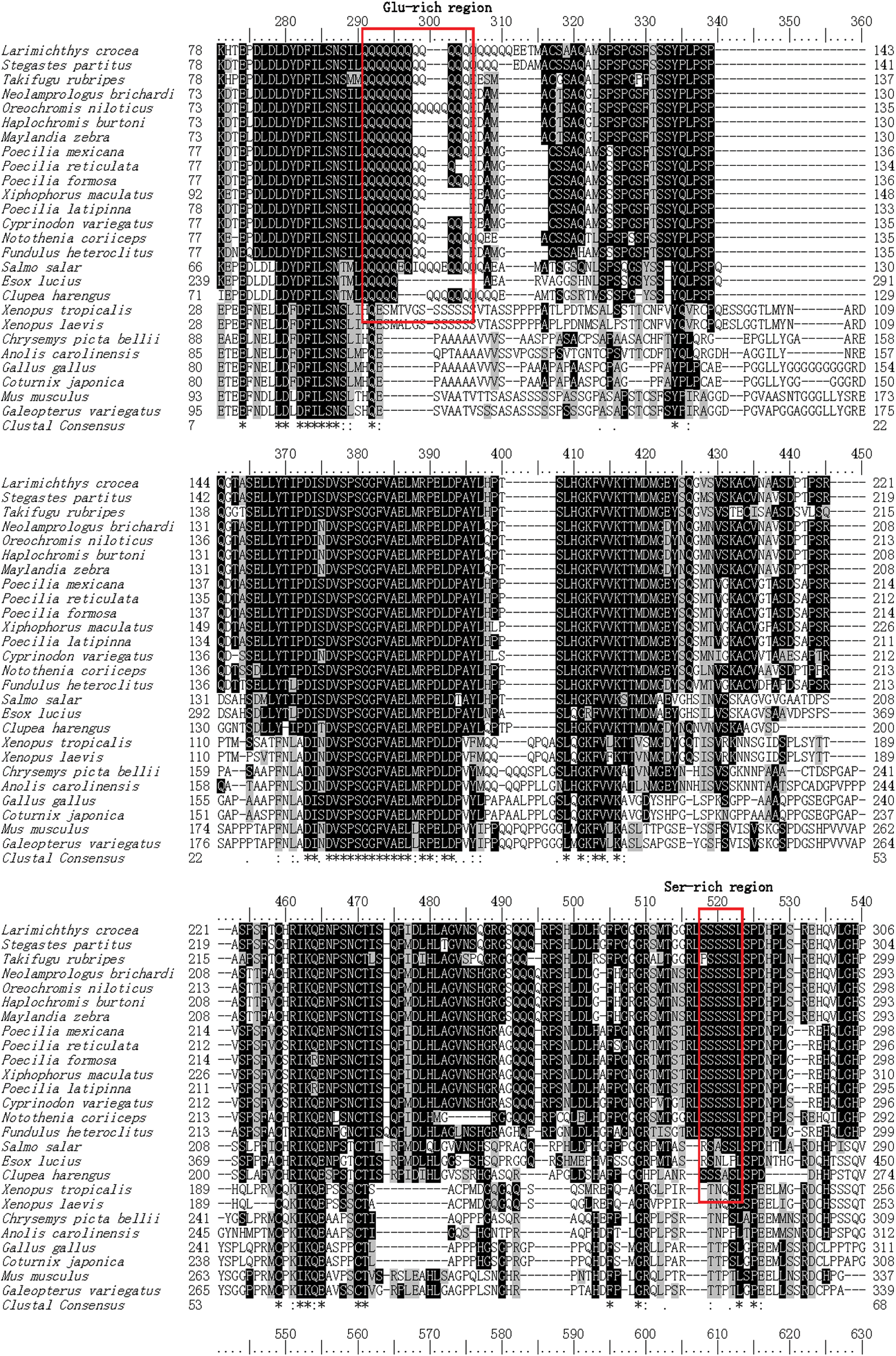
The homology analysis of Klf4 by multiple sequence alignment was performed between large yellow croaker and other known vertebrates. Lc-Klf4 shared the high identity with teleosts (68–93%) and low with amphibians, reptiles, birds, and mammals (under 56%) (Table 2, Fig. 3). In teleosts, the particularly high identity with Lc-Klf4 was shown as follows: Stegastes partitus (94%), Oreochromis niloticus (90%), Haplochromis burtoni (90%), Notothenia coriiceps (89%), Maylandia zebra (89%), Takifugu rubripes (89%), and Poecilia reticulata (86%). Interestingly, the multiple sequence alignment demonstrated that three ZnF-C2H2 domains of Klf4 maintained highly conservative levels in all vertebrates (above 95%); however, Ser-rich region and Glu-rich region were conserved only in teleosts (Table 2, Fig. 3).

