
Abstract
Gremlin1, the antagonist of bone morphogenetic protein-7 and one of the target genes of transforming growth factor (TGF)-β signal pathway, plays an important role in embryonic development and its expression decreases along with aging. To explore the expression of gremlin1 in liver fibrosis and the causal link between gremlin1 and hepatic stellate cell (HSC) activation, we detected the expression of gremlin1 in mice with hepatic fibrosis induced by porcine serum using real time quantitative PCR (RT-qPCR) and immunohistochemical staining. The hepatic fibrosis mice were evaluated by the external feature of the liver, histology, hepatic function, collagen deposition, and the expression of fibrosis-related genes (genes COLIα2 and COLIVα2) in the liver. In the HSC-T6, western blotting was used to analyze the expression of α-smooth muscle actin (α-SMA), COL1α, and TGF-β1 in conditions of overexpression of gremlin1 or gremlin1 being knocked down by specific siRNA, respectively. The results showed that the mRNA expression of the gremlin1 gene was significantly increased consistent with increased expression of COLIα2 and COLIVα2 in the liver tissue of the hepatic fibrosis mice. Increased expression of gremlin1 coincided with the same area of the collagen deposition. Furthermore, the results also showed that the expression of α-SMA, COLIα1, and TGF-β1 was consistent with the expression of gremlin1 not only in the HSC-T6 overexpressing gremlin1 but also in the HSC-T6 that gremlin1 is knocked down by specific siRNA. The findings suggest that gremlin1 might play an important role in the progression of hepatic fibrosis and that it modulates HSC activation.
Introduction
G
Gremlin, which is important in embryonic development (Merino et al., 1999; Michos et al., 2004), was first isolated from the neural crest of the Xenopus as an antagonist of BMP signaling pathway (Sudo et al., 2004). Although the expression of gremlin usually decreases in adulthood (Wade et al., 2005), it can also increase obviously in the fibrosis diseases (Koli et al., 2006; Carvajal et al., 2008). TGF-β stimulation upregulated gremlin expression in human kidney epithelial cells, and the expression of gremlin was also elevated in diabetic nephropathy. Transcriptional profiling in a mouse hepatic fibrosis model also suggested that gremlin might be a novel marker of liver fibrogenesis (Boers et al., 2006). Gremlin has three alternative splicing patterns as gremlin1, gremlin2, and gremlin3 (Wordinger et al., 2008), and gremlin1 is the most common isoform. Recent studies have reported higher gremlin1 expression in cirrhosis than that in other liver diseases (Guimei et al., 2012) and a significant increase in gremlin1 levels in rat or mouse hepatic fibrosis models (Boers et al., 2006; Zhao et al., 2014). Nevertheless, it is unknown whether the quantitative relationship between collagen and gremlin1 in hepatic fibrosis exists or not.
Previous studies have revealed that hepatic stellate cells (HSCs) play a pivotal role in hepatic fibrogenesis (Friedman, 2008). In chronic liver injury, the fibrogenic cytokines, including tumor necrosis factor α, TGF-β, interleukin 1, and platelet-derived growth factor, promote HSCs to transdifferentiate, changing from quiescent phenotypes to activated myofibroblastic cells. Activated HSCs express fibrogenic proteins and α-smooth muscle actin (α-SMA), lose their lipid and retinoid storages, and then upregulate extracellular matrix (ECM) synthesis, leading to a large amount of ECM deposited in the liver (Friedman, 2003; Tsukada et al., 2006). TGF-β is the key player in promoting the activation and transdifferentiation of HSCs (Bauer and Schuppan, 2001; Cheng et al., 2009). Meanwhile BMPs, members of the TGF-β superfamily, named for inducing bone and cartilage formation (Marsell and Einhorn, 2009; Bragdon et al., 2011), hampered the progress by antagonizing biological activities of TGF-β (Gressner et al., 2007). TGF-β and BMPs, respectively, bind to serine/threonine kinase receptors type I and type II, subsequently leading to phosphorylation of Smad2/3 (TGF-β signaling pathway) or Smad1/5/8 (BMP signaling pathway), which combine with Smad4 and then shuttle into the nucleus to regulate the expression of their downstream genes (Wang and Hirschberg, 2004; Breitkopf et al., 2006; Walsh et al., 2010). TGF-β and BMP-7 are antagonistic relationships to each other in hepatic fibrogenesis (Kinoshita et al., 2007; Tacke et al., 2007) and gremlin1 plays a role as a bridge between TGF-β and BMP-7 signaling. Nevertheless, it is still unknown what the causal relationship of influence of gremlin1 on the activation of HSCs might be like.
