
Abstract
Chronic prostatitis (CP) with complex pathogenesis is difficult for treatment. c-kit has been associated with the control of cell proliferation of prostate cells. This study aims to evaluate the role of resveratrol, an activator of Sirt1, in regulating the expression of c-kit in CP and investigate the consequent effects on cell cycle. Rat model of CP was established through subcutaneous injections of diphtheria-pertussis-tetanus vaccine and subsequently treated with resveratrol. Hematoxylin and eosin staining was performed to identify the histopathological changes in prostates. Western blotting and immunohistochemical staining examined the expression level of c-kit, stem cell factor (SCF), Sirt1, and cell cycle-associated proteins. The model group exhibited severe diffuse chronic inflammation, characterized by leukocyte infiltration and papillary frond protrusion into the gland cavities, and a notable increase in prostatic epithelial height. Gland lumen diameter was also significantly smaller; the activity of c-kit/SCF in the CP rats was increased significantly compared to the control group. Meanwhile, the cell cycle proteins are dysregulated significantly in CP rats. Resveratrol treatment significantly improved these factors by Sirt1 activation. Dysregulation of cell cycle was involved in the pathological processes of CP, which was improved after resveratrol treatment by the downregulation of c-kit/SCF by activating Sirt1.
Introduction
C
As an important member of tyrosine kinase family, c-kit receptor causes specific expression of certain genes, regulates cell differentiation and proliferation, resists cell apoptosis (Lennartsson and Ronnstrand, 2012; Liang et al., 2013), and plays a key role in cellular functions through activating the downstream signaling molecules following interaction with stem cell factor (SCF) (Ronnstrand, 2004). The abnormality of SCF/c-kit signaling pathway is closely related to some disease (Boissan et al., 2000; Hollingsworth et al., 2016). SCF and c-kit are expressed in a broad range of male reproductive tissues, including the prostate (Cardoso et al., 2014). As reported (Figueira et al., 2016), 17β-estradiol treatment decreased the expression of SCF and c-kit both in human prostate cells and rat prostate, with the consequent effects that the diminished expression of SCF/c-kit was underpinned by the reduced proliferation index and augmented apoptosis, and their role in prostate carcinogenesis also has been indicated (Wiesner et al., 2008). However, there is no report on the role of c-kit/SCF in the development of CP.
Currently, effective therapy of CP remains in the exploratory stage. Resveratrol (Torres et al., 2010) (trans-3, 4, 5-trihydroxy stilbene) is widely found in many plants and is well known as Sirt1 activator. The reliable and extensive anti-inflammatory effects of resveratrol aroused the attention of researchers (Csiszar, 2011). Interestingly, resveratrol produced an antiproliferative effect by regulating the expression levels of proteins involved in inflammation and apoptosis during benign prostatic enlargement (Chung et al., 2015). Also, resveratrol has gained enormous attention in other prostate diseases (Maciej, 2013). Therefore, resveratrol may have a potential role in improving CP.
Therefore, this study was performed to evaluate the role of SCF/c-kit in the development and progression of CP in rats and to investigate the improved effect of resveratrol. The consequent effect of Sirt1 stimulation on cell cycle of prostate cells in vivo was also investigated. The obtained findings will contribute to elucidating the molecular mechanisms underpinning the role of cell cycle in the prostate and represent valuable information to define safety and efficacy of resveratrol-based therapies against CP.
Materials and Methods
Chemicals
Antibodies used for Western blot analyses or immunohistochemical staining were as follows: rabbit anti-c-kit, SCF, Sirt1, p18, p21, p27, cyclin D1, cyclin D3, cyclin-dependent kinases (CDK2, CDK4, and CDK6), phospho-retinoblastoma (p-RB, dilution 1:1000; Cruz Biotechnology, Santa Cruz, CA), and rabbit anti-β-actin (dilution 1:1500; Beyotime, China). Cy3-conjugated Affinipure Rabbit Anti-Goat IgG (H+L, dilution 1:100; Cruz Biotechnology, Santa Cruz, CA).
Resveratrol of >99% purity was purchased from Dalian Meilun Biotechnology Co., Ltd. (Dalian, China). DPT vaccine was obtained from the Wuhan Institute of Biological Products Co., Ltd. (Wuhan, China). All other chemicals used in this study were of analytical grade and commercially available.
Animals and ethics statement
Male Sprague-Dawley rats (180 ± 20 g) were purchased from the Experimental Animal Center of Dalian Medical University (Dalian, China). Rats were allowed access to water and food ad libitum, but were fasted overnight before surgery, with water available.