Protein multiple alignment of Lc-Klf4 between large yellow croaker and other known vertebrates. The GenBank accession numbers of the selected Klf4 proteins are displayed as follows: Larimichthys crocea (XP_010740989.1), Stegastes partitus (XP_008283778.1), Takifugu rubripes (XP_011602852.1), Neolamprologus brichardi (XP_006789081.1), Oreochromis niloticus (XP_003455020.1), Haplochromis burtoni (XP_005931263.1), Maylandia zebra (XP_004549659.1), Poecilia mexicana (XP_014824425.1), Poecilia reticulate (XP_008422660.1), Poecilia formosa (XP_007559021.1), Xiphophorus maculatus (XP_005812733.2), Poecilia latipinna (XP_014883061.1), Cyprinodon variegatus (XP_015254367.1), Notothenia coriiceps (XP_010764282.1), Fundulus heteroclitus (XP_012724460.1), Salmo salar (XP_013980715.1), Esox lucius (XP_010891124.1), Clupea harengus (XP_012689201.1), Xenopus tropicalis (NP_001017280.1), Xenopus laevis (AEK12773.1), Chrysemys picta bellii (XP_005283537.1), Anolis carolinensis (XP_008113506.1), Gallus gallus (XP_004949426.1), Coturnix japonica (XP_015705130.1), Mus musculus (NP_034767.2), and Galeopterus variegatus (XP_008581926.1). ZnF-C2H2 domains are marked by underline. Ser-rich region and Glu-rich region are indicated by box. Color images available online at

The phylogenetic tree was constructed by the neighbor-joining method of MEGA 5.0 (Fig. 4) to clarify the evolutional relationship of Klf4 in vertebrates. The results showed that two Klf4 groups were clustered in vertebrates: teleost group and tetrapod group (containing amphibians, reptiles, birds, and mammals). Three subgroups were divided in the teleost group, which were clustered by Perciformes, Cyprinodontiformes, and other teleosts, respectively. Thereinto, Perciformes subgroup was clustered first with Cyprinodontiformes subgroup, then with other teleost subgroup. Furthermore, Perciformes subgroup was divided into two smaller branches. Obviously, Lc-Klf4 belonged to one of Perciformes subgroup branch with being clustered first with S. partitus, next with N. coriiceps. Notably, T. rubripes was clustered in Perciformes subgroup and shared the close relationship with Lc-Klf4 (Fig. 4).
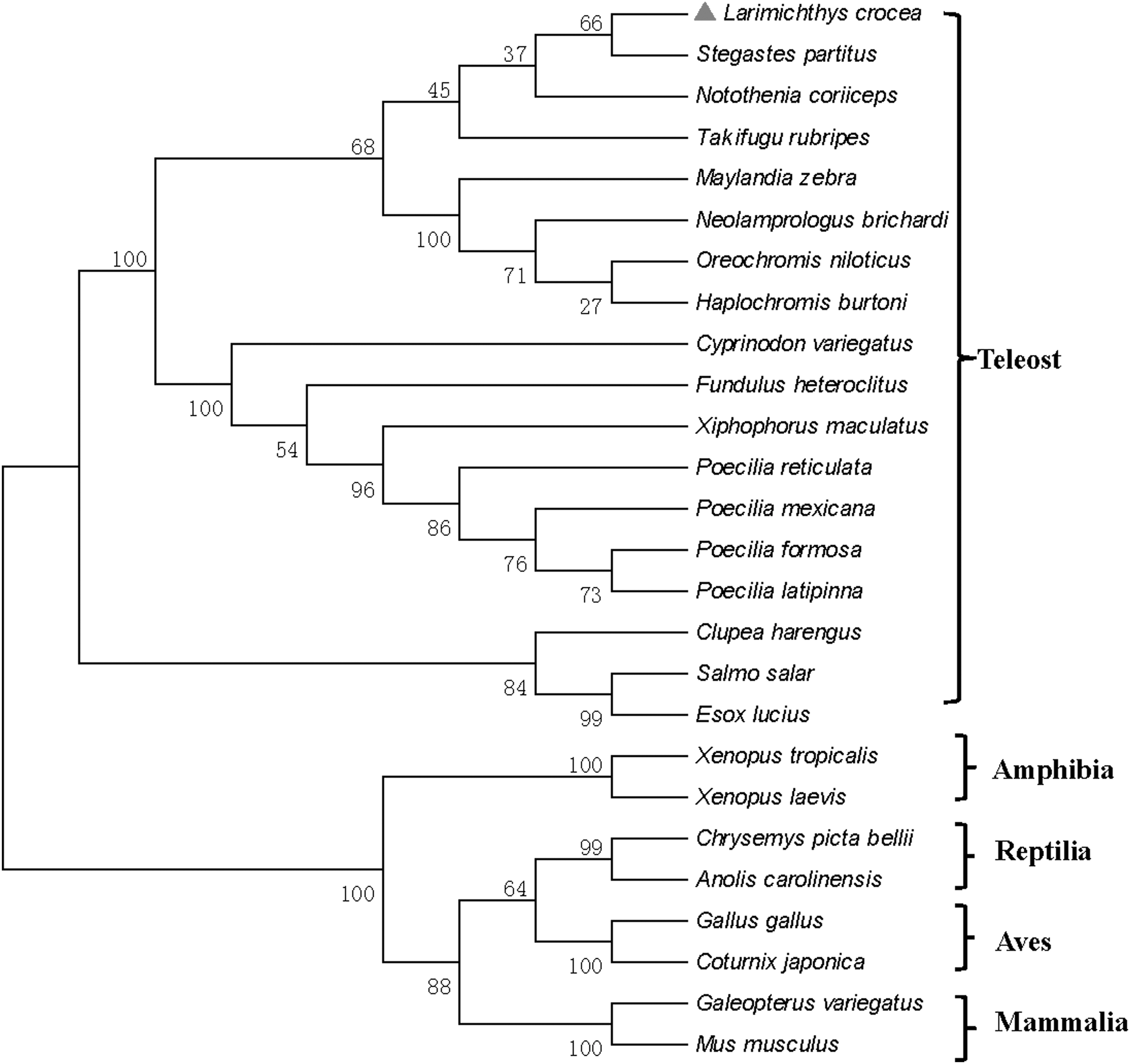
Phylogenetic tree of Klf4 protein from large yellow croaker and other known vertebrates. Numbers indicate the bootstrap confidence values of 1000 replicates.