In this study, we hypothesized that gremlin1 might modulate HSC activation. To test this possibility, first, we investigated the quantitative relationship between collagen and gremlin1 in a mouse hepatic fibrosis model. Subsequently, we explored the influence of upregulation and downregulation of gremlin1 expression on HSC activation in vitro.
Materials and Methods
Hepatic fibrosis animal models
The approval for animal experiments was obtained from the Animal Ethics Committee of the Animal Laboratory Center of China Three Gorges University (CTGU), before the research. All the animal experiments were performed in specific pathogen-free facilities at the Animal Laboratory Center of CTGU. The animals received humane care in compliance with the criteria outlined in the “Guide for the Care and Use of Laboratory Animals.” The mice were purchased from the Disease Control Center of Hubei Province (China) and acclimatized to the appropriate conditions for a week before in vivo studies.
Twenty-four healthy 6-week-old female Balb/c mice (18 ± 22 g) were randomly divided into experimental (n = 14) and normal (n = 10) groups, and i.p. injected with 0.1 mL/10 g total porcine serum and 0.1 mL/10 g saline, respectively, once a week for 6 weeks. After weighing and gross inspection, all the mice were anesthetized with urethane and sacrificed, followed by collection of blood, liver, and spleen samples. The blood was used for the biochemical assay. Some portions of liver tissues were fixed in formalin for histological and immunohistochemical (IHC) analyses while the others were rapidly frozen in liquid nitrogen and stored at −80°C for quantitative PCR (qPCR) analysis.
Biochemical assay
The enzyme activities of alanine aminotransferase (ALT), aspartate aminotransferase (AST), total protein (TP), albumin (AP), globin (GP) were measured by Olympus 560 analyzer (Olympus, Tokyo, Japan) in the second clinic medical college of CTGU, China.
Histopathology and immunohistochemistry
Liver tissues of mice were harvested, fixed in 10% neutral buffered formalin for 24 h, processed, and embedded into paraffin blocks. Routine hematoxylin and eosin (H&E) staining was performed on 4 μm sections of tissue cut from formalin-fixed, paraffin-embedded (FFPE) blocks. Morphology of liver and proliferation of total collagen were analyzed using H&E and Sirius red stain, respectively. The degree of hepatic fibrosis in mice was assessed using the Scheuer's scoring system as follows: grade 0, no fibrosis; grade 1, portal fibrosis; grade 2, periportal fibrosis; and grade 3, septal fibrosis (Scheuer, 1991).
For immunohistochemistry, FFPE tissue blocks were sectioned at 4 μm, baked for 1.5 h for deparaffinization at 65°C, and rehydrated before antigen retrieval using a standard xylene/alcohol protocol. The endogenous peroxidase activity was blocked with 3% hydrogen peroxide in TBST for 5 min. An antigen retrieval step was conducted in citrate buffer (10 mM, pH 6) for 10 min under low microwave power. The blocker was drained and gremlin1 primary antibody (C-19; Santa Cruz) was applied at a concentration of 1 μg/mL. The control sections were incubated with the appropriate IgG negative control. The slides were incubated at 37°C for 1 h, followed by overnight incubation at 4°C (primary antibody at 1:50 dilution) and 1 h at 37°C with rabbit F(ab′)2 polyclonal secondary antibody to goat IgG—H&L (HRP) (Abcam, Cambridge, United Kingdom). The final detection step was carried out using 3, 3′-diaminobenzidine (DAB; Dako, Carpinteria) as the chromogen. The sections were counterstained with Mayer's hematoxylin for 3 min and coverslipped.