Ethical approval
The ethics committee of Dalian Medical University approved all animal experiments, which were performed in accordance with institutional guidelines. All animal experiments comply with the ARRIVE guidelines and are carried out in accordance with the U.K. Animals (Scientific Procedures) Act, 1986, and associated guidelines, EU Directive 2010/63/EU for animal experiments, and the National Institutes of Health guide for the care and use of Laboratory animals (revised 1978; NIH Publications No. 8023).
Rat model of CP
For the purification of rat prostatic protein, male Sprague-Dawley rats (240–300 g) were sacrificed and prostate tissue was removed under sterile conditions and washed with saline solution. Prostate tissue was placed into a physiological saline solution containing 0.5% Triton X-100 and homogenized in an ice-water bath with a glass homogenizer. The homogenized liquids were centrifuged (10,000 g) for 10 min, and protein was diluted to 15 mg/mL with phosphate-buffered saline (PBS) (0.1 mol/L and pH7. 2).
Rats were subcutaneously injected with a DPT vaccine (0.5 mL/kg) and a multipoint injection of a mixture (1.0 mL) of purified rat prostatic protein and Freund's Complete Adjuvant (1:1) at 0, 15, and 30 days (22). The rat model of CP was established after 45 days. Rats were randomly divided into three groups (eight rats in each group; n = 8): 1. Control group: normal rats orally administered saline for 10 days; 2. CP group: CP rats orally administered saline for 10 days; 3. Resveratrol group: CP rats orally administered resveratrol (10 mg/kg) for 10 days (Cheng et al., 2014).
Morphological changes
The rats were sacrificed at the end of study. The prostate was removed, fixed in 10% (v/v) neutral formalin, processed using standard histological techniques, stained with hematoxylin and eosin (H&E), and examined for morphological changes. The samples were used to determinate the protein expression of c-kit, SCF, Sirt1, cell cycle-associated proteins using western blot analysis, immunohistochemical staining, and immunofluorescence labeling.
Western blot analysis
Proteins were extracted from rat prostate using a protein extraction kit (KeyGen Biotech, Nanjing, China) according to the manufacturer's instructions. Protein was measured according to the bicinchoninic acid procedure (Solarbio, Beijing, China), with bovine serum albumin as the standard. Proteins (20 μg) were resuspended in electrophoresis sample buffer containing β-mercaptoethanol and separated using electrophoresis on a precast 10% SDS-polyacrylamide gel (Bio-Rad, Hercules, CA), followed by electrotransfer to a polyvinylidene fluoride membrane (Millipore, Bedford, MA). Membranes were blocked using 5% nonfat milk in Tris-buffered saline with 0.1% Tween-20 (TBST) for 2 h at 37°C. β-Actin served as the loading control. Membranes were incubated overnight at 4°C with a 1:1000 dilution of polyclonal antibodies for c-kit, SCF, p18, p21, p27, cyclin D1, cyclin D3, CDK2, CDK4, CDK6, and p-RB (Cruz Biotechnology, Santa Cruz, CA), and a 1:1,500 dilution of a monoclonal antibody for β-actin (Beyotime, China). Membranes were subsequently washed with TBST, and the blots were incubated with secondary antibodies. Blots were extensively washed with TBST and exposed to enhanced chemiluminescence-plus reagents from Beyotime Institute of Biotechnology (Haimen, China) according to the manufacturer's protocol. Emitted light was documented using a BioSpectrum-410 multispectral imaging system with a Chemi HR camera 410 (Bio-Rad, Hercules, CA). Protein bands were visualized and photographed under transmitted ultraviolet light. The image was used for semiquantitative measurements based on band densitometry.
Immunohistochemical staining
Histological sections of rat prostate (4 μm thick) were mounted on poly-L-lysine-coated slides. Slides were deparaffinized in xylene and rehydrated in graded alcohols. Sections were pretreated with citrate buffer (0.01 mol/L citric acid, pH 6.0) for 20 min at 95°C and immersed in PBS containing 3% H2O2 for 10 min at room temperature. Sections were exposed to 10% normal goat serum in PBS for 30 min at room temperature and incubated at 4°C overnight with rabbit polyclonal anti-c-kit antibodies (1:100 dilution). Sections were rinsed with PBS, incubated with biotinylated goat anti-rabbit IgG for 20 min at room temperature, and treated with 3,3-diaminobenzidine chromogen for 5 min at room temperature. Sections were counterstained with hematoxylin for 2 min.
