
Abstract
Acute lung injury (ALI) and acute respiratory distress syndrome (ARDS) are life-threatening inflammatory conditions with no effective pharmacological treatment. Previous studies suggested that mesenchymal stromal/stem cell (MSC) infusion resulted in better survival in mouse ALI models and presented low toxicity in human subjects. Therefore, in this study, we investigated the possibility of treating a murine model of ALI using MSCs with constant interleukin-10 overexpression (IL-10-MSC) by retroviral infection. ALI in mice was induced by intratracheal lipopolysaccharides (LPS) instillation. After 96 h, 80% of mice receiving IL-10-MSCs survived, whereas the survival rate of the mice receiving other treatments was only 20–50%. Mice receiving IL-10-MSCs also demonstrated significantly less weight loss (p < 0.01), and lower protein level and TNF concentration in the BAL (p < 0.01). Interestingly, IL-10-MSCs given to mice 3 and 1 day before ALI induction still conferred significant protection against ALI. While direct IL-10 transfusion resulted in an intensive, but transient peak in serum IL-10 level, IL-10-MSCs provided a milder, but more persistent increase in serum IL-10 level, together with significantly higher levels of IL-10-producing T cells and B cells, both in the spleen and in the lung. IL-10-MSCs given 3 days before LPS challenge resulted in higher pulmonary infiltration of IL-10-producing T cells and B cells in mice. On average, mice that survived the LPS challenge for 96 h presented higher pulmonary infiltration of IL-10-producing T cells and B cells than mice that deceased within the experimental period. Together, these results demonstrated that IL-10-MSCs offered superior protection against LPS-induced ALI when given before or at the time of ALI induction, and significantly increased the frequencies of IL-10-expressing T cells and B cells. IL-10-MSCs may thus represent a promising new treatment option in ALI/ARDS.
Introduction
A
Mesenchymal stromal/stem cells (MSCs) can be derived from the adult bone marrow and possess stem cell characteristics, such as the capacity to self-renew and differentiate into multiple cell types (Kratchmarova et al., 2005). Moreover, in animal models of ALI, MSCs present low immunogenicity and possess the capacity to secrete multiple paracrine factors, such as antimicrobial peptides, endothelial and epithelial growth factors, and anti-inflammatory cytokines (Gupta et al., 2007; Lee et al., 2009; Krasnodembskaya et al., 2010; Mei et al., 2010; Walter et al., 2014), which are potentially useful functions in treating ALI. Pioneering studies have already investigated the use of MSCs in multiple inflammatory disorders, such as sepsis, diabetes, Crohn's disease, and graft-versus-host disease (Parekkadan and Milwid, 2010). Clinical trials are being conducted to examine the possibility of MSC transfusion as an ALI/ARDS therapy. Results from a phase I trial demonstrated that MSC was reasonably well tolerated by patients with moderate to severe ARDS (Wilson et al., 2015). MSCs could also be engineered to express cytokines of interest, with the intention of enhancing their functions. Studies in murine collagen-induced arthritis and graft-versus-host disease demonstrated that IL-10-transduced MSCs were more effective at suppressing pathogenic inflammation in these diseases, with reduced T cell inflammation (Min et al., 2007; Choi et al., 2008).
Based on previous results, we investigated the efficacy and mechanism of IL-10-overexpressing MSCs in treating lipopolysaccharides (LPS)-induced ALI.
Methods
Mice
Male C57BL/6 between 6 and 8 weeks of age were housed in the animal facility of Renji Hospital and used in all experiments. All groups started with 5 or 10 randomly selected mice as specified per experiment, and the experimenters were blinded about the grouping of mice. All experimental procedures were approved by the ethics committee at Renji Hospital.
LPS challenge
Mice were first anesthetized with 150 μg/g ketamine and 13.5 μg/g acepromazine by intraperitoneal injection. ALI was then induced by intratracheal instillation of 5 μg/g LPS from Escherichia coli O55:B5 (Sigma Aldrich), after which mice were placed in 100% oxygen chamber for recovery for 1–2 h. All surviving mice by 96 h were sacrificed by exsanguination from inferior vena cava.
MSC isolation, culturing, and transfection
IL-10-MSCs (green fluorescent protein [GFP]+IL-10+) and control MSCs (GFP+) were produced as previously described (Min et al., 2007; Choi et al., 2008). Briefly, murine bone marrow cells were collected by flushing the femur and tibia with DMEM and were incubated in 75 cm2 flasks at 1 × 106 cells/mL in DMEM supplemented with 10% FBS, 2 mM
MSC injection
1 × 106 MSCs or 5 μg per mouse recombinant murine IL-10 CF (BioLegend) in 50 μL phosphate-buffered saline (PBS) or 50 μL PBS only were injected into anesthetized mice i.t. For injection on day 0 of ALI induction, the mice were rested for 4 h after LPS instillation and then injected with MSCs or PBS. All mice were placed in 100% oxygen chamber for 1–2 h of recovery before returning to the original chamber.