Embryonic expression patterns of Lc-Klf4
qRT-PCR analysis of embryo Lc-Klf4
The result of qRT-PCR showed that Lc-Klf4 was expressed in all examined embryonic stages with different levels except in multiple-cell stage and mid-blastula stage (Fig. 5). The expression of Lc-Klf4 presented a trend of first decline then rise. The highest expression was detected in closure of blastopore stage (p < 0.01), closely followed by in appearance of optic vesicle stage (p < 0.01), then in formation of eye lens stage (p < 0.05) and formation of yolk plug stage (p < 0.01). In addition, the low and equal expression level of Lc-Klf4 was found in the other embryonic stages (p > 0.05).
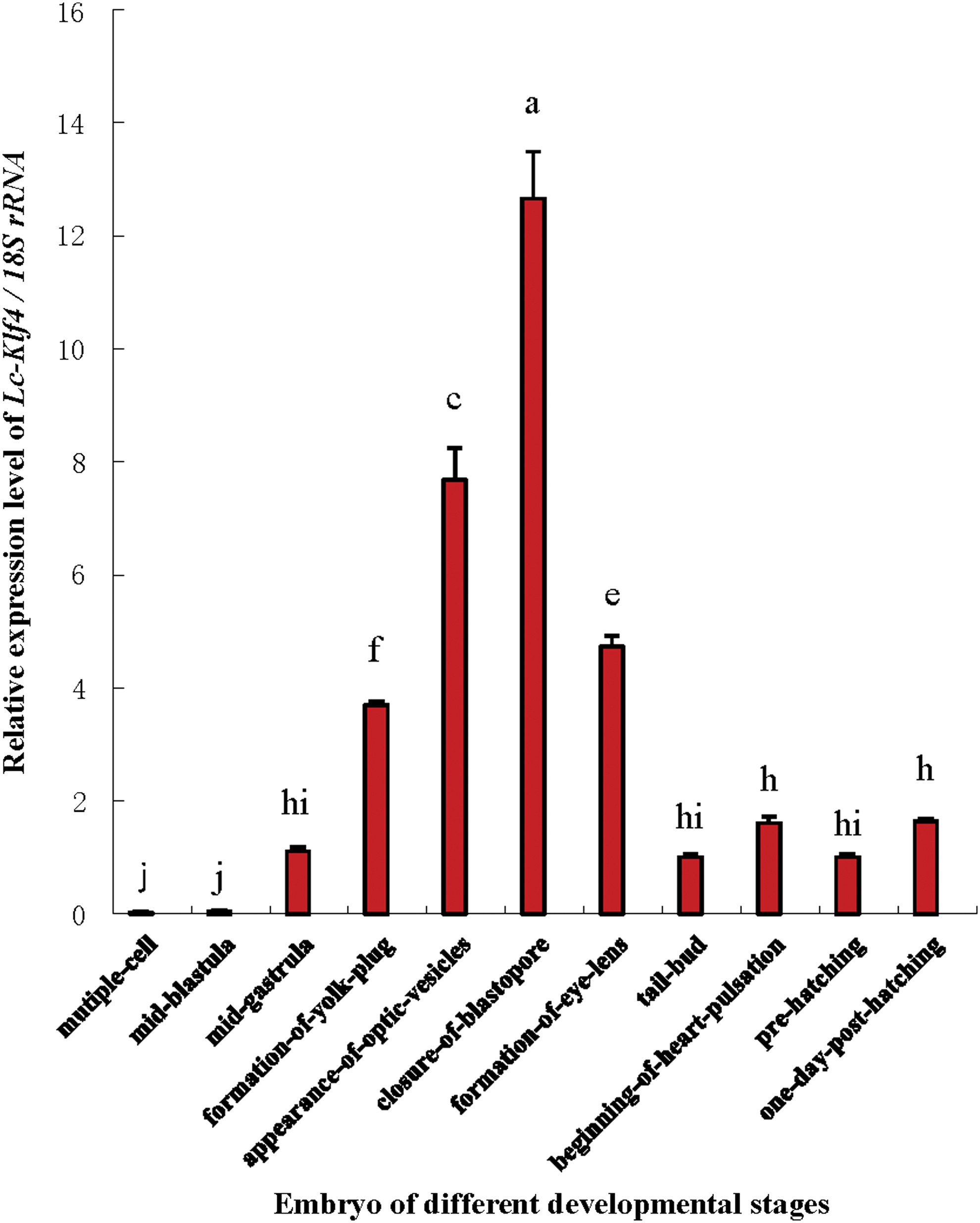
The expression analysis of Lc-Klf4 in different developmental embryonic stages of large yellow croaker. The value of y-axis is assigned by the following: a value of 1 = 1 × RQ value. Each bar represents mean ± SEM (n = 4). The lowercase letters above error bars signify difference as follows: the same letters are not significantly different (p > 0.05), the adjacent letters are significantly different (p < 0.05), and the intervallic letters are extremely significantly different (p < 0.01) in each of the two embryonic development stages. 18S rRNA serves as the reference gene. Color images available online at
Localization of Lc-Klf4 in embryogenesis
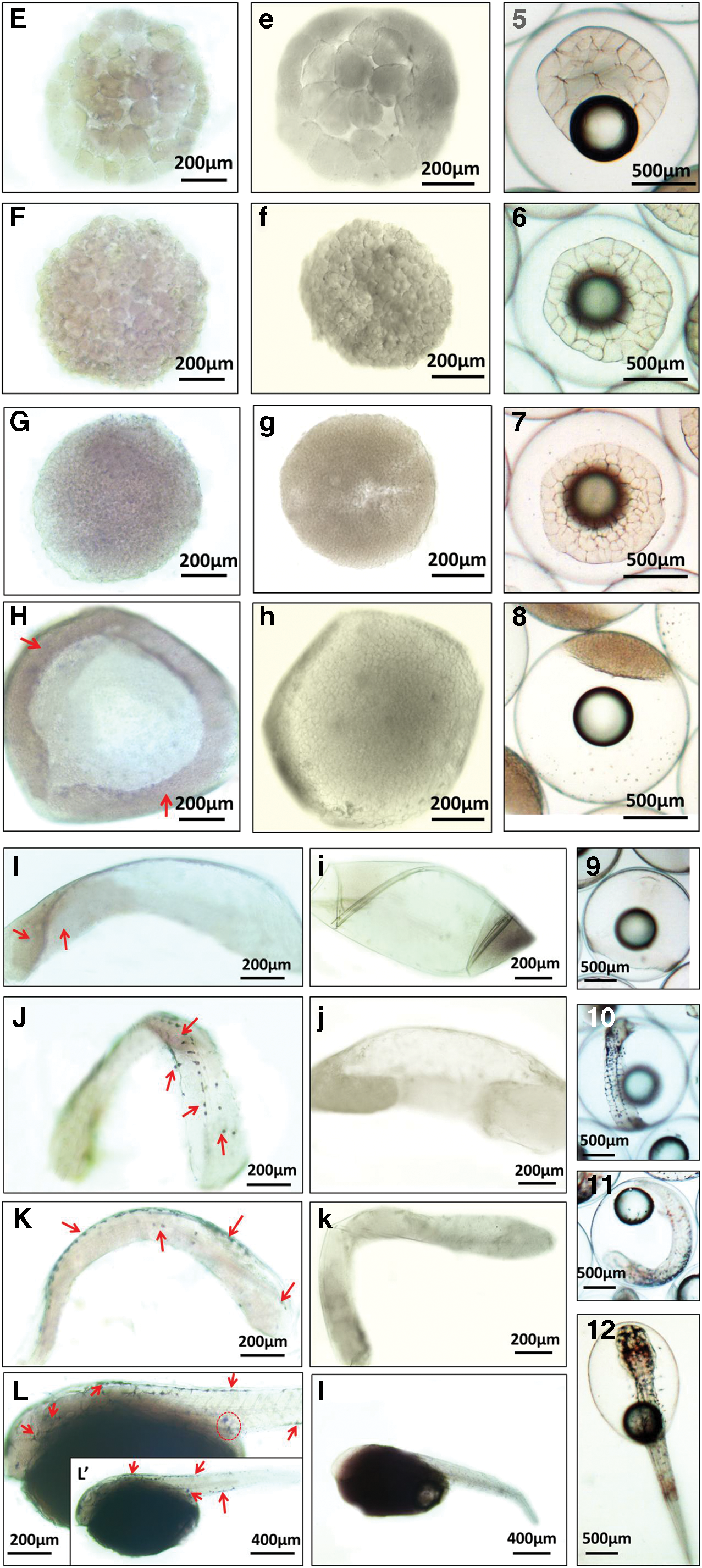