RNA isolation, reverse transcription, and qPCR
Liver tissue (100 mg) and HSC-T6 cells were homogenized on ice with 1 mL and 800 μL TRIzol reagent (Invitrogen, Thermo Scientific), respectively. Total RNAs were extracted and reverse transcribed to cDNA according to the manufacturer's instructions (Fermentas, Thermo Scientific), followed by quantification by measuring the ratio of absorbance at 260 and 280 nm (A260/A280 = 1.8–2.0) with the Multiskan spectrum instrument (Thermo Scientific). All primers for real-time qPCR assays were synthesized by Sangon Biotech (Shanghai, China). The sequences are listed in Table 1.
qPCR analysis was performed with the fluorescence qPCR system (CFD-3220; MJ Research, Bio-Rad) using the Fast SYBR Green Master Mix (Thermo Scientific). The cycling protocol was as follows: initial denaturation at 94°C for 5 min, followed by 30 cycles of 94°C for 30 s, 58°C for 30 s, and 72°C for 1 min. A melting curve was generated to ensure no contamination. qPCR of gremlin1, collagen Iα2, and collagen IVα2 genes and the normalizing gene, β-actin, was performed in triplicate for each sample and included a no-template control to rule out contamination and primer-dimer formation. The data collected automatically can be used to analyze the threshold cycle (Ct) value. Expression changes of gremlin1, collagen Iα2, and collagen IVα2 mRNAs were detected using the 2−ΔΔCt method (Livak and Schmittgen, 2001).
Preparations for recombinant plasmids
The pcDNA3.1-gremlin1 was constructed by inserting coding sequence (CDS) of Rattus norvegicus gremlin1 (NM_019282.3) into the multiple cloning site (MCS) of pcDNA 3.1 (Invitrogen, Thermo Scientific) between restriction enzyme sites KpnI and XhoI.
The pLKO.1-si-gremlin1 1#, pLKO.1-si-gremlin1 2#, pLKO.1-si-gremlin1 3#, pLKO.1-si-gremlin1 4#, and pLKO.1-si-NC were constructed by inserting the cDNA encoding candidates of gremlin1-specific shRNA and scramble shRNA into a cloning site between AgeI and EcoRI in PLKO.1 vector, respectively. The cDNAs encoding shRNAs (1#, 2#, 3#, 4#, and the scrambled shRNA) are annealed of two complementary DNA strands designed in Ambion online service (listed in Table 2) (Ambion, Thermo Scientific).
All the DNA segments, including the primers for cDNA cloning and RT-qPCR used in this study, were synthesized by Sangon Biotech (Shanghai, China), and all the recombinant plasmids were further confirmed by DNA sequencing (Sangon Biotech, Shanghai, China).
Cell culture, plasmid transfection of HSC-T6 cells
The HSC-T6 cell line was maintained in our laboratory. HSC-T6 cells were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% newborn calf serum (Invitrogen, Thermo Scientific). Cells were seeded in a six-well plate for western blot at a density of 4 × 105 cells per well in DMEM in a humidified atmosphere containing 5% CO2 for 24 h at 37°C.
HSC-T6 cells were transiently transfected with pcDNA3.1-gremlin1 or the gremlin1-specific short hairpin RNA (shRNA) expression plasmid based on pLKO.1, respectively, using TurboFect transfection reagent (Thermo Scientific), according to the manufacturer's instructions. The cells were collected 48 h after transfection to perform western blot analysis.
Western blot analysis
The cells were harvested and lysed in lysis buffer, 25 mM Tris–HCl pH 7.5, 2.5 mM ethylene diamine tetraacetic acid (EDTA), 137 mM NaCl, 2.7 mM KCl, 1% sodium deoxycholic acid, 0.1% SDS, 1% Triton X-100, and 2 mM phenylmethanesulfonyl fluoride (PMSF), and a protease inhibitor cocktail for 30 min at 4°C (Sigma-Aldrich Corp.). The supernatant fractions were transferred to a fresh tube after centrifugation at 20,238 g for 30 min at 4°C using a centrifuge 5424 R (Eppendorf, Hamburg, Germany). Protein concentrations were measured using a BCA Protein Assay kit (Thermo Scientific). An equal amount of protein from each sample was separated via sodium dodecyl sulfate/polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene difluoride membrane incubated with different primary antibodies for 12 h at 4°C. After washing with TBST buffer (20 nM Tris–HCl pH 7.5, 150 mM NaCl, 0.05% Tween-20), the sections were incubated with a secondary antibody for 45 min at room temperature. The membranes were subsequently washed with TBST. The chemiluminescence on the membrane was detected using the ChemiQ 4800mini imaging system (Ouxiang, Shanghai, China). Densitometric analyses of band intensities were performed using ImageJ software (version 1.38 × ; National Institutes of Health). Primary and secondary antibodies as well as dilution ratios for western blot assays are listed in Table 3.