BAL protein analysis
Standard BAL was performed in mice as described previously (D'Alessio et al., 2009; Li et al., 2016). Specifically, a 20-gauge catheter was inserted into the mouse trachea and 0.7 mL sterile PBS were flushed back and forth twice. The aspirate fluid was then centrifuged at 600 g for 8 min at 4°C to separate out the supernatant from the cell pellet. The concentration in the protein was measured using the Modified Lowry Protein Assay kit (Thermo Fisher) following the manufacturer's instructions. The concentration of TNF was measured in BAL using commercial ELISA kits (R&D Systems).
Lung tissue processing
Lungs were excised, minced, and digested in RPMI 1640 (Gibco) supplemented with 2.4 mg/mL collagenase I and 20 μg/mL DNase (Sigma Aldrich) for 1-h digestion in a 37°C water bath with gentle shaking. A single cell suspension was obtained by pushing the digestion product through a cell strainer with 70-μm pores (Corning). The cells were then mixed with stock isotonic percoll to make 23% percoll solution, which was layered on top of 70% percoll and centrifuged at 500 g for 30 min. The lung-infiltrating lymphocytes were harvested at the interphase and washed in the culture medium to remove percoll. Splenocytes were collected by gently smashing the excised spleens through a cell strainer with 7-μm pores, with washing using sterile RPMI 1640 complete medium.
Flow cytometry
Confirmation of MSC identity was performed by surface staining with anti-mouse CD11b, CD31, CD34, and CD45 antibodies (BD) on MSCs between passages 15 to 25, the same passages used to inject mice. Expression of intracellular IL-10 was examined by first surface staining lung