WISH of different developmental embryos demonstrated the distribution of Lc-Klf4 (Fig. 6). Almost no positive signal was detected from 2-cell stage to blastula stage (Fig. 6A–G). In the early gastrula stage, the weak positive signals began to appear in the involute cells, then distributed at the rear of embryos in the late gastrula stage (Fig. 6H, I). The strong positive signals were present in the closure of blastopore stage with punctate distribution in the back of embryos (Fig. 6J) and concentrated gradually in the center of embryonic back and head (Fig. 6K, L, L′). Besides, in the 1-day posthatching stage, positive signals were found in the primordial germ cell (PGC) area and intestine area of embryos (Fig. 6L, L′).

Whole-mount in situ hybridization of Lc-Klf4 in embryogenesis of large yellow croaker.
Adult tissue expression patterns of Lc-Klf4
qRT-PCR analysis of Lc-Klf4 in different adult tissues of both sexes
qRT-PCR analysis of Lc-Klf4 expression in 11 different tissues of female and male large yellow croaker is exhibited in Figure 7A. No matter whether male or female, Lc-Klf4 was expressed widely in examined tissues with different degrees. In female, the highest expression was detected in the brain (p < 0.01), next in gill (p < 0.01), then in eye (p < 0.05), and heart (p < 0.01) in turn. In the other tissues, Lc-Klf4 was expressed with low level. In contrast with female, the highest expression of Lc-Klf4 in male was examined in the heart (p < 0.01), subsequently in testis (p < 0.05), then followed by gill (p < 0.05), and brain (p < 0.01). The expression of Lc-Klf4 in the other tissues was equally low (p > 0.05).