α-SMA, α-smooth muscle actin; TGF, transforming growth factor.
Statistical analysis
The data are expressed as mean ± SD. Statistical analysis was performed by SPSS 10.0 statistical analysis software. A value of p < 0.05 was considered significant.
Result
The expression of gremlin1 was increased in hepatic fibrosis model mouse liver
To explore the expression of gremlin1 in liver tissue with fibrosis, a mouse model of hepatic fibrosis was induced by porcine serum. External features of the mice liver appeared as multinodular features and a hard consistency after the injection of 0.1 mL/10 g of porcine serum, once per week for 6 weeks, suggesting an obvious hepatic fibrosis change (Fig. 1A). The hepatosomatic index (HSI) and spleen index (SI) of the hepatic fibrosis model mice were higher than that of the saline injection mice. Besides ALT, AST and GP were higher but AP was lower in hepatic fibrosis model mice than that in the saline injected mice (Table 4).
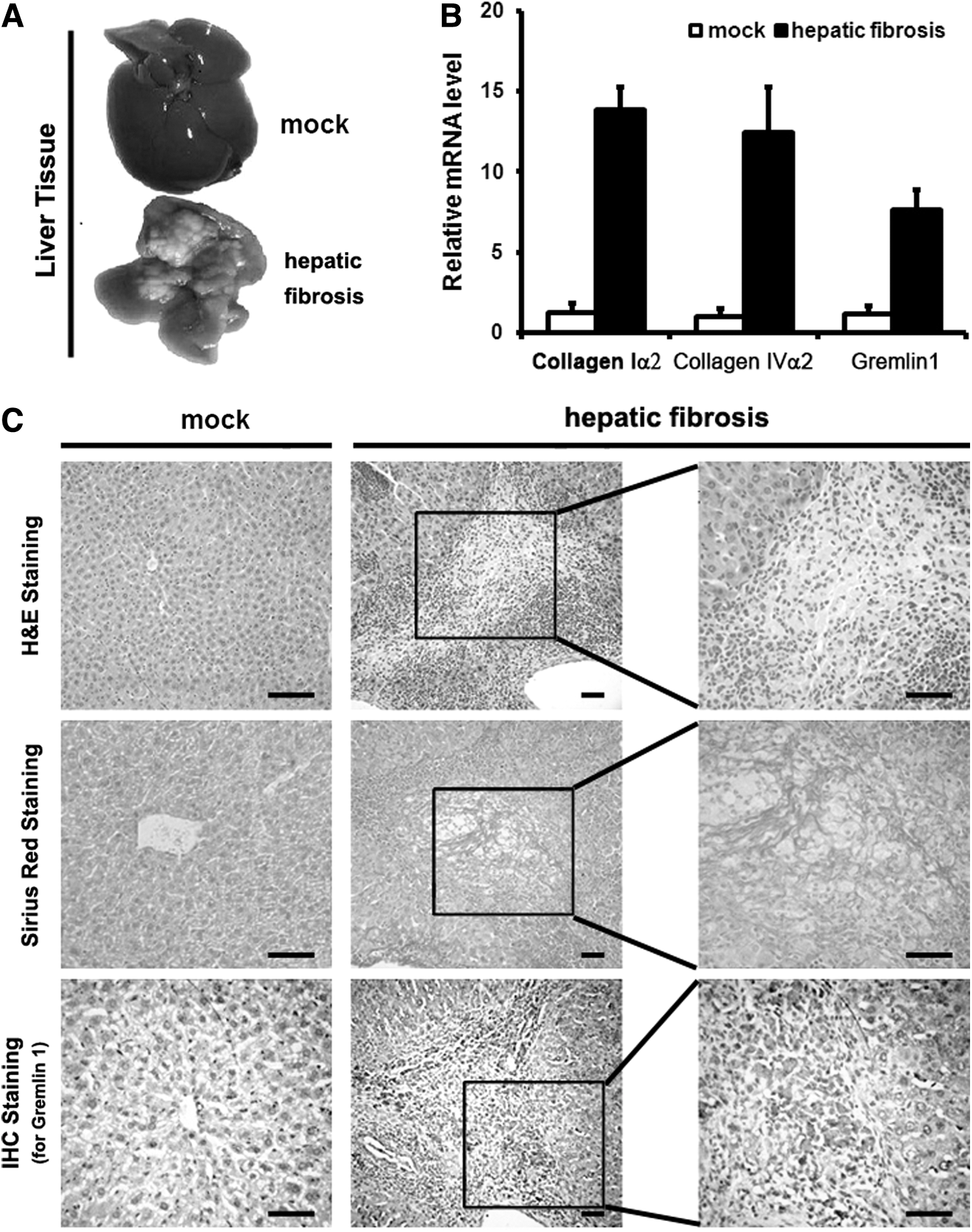
Gremlin1 is upregulated in hepatic fibrosis tissue of model mice.
p < 0.01 hepatic fibrosis model mice versus mock group mice.
p < 0.05 hepatic fibrosis model mice versus mock group mice.
ALT, alanine aminotransferase; AP, albumin; AST, aspartate aminotransferase; GP, globin; HSI, hepatosomatic index; SI, spleen index; TP, total protein.
The hepatic fibrosis model mice showed bridging fibrosis connecting portal areas, and central veins in a portal to portal and portal to central pattern were accompanied by a large amount of infiltrating inflammatory cells and portal fibrosis, observed after being stained by Sirius red, whereas only mild inflammatory cell infiltration around the portal area was found in negative control mice, with