lymphocytes with anti-CD3, CD4, CD8, and CD19 antibodies (BD) for 30 min in 4°C. The cells were then washed and treated with BD CytoFix/CytoPerm for 15 min in 4°C. Cells were washed again, strained with anti-mouse IL-10 (BD) for 30 min in 4°C, washed twice, and examined in BD FACSCanto cytometer.
Statistical analysis
The Prism 6 software was used for all statistical analyses. Two-tailed p value less than 0.05 was used as a standard for statistical significance.
Results
IL-10-overexpressing MSCs promoted better survival in LPS-induced ALI
We first produced IL-10-overexpressing GFP-transfected MSCs (IL-10-MSC, GFP+IL-10+) and control GFP-transfected MSCs (control MSC, GFP+) using retroviral infection as previously described (Min et al., 2007; Choi et al., 2008). Following transfection, between 90% and 97% of IL-10-MSCs and control MSCs expressed GFP, and between 88% and 94% IL-10-MSCs expressed intracellular IL-10 (Fig. 1A). The IL-10 secretion in IL-10-MSCs was further confirmed by enzyme linked immunosorbent assay (ELISA), while the treatment-naive uninfected MSCs and control MSCs did not express IL-10 above the detection limit (Fig. 1B). Then, in mice with LPS-induced ALI, 1 × 106 IL-10-MSC, control MSC, or uninfected MSC in 50 μL PBS or 50 μL PBS only without any MSC was injected through intratracheal administration. Mice receiving IL-10-MSC demonstrated significantly improved survival than mice receiving control MSC, uninfected MSC, or PBS (Fig. 1C). The body weight of mice was also examined every 24 h after ALI induction until sacrifice or death. Mice receiving IL-10-MSC had significantly higher body weight than mice receiving control MSC, at 48 and 