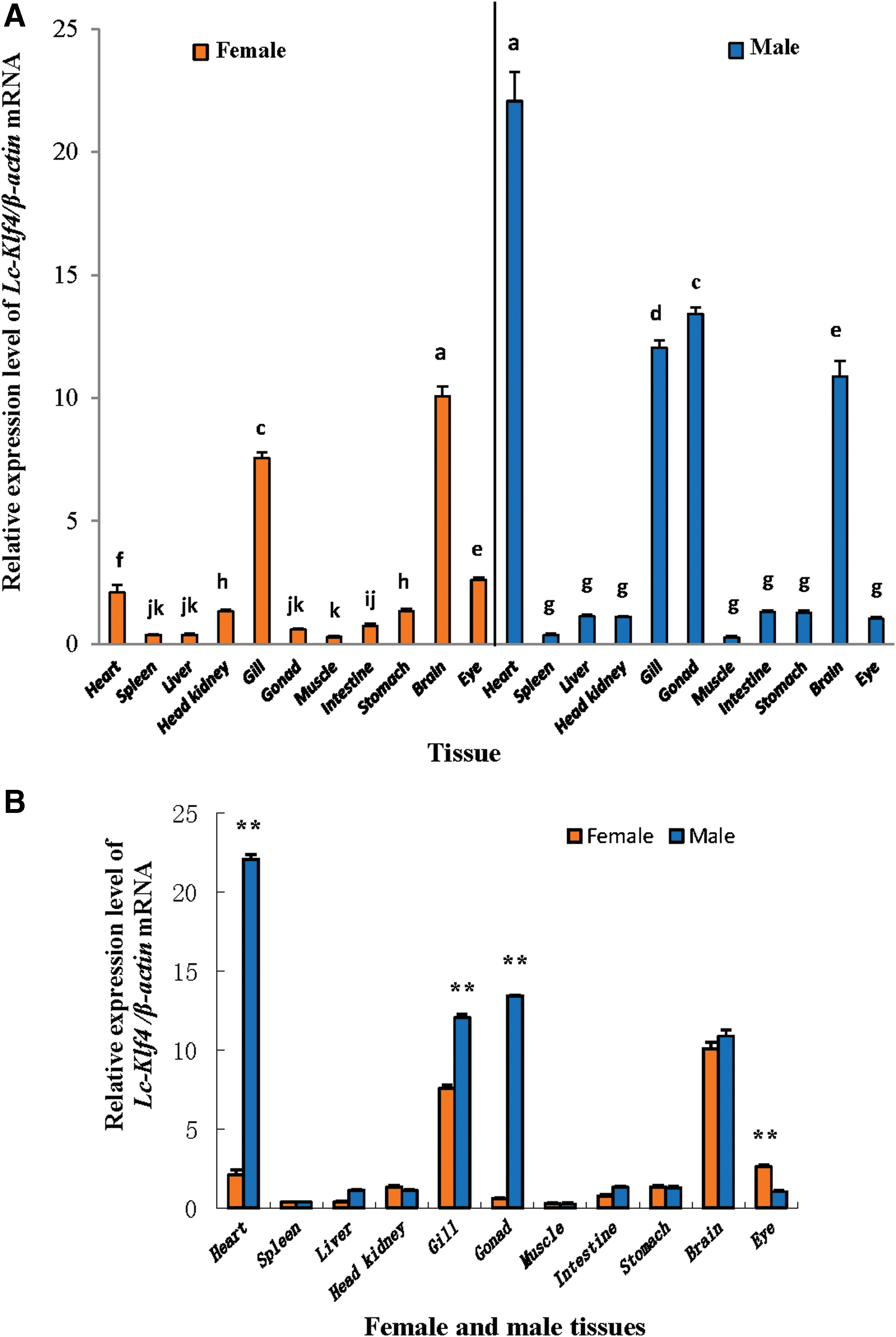
The expression of Lc-Klf4 in different tissues of female and male large yellow croaker
Comparison of Lc-Klf4 expression level in tissues between female and male was performed in Figure 7B. The extremely significant difference appeared in the brain (about 10.6-fold), gonad (about 23.0-fold), and gill (about 1.6-fold), with the level in male higher than in female (p < 0.01), and in eye (about 2.6-fold) with the level in female higher than in male (p < 0.01). Interestingly, Lc-Klf4 was expressed highly and equally in both female and male brains (p > 0.05). Gender difference was absent in the other tissues with low expression level (p > 0.05).
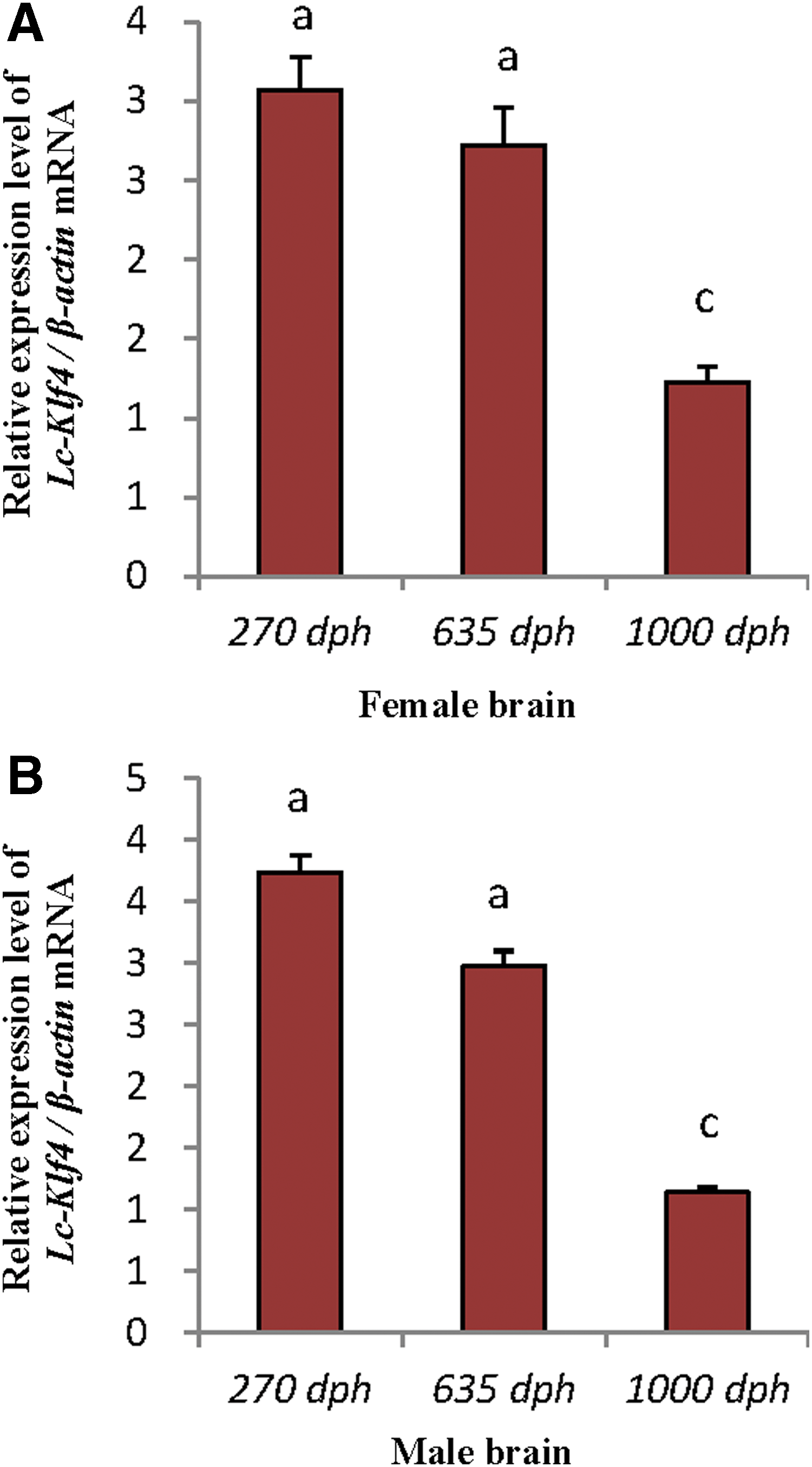
qRT-PCR analysis of Lc-Klf4 in different developmental brain of both sexes
High level of Lc-Klf4 was detected in brains of both sexes. No matter whether male or female brain, the expression of Lc-Klf4 exhibited the same pattern: the level in 270-dph brain and 635-dph brain was equal (p > 0.05) and higher than 1000-dph brain (p < 0.01) (Fig. 8A, B).

The expression analysis of Lc-Klf4 in different developmental testes of large yellow croaker. Each bar represents mean ± SEM (n = 4). The lowercase letters above error bars signify difference as follows: the same letters are not significantly different (p > 0.05) and the intervallic letters are extremely significantly different (p < 0.01) in each of the two developmental testes. β-actin serves as the reference gene.
qRT-PCR analysis of Lc-Klf4 in different developmental testis
The expression of Lc-Klf4 changed with the tendency of first rise, then decline, in testes of different development stages (Fig. 9). The highest expression level appeared in 635-dph testis (mainly including spermatocytes) (p < 0.01), followed by 270-dph testis (mainly including spermatogonia), and 1000-dph testis (mainly including sperm) equally (p > 0.05).
The expression analysis of Lc-Klf4 in different developmental brains of female
Localization of Lc-Klf4 in spermatogenesis
Detected by in situ hybridization on 5-μm-thick paraffin sections, the pattern of Lc-Klf4 in spermatogenesis was obtained. Lc-Klf4 was widely and highly distributed in different developmental spermatids, with positive signals appearing both in cytoplasm and nucleus of all spermatids (Fig. 10A–C). Among them, the distribution of Lc-Klf4 in immature seminiferous lobules (mainly including spermatogonia and spermatocytes) was higher than in mature seminiferous lobules (mainly including spermatids and sperms) (Fig. 10A–C). With spermatogenesis, the positive signal of Lc-Klf4 was weakened slightly (Fig. 10A, C).