no collagen deposition (Fig. 1C and Table 5). By IHC analysis, we identified gremlin1-positive area consistent with collagen deposition. On the contrary, gremlin1-positive area was rarely observed in the negative control group (Fig. 1C). We also analyzed the expression of gremlin1, COLIα2, and COLIVα2 by qPCR. The results showed that the concentration of the gremlin1 mRNA (7.62 ± 1.24), the COLIα2 mRNA (13.81 ± 1.48), and the COLIVα2 mRNA (12.44 ± 2.81) increased in hepatic fibrosis model mice compared with the mock group mice (1.15 ± 0.49, 1.28 ± 0.54, 1.03 ± 0.45, p < 0.01), respectively (Fig. 1B), suggesting that the expression of gremlin1 in hepatic model mice induced by porcine serum is elevated and in consonance with the expression of COLIα2 and COLIVα2.
p < 0.01 porcine serum-treated group versus mock control group.
The overexpression of gremlin1 enhances the activation of HSCs
To elucidate the effect of gremlin1 on the HSCs, eukaryotic expression vector pcDNA3.1-gremlin1 was transfected into HSC-T6 cells. After 48 h of transfection, all the cells were collected and the TPs were analyzed by western blotting assay. First, we detected the expression of α-SMA, a marker of activated HSCs, which was upregulated visibly in HSC-T6 cells. Subsequently, the expression of COLIα1, one of the ECMs being secreted by activated HSCs, was confirmed to be upregulated. The TGF-β expression was also analyzed, and the result showed that TGF-β expression is obviously upregulated (Fig. 2). These data imply that gremlin1 overexpression in HSCs can promote activation of HSCs, which may due to gremlin1-related up-expression of α-SMA and TGF-β, and can boost the signal transduction mediated by TGF-β, leading to enhanced ECM synthesis.