Immunofluorescence labeling
Slides with rat prostate sections (4 μm thick) were rinsed with PBS, permeabilized with 0.1% Triton X-100 in PBS for 10 min, and blocked with 2% Bull Serum Albumin in PBS for 30 min. The specimen slides were incubated with a primary anti-c-kit antibody at 4°C overnight. The specimens were washed thrice with PBS and incubated with Cy3-conjugated Affinipure Rabbit Anti-Goat IgG (H + L, dilution 1:100; Cruz Biotechnology, Santa Cruz, CA;) at 37°C for 1 h. Slides were washed and the immunofluorescent images were captured using an 80i Nikon microscope (Tokyo, Japan).
Data analysis
Statistical analysis was computed using SPSS 13.0 software. Group data are expressed as mean ± standard deviation (SD). Statistically significant differences between two data sets were compared using one-way analysis of variance. A p-value of p < 0.05 or p < 0.01 indicated a statistically significant result in all statistical analyses.
Results
Resveratrol improved the pathological changes in CP rats
To demonstrate the effect of resveratrol on CP, morphological changes of prostates were examined by H&E staining. Compared with the control group, the model group exhibited severe diffuse chronic inflammation, characterized by papillary frond protrusion into the gland cavities, and a notable increase in prostatic epithelial height. Gland lumen diameter was also significantly smaller in the lateral lobe of the prostate. However, these changes were significantly suppressed in the rats treated with resveratrol (Fig. 1).
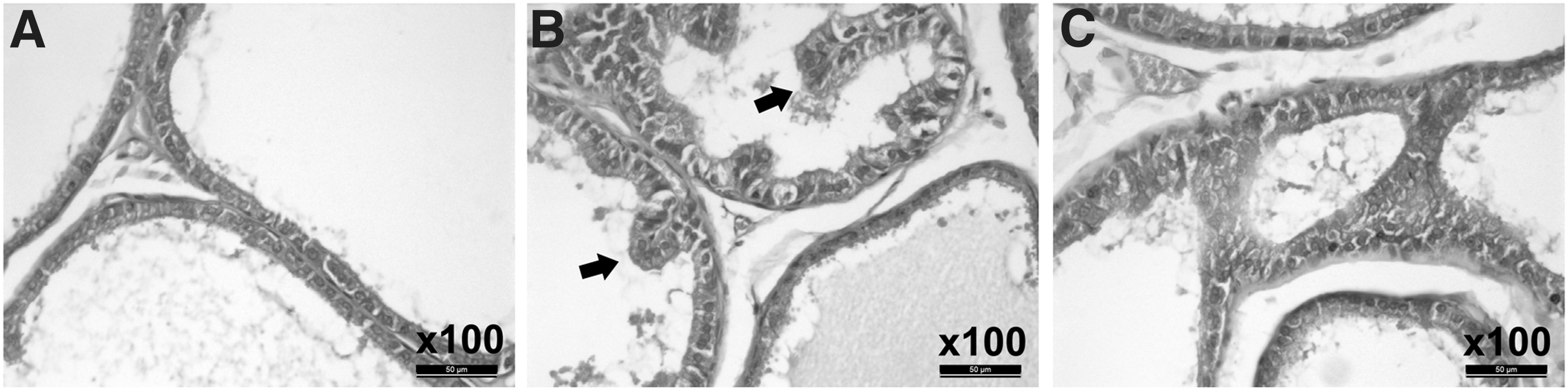
Changes in prostate photomicrographs of CP rats. The prostates are stained with H&E
Resveratrol downregulated the expression of SCF and c-kit proteins by Sirt1 activation
C-kit/SCF has been associated with the control of prostate cell proliferation/apoptosis and prostate carcinogenesis, although in the case of CP this remains unknown. Western blots showed that the expression of SCF and c-kit proteins in the prostate of the CP group increased significantly compared to the control group, while there was no significant difference in Sirt1 expression level between two groups (Fig. 2). These results indicate that the pathogenesis of CP may be associated with the upregulation of c-kit/SCF, independent of Sirt1. However, resveratrol treatment in CP rats significantly decreased the expression of SCF and c-kit protein and increased the Sirt1 expression in the prostate of the CP group (Fig. 2). These results reasoned us to speculate that the inhibition effect of resveratrol on c-kit/SCF was medicated by Sirt1.