72 h, but not 96 h, after ALI induction (Fig. 1D). IL-10-MSC-injected, control MSC-injected, and uninfected MSC-injected mice demonstrated less weight loss than PBS-injected mice at 48, 72, and 96 h after ALI induction. Control MSC-injected mice and uninfected MSC-injected mice demonstrated a trend of recuperation at 96 h, while the PBS-injected mice did not.
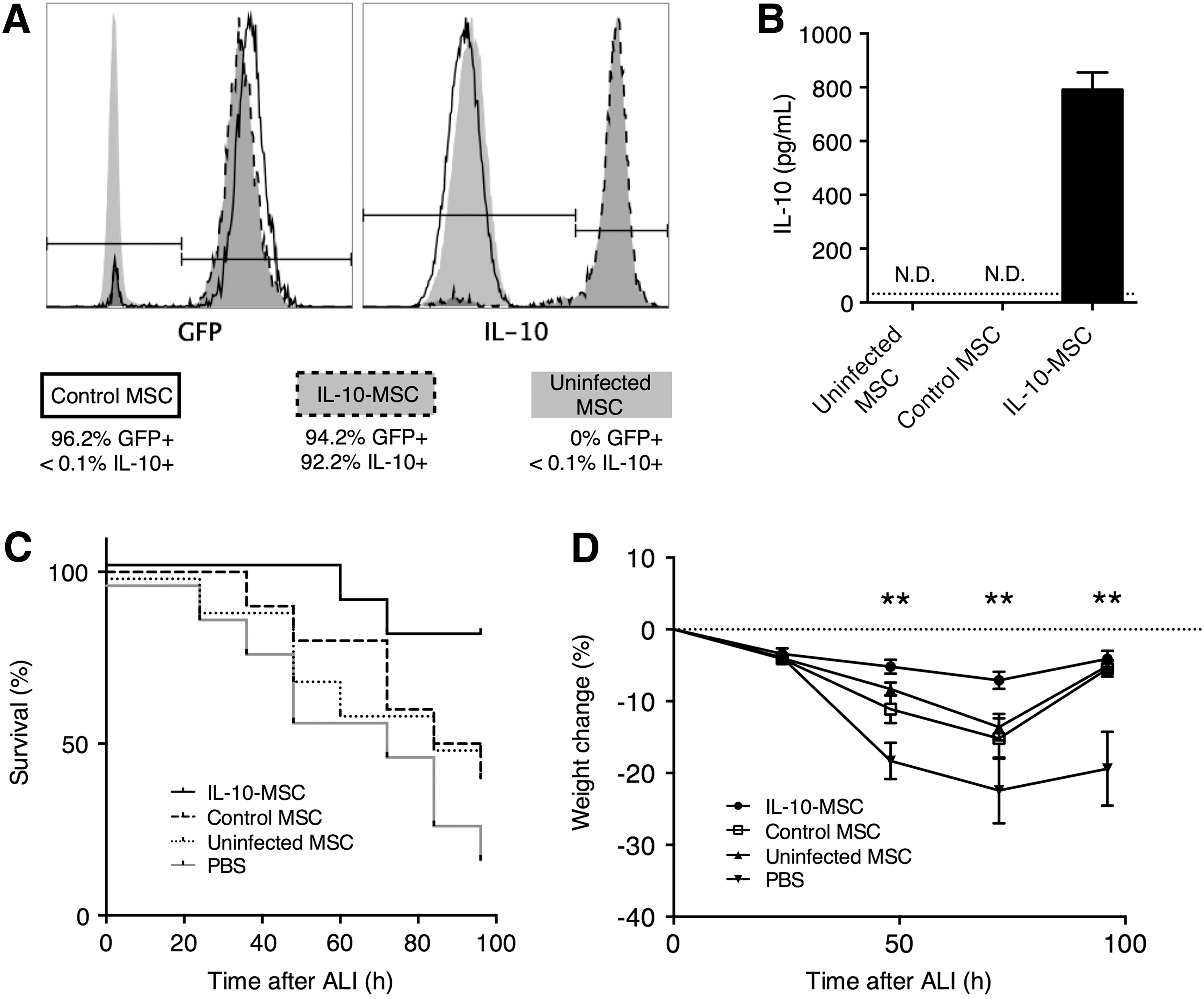
Effect of IL-10-MSC injection in mice with LPS-induced ALI.
The BAL protein level is a measure of endothelial and epithelial permeability and has been used as a marker of ALI severity (Gupta et al., 2007; D'Alessio et al., 2009). At 24 h after ALI induction, no significant differences in BAL protein level were observed among mice receiving IL-10-MSC, control MSC, uninfected MSC, or PBS (Fig. 2A). At 48 and 72 h, mice receiving IL-10-MSC, control MSC, and uninfected MSC presented significantly lower BAL protein level than mice receiving PBS. TNF is a proinflammatory cytokine implicated in ALI pathogenesis (Patel et al., 2013). At 24, 48, and 72 h post-ALI induction, mice receiving IL-10-MSC, control MSC, and uninfected MSC presented significant TNF concentration in the BAL than mice receiving PBS (Fig. 2B).
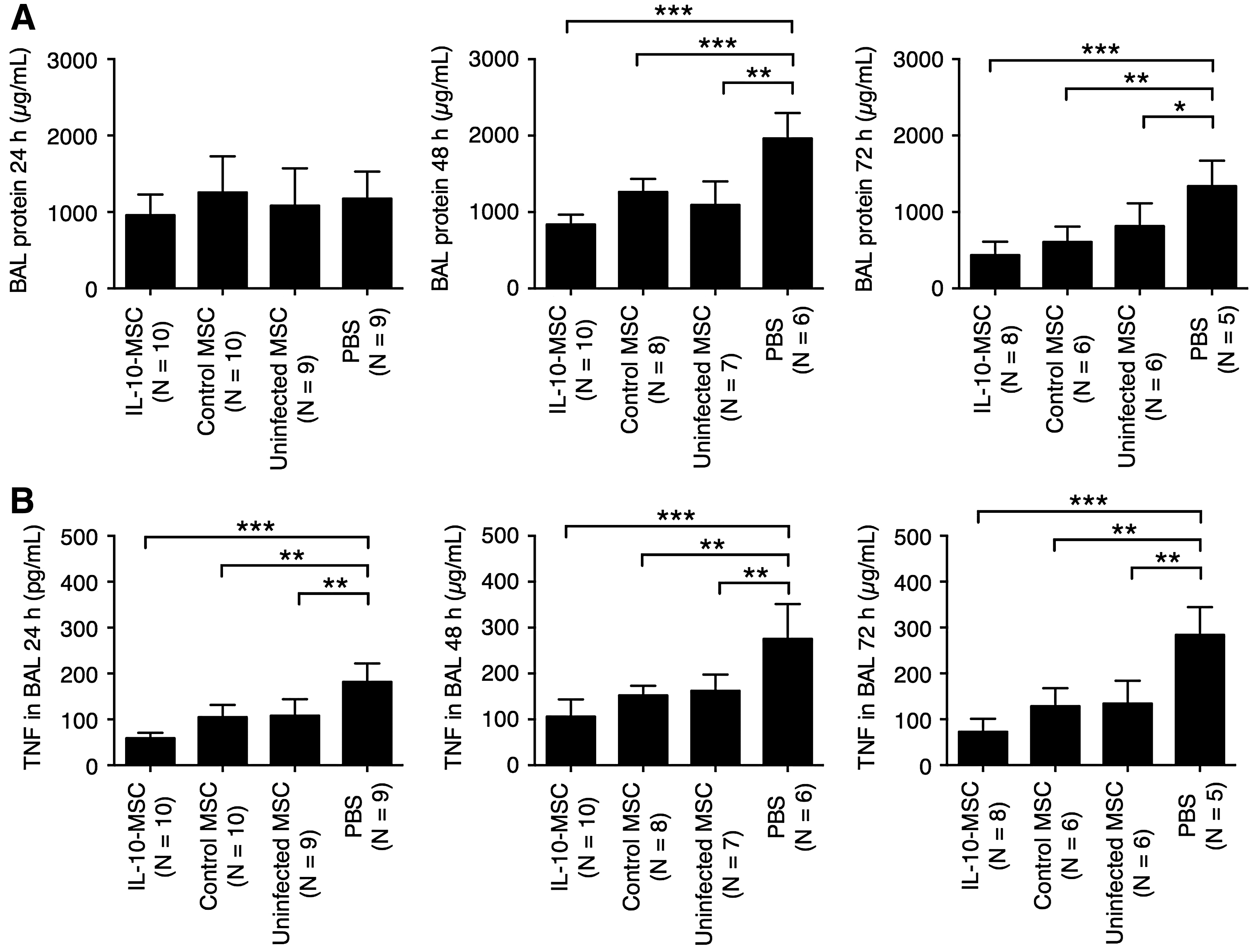
Effect of MSC injection in BAL protein level and TNF concentration.
Protection against LPS-induced ALI could be mediated by IL-10-MSCs injected 3 days before ALI induction