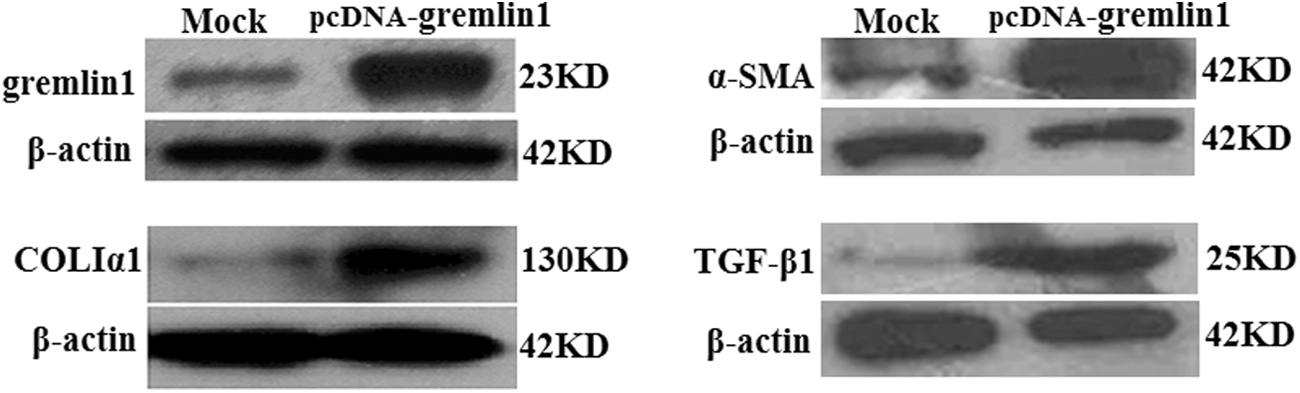
Overexpression of gremlin1 enhances HSC-T6 cell activation. Western blot showed that expression of gremlin1 increased obviously in HSC-T6 cells transfected with pcDNA-gremlin1 plasmid. Western blot showed that expression of collagen Iα1, α-SMA, and TGF-β also increased in HSC-T6 cells transfected with pcDNA-gremlin1 plasmid. α-SMA, α-smooth muscle actin; HSC, hepatic stellate cell; TGF, transforming growth factor.
Downregulation of gremlin1 expression suppresses the activation of HSCs
To test the influence of downregulation of gremlin1 on activation of HSCs, we used lentiviral vectors to express small interference RNA (siRNA) against rat gremlin1. First, the gremlin1-specific shRNA candidates and the scrambled sequence were designed based on rat gremlin1 mRNA from online service (Ambion), then we constructed the recombinant plasmids pLKO.1-gremlin1-sir by synthesizing the cDNAs (Sangon Biotech, Shanghai, China) of the shRNAs and inserting it to the plasmids pLKO.1. HSC-T6 cells were transfected transiently with five recombinant plasmids pLKO.1-gremlin1-sir1/sir2/sir3/sir4 and scr (termed Gsir1, Gsir2, Gsir3, Gsir4, and Gscr), respectively, and the efficiencies of the siRNA to knock down gremlin1 were checked by western blotting assay, the results of which revealed two shRNAs, Gsir1 and Gsir2, able to knock down gremlin1 partially, and we selected Gsir2 to complete the follow-up experiment. As predicted, the expression of α-SMA, COLIα1, and TGF-β significantly decreased after being transfected with Gsir2 (Fig. 3).

Knockdown of gremlin1 expression leads to suppression of HSC activation. Transfection of HSC-T6 cells with gremlin1-specific shRNA-expressing vector, pLKO.1-si-gremlin1 3# or 4#, led to partial downregulation of gremlin1, being observed via western blot. Transfection of HSC-T6 cells with pLKO.1-si-gremlin1 3# significantly suppressed collagen Iα1, α-SMA, and TGF-β expression.