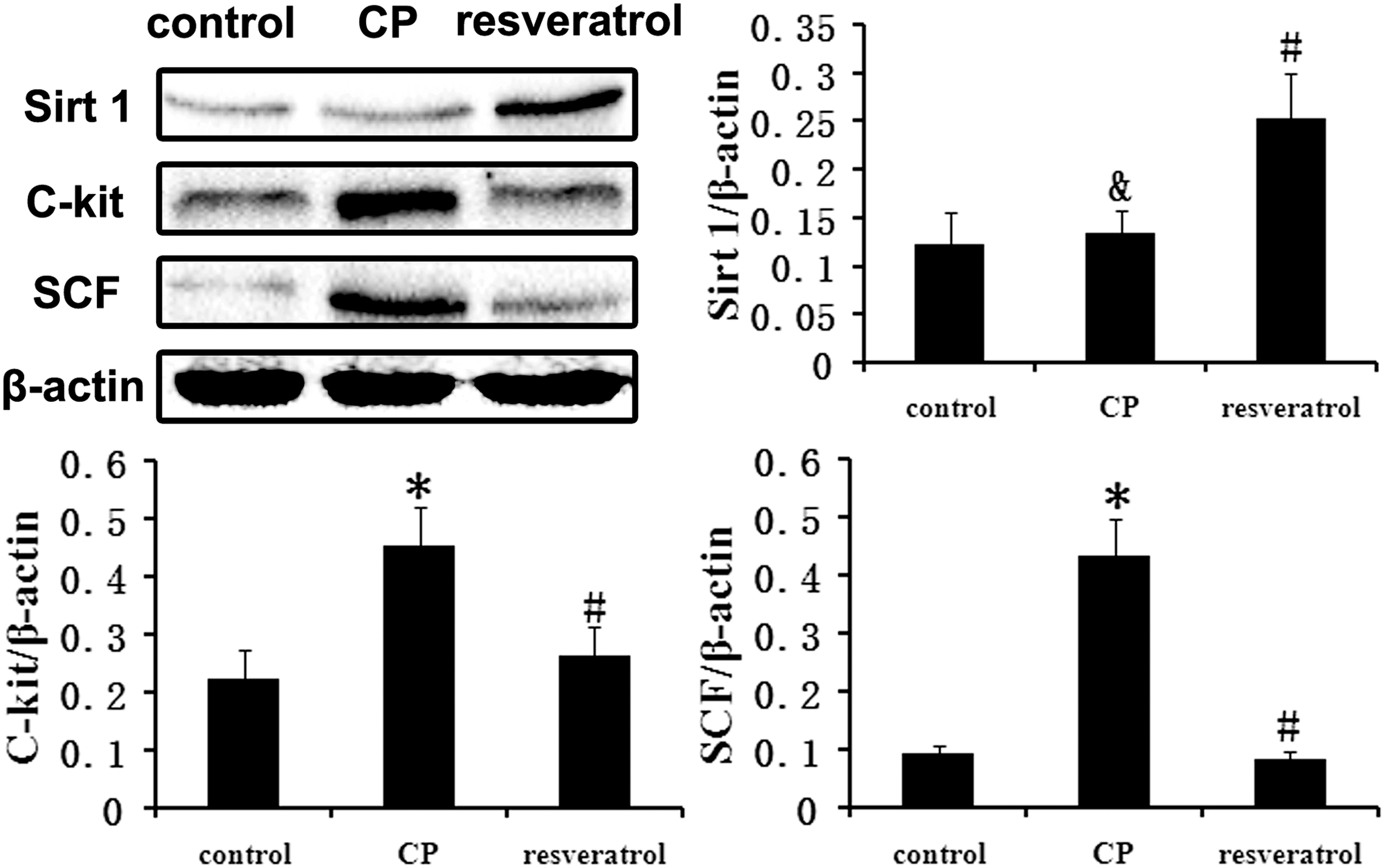
Effect of resveratrol on the expression levels of Sirt1, SCF, and c-kit. Compared with the normal group, the c-kit and SCF were significantly upregulated in the CP group. However, the upregulation was significantly suppressed in the rats treated with resveratrol by increased Sirt1. The bar graph shows the relative expression ratio of each protein calculated after normalization to β-actin. Data are expressed as mean ± SD. (&, p > 0.05 vs. control group *, p < 0.01 vs. control group; #&, p < 0.01 vs. CP group; n = 8). SCF, stem cell factor; SD, standard deviation.
Immunohistochemistry and immunofluorescence results were similar to the above-listed results. The expression levels of c-kit protein in the prostates from the CP group were significantly increased compared to the control group. However, resveratrol treatment of CP rats significantly decreased the expression levels of c-kit protein compared to the CP group (Fig. 3).