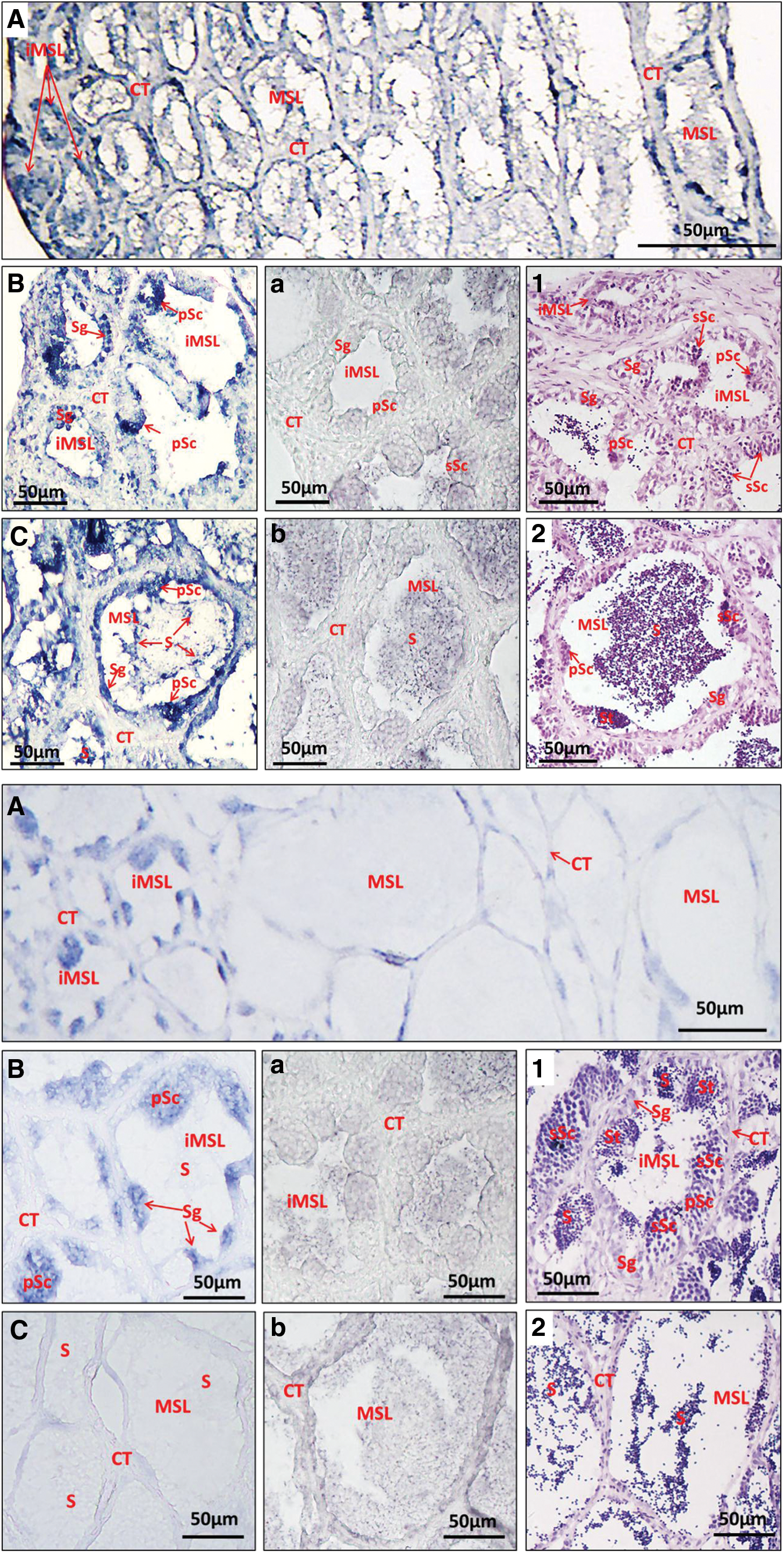
Localization of Lc-Klf4 in large yellow croaker during spermatogenesis by in situ hybridization.
Discussion
As is well known, Klf4 is a classical pluripotency factor and belongs to the Klf family. In this article, we analyzed Lc-Klf4 by bioinformatic method for illustrating its structure and evolution. Lc-Klf4 DNA comprises five exons and four introns as rabbit Klf4 (Tancos et al., 2015). Like Klfs (Turner and Crossley, 1999), Lc-Klf4 contained three highly conserved classical C2H2 zinc fingers at C-terminal, serving as DNA binding domain. C2H2 of Lc-Klf4 contains three α-helices, which bind to the CACCC box of DNA by responsive motif GGGTG (Fig. 1). The N-terminal of Klf4 functions as transcriptional activation and/or repression (Rowland and Peeper, 2006).
Two subtypes of Klf4 were isolated from zebrafish (named Klf4a and Klf4b) (Kawahara and Dawid, 2000; Li et al., 2011). Generally, duplicated genes share different fates and functions compared with the single-copy gene, for example, zebrafish Klf4a is syntenic to mammalian Klf4 with maternally expressed, but no early phenotype and later, functions by repressing proliferation and promoting differentiation of intestinal cell (Li et al., 2011) as mammalian Klf4 (Shields et al., 1996; Katz et al., 2005). However, Klf4b is syntenic to Xenopus Klf17/Neptune and activates zygotically (Kawahara and Dawid, 2000; Onichtchouk et al., 2010) by involving in hatching gland formation and primitive erythropoiesis (Gardiner et al., 2005, 2007). So far, no subtypes of Klf4 are found in studied vertebrates except zebrafish. Similarly, only one single subtype of Klf4 was isolated from large yellow croaker (named Lc-Klf4). Study is needed to determine whether the subtype differentiation of zebrafish Klf4 is an independent divergence or existence in other vertebrates.