Discussion
Gremlin plays an important role in embryonic formation and in early development. At the mid-blastula transition, gremlin is detected in the entire blastoderm. In the gastrula, gremlin becomes restricted to the dorsolateral region of the embryo, and during somitogenesis, it is strongly expressed in the presomitic mesoderm and developing somites, and in the ventral neural tube (Nicoli et al., 2005). In the embryonic development, gremlin promotes limb formation through regulating BMP (Zuniga et al., 2004) and triggers the growing development of the ureter, kidney, and lung through antagonism to the function of BMPs (Martinez et al., 2002). In adults, the gremlin gene is only expressed in the brain, testes (Topol et al., 1997), and ovary (Topol et al., 2000), but its expression is a little or none in the heart, liver, kidney, skeletal muscle, and fat cells (Wade et al., 2005). However, it is obviously elevated in fibrosis diseases, such as interstitial fibrosis and diabetic nephropathy (Zeisberg and Kalluri, 2008) or lung interstitium and fibroblasts of patients with idiopathic pulmonary fibrosis (Koli et al., 2006). To investigate the gene expression profile during different stages of the process in which quiescent HSCs are activated and transdifferentiated to myofibroblasts, Boers W found that gremlin expression increased in this process (Boers et al., 2006). After hepatic fibrosis models of mice were set up by injecting the pig serum into the mice intraperitoneal cavity and were evaluated by the external feature of the liver, HSI, SI, hepatic function analysis of the serum (ALT, AST, TP, AP, GP), histologic features, collagen deposition, and expression of fibrosis-related genes (genes COLIα2 and COLIVα2) of the liver, we also showed that gremlin1 gene was expressed in the hepatic tissue of the hepatic fibrosis mice by qPCR and IHC and its expression was consistent with COLIα2 and COLIVα2. All the findings listed above suggest that expression of gremlin1 is elevated in hepatic fibrosis. However, the causal link between gremlin1 and HSC activation is unclear.
The balance between TGF-β and BMP signaling plays an important role in tissue homeostasis, and gremlin, one of BMP antagonists, is a downstream profibrotic mediator of TGF-β in cultured renal cells (Rodrigues-Diez et al., 2012). TGF-β may promote the expression of gremlin through Smad2/3/4 complex binding to TGF-β responsive element (TRE), which is an enhancer of TGF-β target genes. At the same time, gremlin may promote TGF-β signaling, for example, tubular gremlin1 overexpression is correlated with the renal upregulation of profibrotic factors, such as TGF-β and α-SMA, resulting in increased susceptibility to renal damage (Droguett et al., 2014). Gremlin1 overexpression induces submesothelial thickening, fibrosis, and angiogenesis in C57BL/6 mouse peritoneum (Siddique et al., 2014) and results in activation of alveolar epithelial cells with proliferation and apoptosis, as well as partly reversible lung fibrosis (Farkas et al., 2011). To further clarify the relationship of gremlin1 with TGF-β, collagen, and HSC activation, the expression of gremlin1 was upregulated and downregulated in the HSC-T6, respectively. We have found that the expression of α-SMA, COLIα1, and TGF-β1 is consistent with the expression of gremlin1. So, gremlin1 promotes HSC activation and it may play an important role in the development of hepatic fibrosis. Gremlin1 is a highly conserved 184 amino acid protein that contains a signaling peptide (1–24) and nuclear localization signal sequences (positions 145, 166, 163, 164). Gremlin1 exists in both secreted and cell-associated forms. Mechanically, we speculate that the secreted gremlin1 may promote the expression of TGF-β1 indirectly through antagonizing the function of BMPs and the cell-associated gremlin1 may enter into the nucleus and upregulate the expression of TGF-β1 directly. However, these mechanisms need further to be confirmed. Because of the relationship of gremlin1 with the TGF-β1, α-SMA, and COLIα1, gremlin1 may become a therapy target gene of hepatic fibrosis.
Footnotes
Acknowledgments
The authors thank Shan-Bing Yin for English language editing and Yan-Ling Liu for the experiment. They are thankful for the financial support of the National Natural Science Foundation of China (Grant Nos.: 81670555, 81070348, 81200307, and 81170412). It was also funded by the Opening Foundation of Hubei Key Laboratory of Tumor Microenvironment and Immunotherapy (China Three Gorges University) (No. 2016KZL05), the Science Research Innovation Foundation of Graduate Student of China Three Gorges University (No. 2016PY051), and the Science Research Innovation Foundation of Undergraduate Student of China Three Gorges University (No. 2016YCX002).
Authors' Contributions
Y.-Q.Z. and L.-Y.W. contributed equally to perform the experiments and to write the article; X.-M.H., Y.-R.N., and C.W. provided some help for this work; J.-F.W. and C.-B.L. are corresponding authors and guided the research; J.-F.W. and C.-B.L revised and approved the article to be published. All authors read and approved the final article.
Disclosure Statement
No competing financial interests exist.