The previous experiments demonstrated that IL-10-MSCs provided potent protection to mice with LPS-induced ALI. To further characterize IL-10-MSC-mediated protection, we injected IL-10-MSC at −3, −1, 0 (time of ALI induction), and 1 day after ALI induction. We found that IL-10-MSC injection before and at the same time of ALI induction resulted in better mouse survival, with 80%, 70%, and 70% survival at 96 h for −3, −1, and 0 day injection, respectively (Fig. 3A). Mice with IL-10-MSC injection 1 day after ALI induction also presented better survival than mice with PBS injection at 96 h (50% survival in IL-10-MSC 1 day after ALI vs. 30% survival in PBS), but at 24 and 36 h after ALI induction, both IL-10-MSC-injected 1 day after ALI and PBS-injected mice presented similar mortality. The BAL protein concentration and TNF concentration in BAL were subsequently examined. At 24 h after ALI induction, mice receiving IL-10-MSC −3 or −1 day after ALI induction demonstrated significantly lower BAL protein level and TNF concentration than mice receiving IL-10-MSC 1 day after ALI or PBS (Fig. 3B, C). At 48 h, mice receiving IL-10-MSC −3, −1, or 0 day after ALI induction demonstrated significantly lower BAL protein level than mice receiving IL-10-MSC 1 day after ALI or PBS. At 72 h, mice receiving IL-10-MSC at every time point demonstrated significantly lower BAL protein level and TNF concentration than mice receiving PBS. Together, these results demonstrated that IL-10-MSCs could offer protection against ALI pathogenesis at the time of ALI induction as well as up to 3 days before ALI induction.
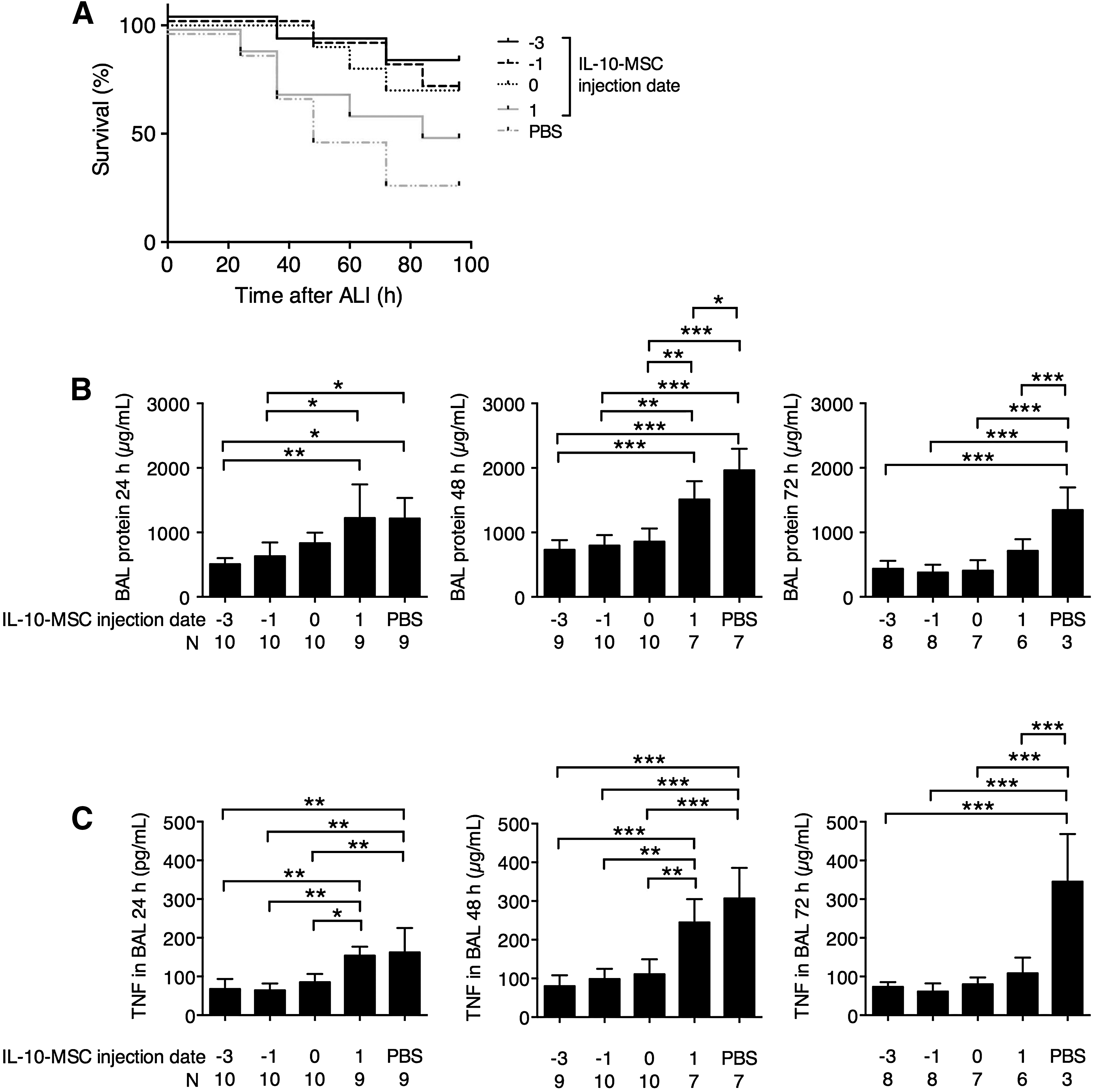
Effect of IL-10-MSC injection at various time points.
IL-10-MSCs resulted in sustained enrichment of serum IL-10 and IL-10-expressing T cells