The phylogenetic analysis displayed that Lc-Klf4 shared high identity with teleosts (68–94%), but low with tetrapods (<55%) (Table 2, Fig. 3), which meant high conservative function of Klf4 in teleosts and evolutionary differences between teleosts and tetrapods. The low identity of Lc-Klf4 with tetrapods may be a result of specific WGD event in fish lineage (Amores et al., 1998). Moreover, in teleosts, Lc-Klf4 showed the highest identity with S. partitus (94%) (Table 2) supported by the clustering result first with S. partitus (Fig. 4). Besides, T. rubripes (Tetraodontiformes) was clustered in Perciformes subgroup and shared a closer relationship with Lc-Klf4 than some Cyprinodontiformes (Fig. 4), which was inconsistent with the traditional classification system. The high identity of Klf4 among these teleosts indicated that they share a similarly conserved function (Tancos et al., 2015).
The expression level of the gene is closely related to its biological function, so we studied the expression of Lc-Klf4 in adult tissues and embryos in this article for clarifying its function preliminarily. qRT-PCR results of tissues showed that Lc-Klf4 was expressed widely, including in the brain, gill, eye, and heart of females and in the heart, testis, gill, and brain of males (Fig. 7A). The similar wide expression of Klf4 was exhibited in medaka with high expression level in gonad, brain, skin, heart, and gut, especially highest in testis (Wang et al., 2011). Inconsistently, no Lc-Klf4 was detected in ovary as well as in mouse (Behr and Kaestner, 2002; Tancos et al., 2015). There were differences and similarities of Lc-Klf4 tissue expression pattern compared with mammals. For example, Lc-Klf4 was absent in adult stomach and intestines, which was high in mammals (Bieker, 2001; Flandez et al., 2008; Tancos et al., 2015), but highly present in gills with no expression in mammals. Notably, the expression of Lc-Klf4 was examined in the intestine area of embryos (Fig. 6L, L′), which indicated the possibility of involving in intestinal development. The similar expression of Lc-Klf4 existed in the brain and heart of rabbit (Tancos et al., 2015) and in testis and eye of mouse (Shields et al., 1996; Behr and Kaestner, 2002; Swamynathan et al., 2007). These similarities and differences of Klf4 expression may be due to gene conservation and species differences. Moreover, the expression of Lc-Klf4 presented gender differences in the heart, gill, and gonad with male>female and in eye with female>male (Fig. 7B), which suggested that Lc-Klf4 might function differently in both sexes. Interestingly, the expression of Lc-Klf4 seemed to be more inclined to males. Our results provide a beneficial reference to the mechanism research of gender differences.