A number of studies have demonstrated that increased IL-10 expression is protective against aggravated lung injury (Inoue, 2000; Wu et al., 2009; Hiroshima et al., 2014). While direct IL-10 injection resulted in high initial serum IL-10 level, which was diminishing 48 h postinjection, IL-10-MSCs delivered a prolonged increase in serum IL-10 levels throughout the duration of the experiment (Fig. 4A). To explain the increasing trend of serum IL-10 in IL-10-MSC-injected mice, we examined the expression of IL-10 by mouse splenocytes. Although direct IL-10 injection did not significantly increase the frequency of IL-10-producing CD4+ T cells or B cells, IL-10-MSC injection resulted in significantly higher frequencies of IL-10-producing CD4+ T cells and IL-10-producing B cells, both at 48 and 96 h postinjection (Fig. 4B, C). Interestingly, we also observed a group of IL-10-producing CD8+ T cells in splenocytes from mice receiving IL-10-MSCs, whereas in mice receiving IL-10 or PBS, IL-10-producing CD8+ T cells were absent or only observed at very low levels (Fig. 4D). Together, these data demonstrated that IL-10-MSCs, but not direct IL-10, could potently upregulate the frequency of IL-10-producing lymphocytes in mice.
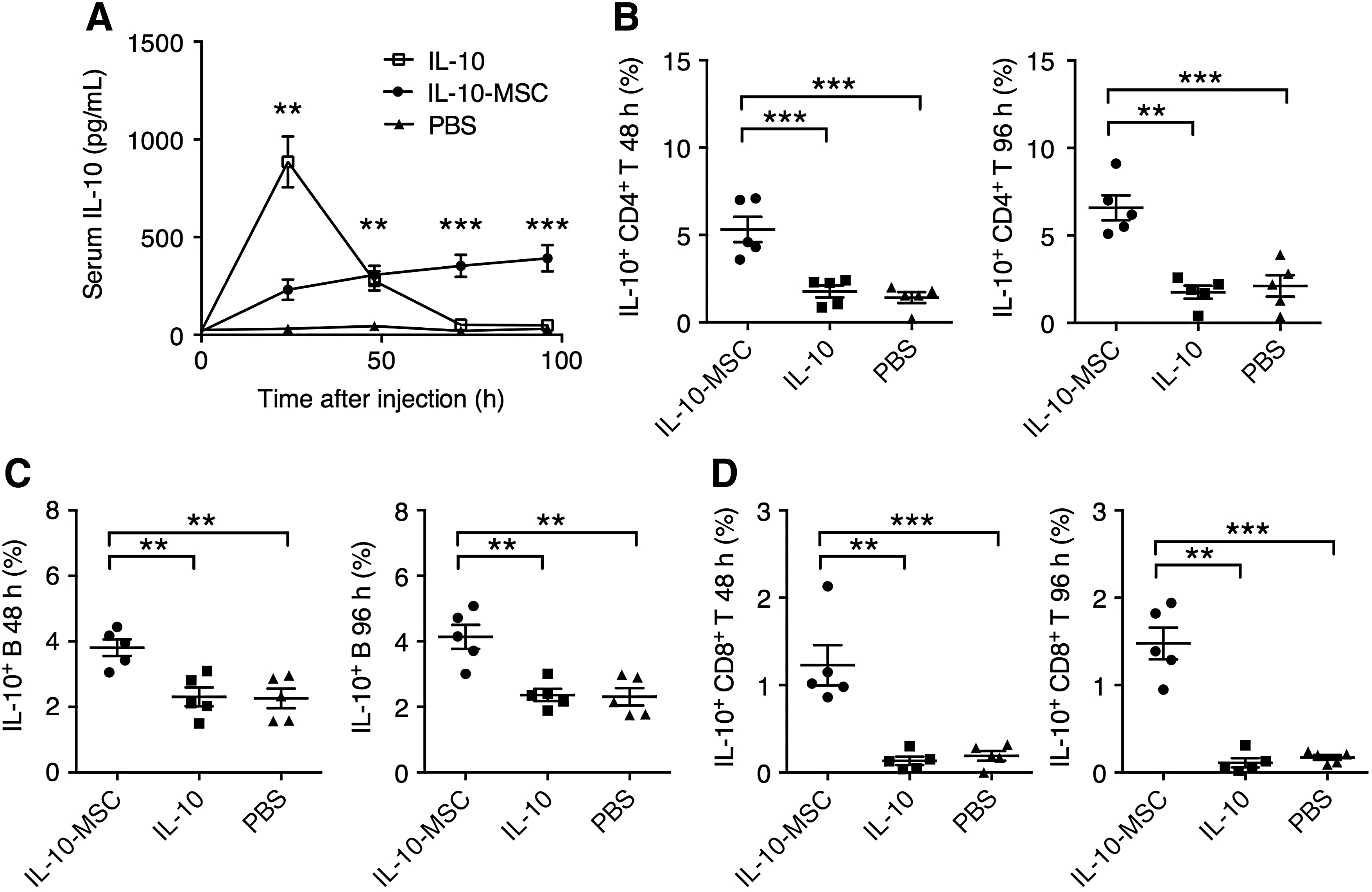
Effect of IL-10-MSC injection compared to direct IL-10 injection in mice.
IL-10-MSCs induced earlier and stronger lung infiltration of IL-10-producing T cells and B cells
To further explain the IL-10-MSC-mediated survival benefit in mice with ALI, we examined the infiltration of IL-10-producing T and B cells in the lung of IL-10-MSC-injected or PBS-injected mice at 0 (no ALI induction), 48, and 96 h after ALI induction. We found that in mice receiving IL-10-MSC 3 days before ALI, the frequencies of IL-10-expressing CD4+ T cells and B cells were significantly higher at baseline (no ALI induction), and the higher frequencies were maintained throughout the experiment. In mice receiving IL-10-MSC on the day of ALI, an increase in IL-10-expressing CD4+ T cells and B cells was observed in the lung at 48 and 96 h after ALI induction, whereas in mice receiving PBS, higher IL-10-expressing CD4+ T cells and B cells were observed only at 96 h (Fig. 5A, B). Interestingly, a rare subset of IL-10-producing CD8+ T cells was discovered in mice receiving IL-10-MSC at −3 and 0 days after ALI at 48 h after ALI induction, while no induction of IL-10-producing CD8+ T cells was observed in mice receiving PBS (Fig. 5C).
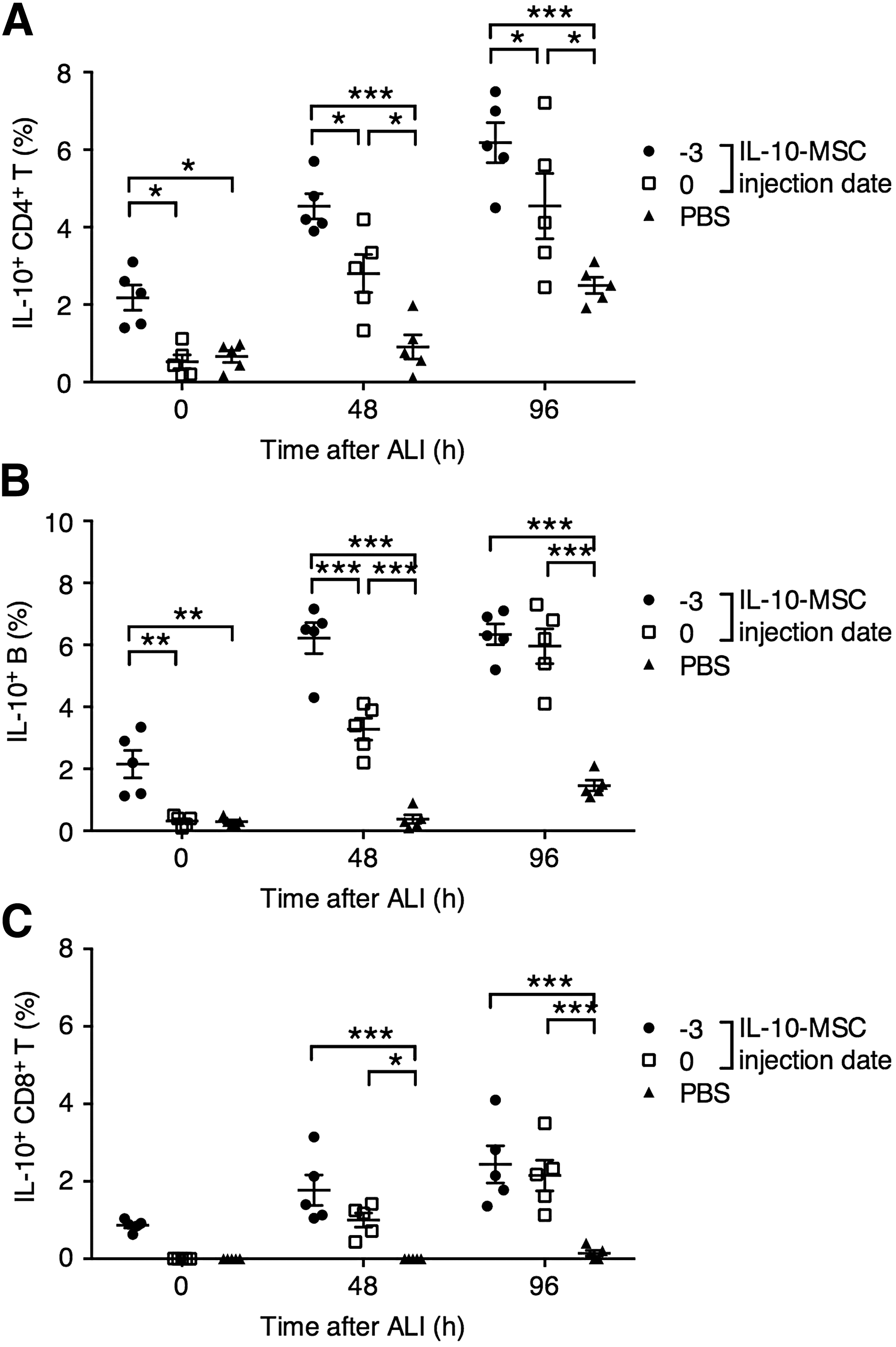
Lung infiltration of IL-10-expressing T cells and B cells in LPS-induced ALI mice receiving IL-10-MSC or PBS.
Mice that survived LPS-induced ALI presented higher pulmonary infiltration of IL-10-producing T cells and B cells
To examine the association between LPS-induced ALI outcome and infiltration of IL-10-producing lymphocytes, we examined the frequencies of IL-10-producing CD4+ T cells, CD8+ T cells, and B cells in mice that deceased within 96 h of ALI induction, or survived for 96 h after ALI induction. The lung tissue from deceased or survived mice in Figures 1C and 3A was pooled and the infiltration of IL-10-secreting lymphocytes was examined. We found that mice that survived ALI demonstrated significantly higher levels of IL-10-producing CD4+ T cells, B cells, and CD8+ T cells (Fig. 6A–C).
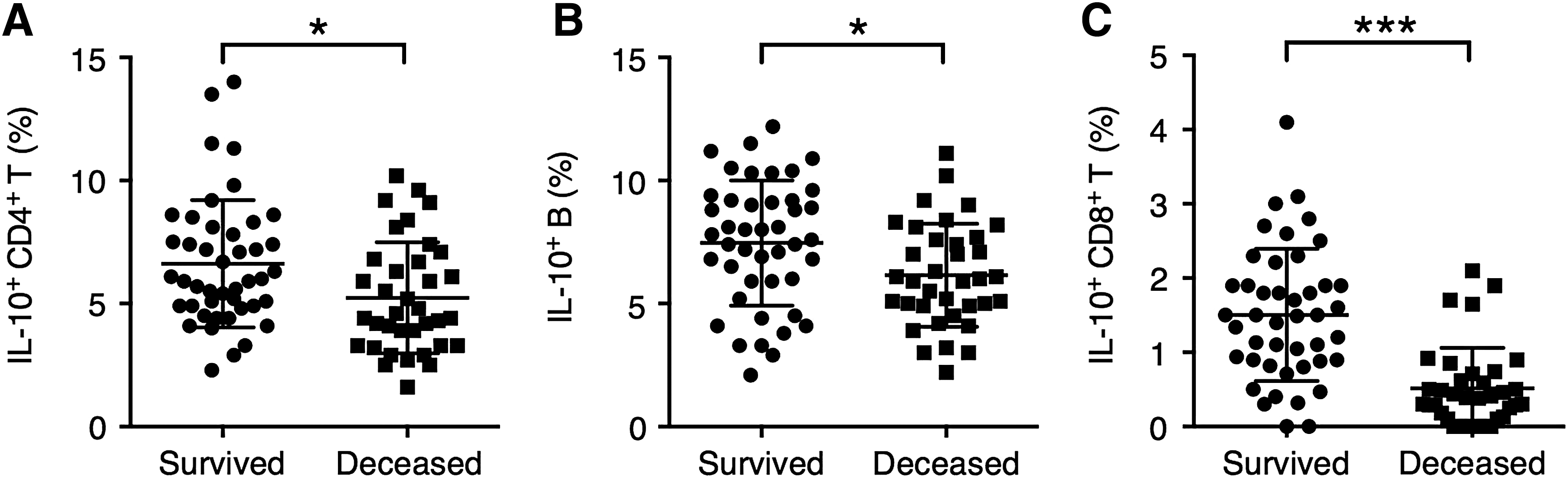
The association between the level of lung infiltration of IL-10-expressing T cells and B cells and the outcome of ALI in mice.