Furthermore, the expression of Lc-Klf4 in different developmental testes was examined by qRT-PCR with the highest expression level in 635-dph testis (Fig. 8). The results from in situ hybridization also confirmed that Lc-Klf4 was widely and highly distributed in all spermatids with a moderate level in spermatogonia, peak in spermatocytes, and decline in spermatids and sperm (Fig. 10). Clearly, the expression of Lc-Klf4 in testis suggested a great possibility for involving in spermatogenesis progression. The similar expression pattern of Klf4 was reported in medaka too (Wang et al., 2011). Likewise, Klf4 functioned as a key factor for postnatal development and differentiation of the mouse testis by strong expression in postmeiotic germ cells undergoing spermiogenesis as well in somatic Sertoli cells (Behr and Kaestner, 2002). Besides, the presence of Lc-Klf4 in PGC area of embryos (Fig. 6L, L′) also supported the view that Lc-Klf4 was closely related with spermatogenesis.
To better clarify the role of Lc-Klf4, the expression in different developmental embryonic stages was studied too. In situ hybridization results showed that Lc-Klf4 was undetected in early embryos (ranging from 2-cell stage to blastula stage) until gastrula stage with high expression in closure of blastopore stage, beginning of heart pulsation stage, and 1-day hatching stage (Fig. 6), which revealed a zygotic expression pattern in embryogenesis. The qRT-PCR analysis of Lc-Klf4 was coincident by lack of expression in early embryos and strong expression in order of closure of blastopore stage, appearance of optic vesicle stage, and formation of eye lens stage (Fig. 5). The similar zygotic expression pattern was observed in mouse Klf4, which lacked expression in early embryos until the blastocyst stage (Ema et al., 2008). Conversely, the expression was mainly maternal in fish and Xenopus (Wang et al., 2011; Cao et al., 2012). For instance, medaka Klf4 was expressed in early embryonic stages ranging from cleavages to the late blastula stage (Wang et al., 2011), as well as rabbit Klf4 (Tancos et al., 2015). A similar difference was also reported in zebrafish with Klf4 expression in early embryos (Luo et al., 2011). Of course, weak zygotic expression was also observed during early embryogenesis in fish and Xenopus (Wang et al., 2011; Cao et al., 2012), so Klf4 was expressed both maternally and zygotically during Xenopus embryogenesis with difference in abundance in different regions of gastrula (Cao et al., 2012). Notably, the embryonic expression pattern of Lc-Klf4 was similar to mouse instead of fish. The question of whether there is an independent divergence on Lc-Klf4 needs further confirmation.
As reported, Klf4 participated in organogenesis during embryonic development. For example, Klf4 functioned by negative regulation of neurite outgrowth in hippocampal and cortical neurons of the central nervous system (CNS) (Moore et al., 2009); knockout of Klf4 resulted in increase of neurite growth of retinal ganglion cells (RGCs) and axon regeneration (Moore et al., 2009; Apara and Goldberg, 2014). In our article, the high expression of Lc-Klf4 in brain was observed too (Fig. 7) with high level in 270-dph and 635-dph brains in both male and female (Fig. 9). These results were coincident with strong expression of Lc-Klf4 in CNS of 1-day hatching stage embryo revealed by WISH (Fig. 6L, L′). Taken together, Lc-Klf4 may be closely related to neurogenesis and neural survival, the mechanism was considered to be further researched. Furthermore, Klf4 also played a vital role in eye development such as in humans (Chiambaretta et al., 2004) and mice (Swamynathan et al., 2007). In detail, Klf4 regulated mouse eye development by developmental regulation of RGCs with limiting retinal ganglion cell axon bundle thickness (Fang et al., 2016). In this article, moderate expression of Lc-Klf4 was detected in the female eye (Fig. 7); high expression level of Lc-Klf4 from appearance of optic vesicle stage to formation of eye lens stage (Fig. 5, 6J, K) confirmed this expression pattern, which seemed to suggest a role of Lc-Klf4 in eye development, especially in the female eye.
In a word, we first analyzed Lc-Klf4 by bioinformatic method in this article. Lc-Klf4 conformed to characteristics of Klfs and conserved evolutionally in teleosts. We also examined temporal–spatial expression patterns of Lc-Klf4 in different tissues and embryos. Lc-Klf4 was expressed high in adult tissues of the heart, gill, gonad, brain, and eye with gender differences. Lc-Klf4 began to be expressed zygotically from gastrula stage with high level in appearance of optic vesicle stage to formation of eye lens stage. The data exhibited vital roles of Lc-Klf4 in spermatogenesis, embryogenesis, and relationship of physiological processes.
Footnotes
Acknowledgments
The work was supported by the Natural Science Foundation of China (No. 41306174, 31272653), the Program of the Science and Technology Department of Fujian Province (No. 2013N0024), the start up fund of Jimei University (No. ZQ2017003), and the Innovation Team Foundation of Jimei University (No. 2010A001).
Disclosure Statement
No competing financial interests exist.