Discussion
Studies have shown that MSC infusion could significantly improve the overall survival of mice with ALI, with reduced level of pulmonary injury (Rojas et al., 2005; Gupta et al., 2007). Two recent trials in small groups of human patients with moderate to severe ARDS demonstrated that MSC infusion did not induce severe infusion toxicity or MSC-related serious adverse effects (Zheng et al., 2014; Wilson et al., 2015). In this study, we demonstrated that IL-10-MSCs were beneficial to mice with LPS-induced ALI. IL-10-MSC-treated mice demonstrated higher survival rate than control MSC-treated, uninfected MSC-treated, and PBS-treated mice. The weight change in IL-10-MSC-treated mice was also lower than control MSC-treated, uninfected MSC-treated, and PBS-treated mice. Moreover, mice receiving IL-10-MSC, control MSC, and uninfected MSC demonstrated lower BAL protein concentration and TNF concentration in stored BAL samples than mice receiving PBS. We subsequently found that IL-10-MSC given 3 or 1 day before ALI induction conferred higher overall survival, lower BAL protein concentration, and lower TNF concentration than mice receiving either IL-10-MSC 1 day after ALI induction or PBS. These results demonstrated that IL-10-MSC could reduce ALI-induced mortality and the severity of inflammation in the BAL after LPS instillation, when given before or on the day of ALI induction. Moreover, IL-10-MSC, control MSC, and uninfected MSC could all reduce inflammation in the BAL after LPS instillation.
Compared to direct IL-10 transfusion, which produced a transient peak in serum IL-10 concentration, we found that IL-10-MSC infusion resulted in persistently high serum IL-10 concentration with an increasing trend over time. Also, previous studies by others demonstrated that certain T cell and B cell subsets were associated with better outcomes in ALI/ARDS and could potentially mediate protective effects in ALI/ARDS, in part, through IL-10 (D'Alessio et al., 2009; Xu et al., 2016; Kapur et al., 2017). Therefore, to explain the increasing trend in serum IL-10 level after IL-10-MSC treatment, we examined the IL-10 production by T cells and B cells in mice receiving IL-10-MSCs. Interestingly, we found that IL-10-MSC infusion significantly elevated the frequency of IL-10-producing CD4+ T cells, B cells, and CD8+ T cells, both in the spleen and the lung. Mice that survived the LPS challenge for 96 h presented higher pulmonary frequencies of these IL-10-producing cells than mice that deceased within this period. These results suggested that in mice infused with IL-10-MSCs, but not IL-10, other immune cells, such as splenic and pulmonary T cells and B cells, could also act as a source of IL-10.
This study has several limitations that need to be resolved in future investigations. First, BAL was only harvested in surviving mice at each time point. Therefore, the measured BAL protein levels and TNF concentrations likely skewed toward the lower end, since the surviving mice likely presented less severe inflammation than deceased mice. This was especially a problem in the control MSC-treated, uninfected MSC-treated, and PBS-treated groups because these groups had higher mortality than the IL-10-MSC-treated group. Also, we demonstrated that IL-10-MSC infusion could result in higher frequencies of IL-10-producing T cells and B cells, but the specific mechanisms involved in this change in T cell and B cell responses are unclear. Possibly, the high IL-10 concentration stimulated antigen-presenting cells, such as macrophages and dendritic cells, to differentiate into a regulatory type (Martinez and Gordon, 2014); these antigen-presenting cells could then promote the development of Treg and B cell subsets. Whether this hypothesis is true in the IL-10-MSC-treated mice requires further examinations. Also, the effect of IL-10-MSC treatment on the innate immune system, especially in pulmonary macrophages, is yet unclear and requires further examinations. In addition, we found that IL-10-MSC infusion resulted in a systemic increase of IL-10-producing T cells and B cells. These cells could suppress immune responses in organs not involved in ALI, and the potential effects of this systemic alteration of the immune status should be investigated further. In addition, this study was performed in a direct lung injury model with intratracheal instillation of LPS. In human ALI and ARDS, ∼20% of cases arose as a secondary phenomenon after other causes, such as acute pancreatitis, sepsis, burn injury, and other traumatic events, and a further 21% of cases arose from mixed factors (Perl et al., 2011). The efficacy of IL-10-MSC infusion in indirect ALI is unclear and should be examined in further studies.
In conclusion, there are three major findings in our study. First, compared to control MSCs and uninfected MSCs, IL-10-MSCs were superior in treating mice with LPS-induced ALI. Second, we showed that IL-10-MSCs could offer protection against LPS-induced ALI when transfused before the ALI induction. And third, we found that IL-10-MSCs resulted in significant changes in the frequency and pulmonary infiltration of IL-10-producing T cells and B cells.
Footnotes
Disclosure Statement
No competing financial interests exist.