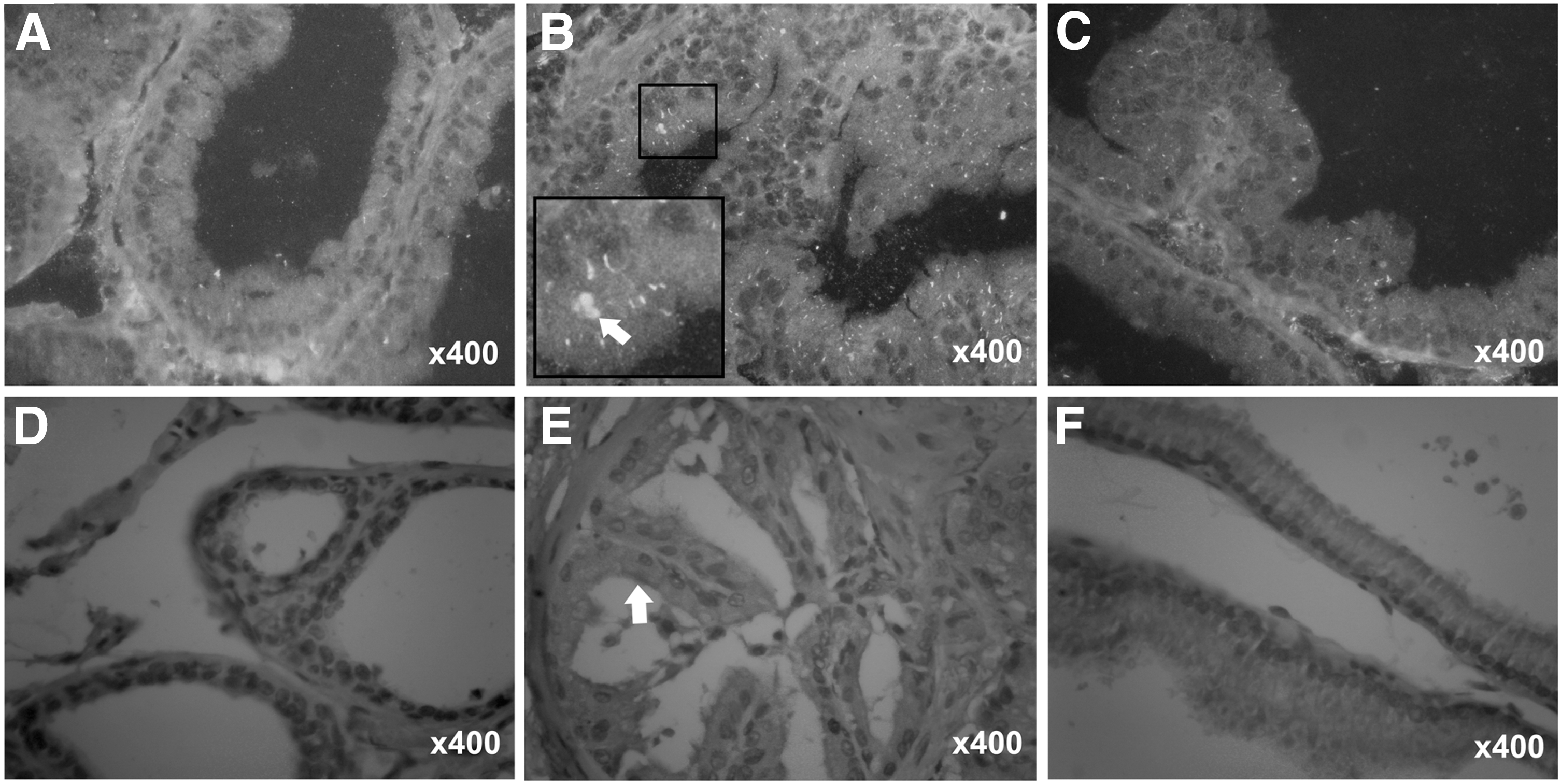
Effect of resveratrol on the expression of c-kit protein in the prostates of CP rats. The prostates were immunofluorescence labeled
Resveratrol improved the dysregulated cell cycle in CP rats
Dysregulation of cell cycle is usually involved in the pathological processes of many complex diseases. To investigate the role of cell cycle in CP pathogenesis, cell cycle proteins, including cyclins, CDKs, and their inhibitors CDKIs, were evaluated. Western blot results showed that the expression level in CP group of p-RB, CDK2, CDK4, CDK6, and cyclin D1 and p21 were significantly decreased, while cyclin D3, p18, and p27 were significantly increased compared with the control group. These results revealed that CP may induce prostate cells to arrest in the G0/G1 phase, and decreased the G1-S phase transition. However, these changes were significantly attenuated in the rats treated with resveratrol (Fig. 4).
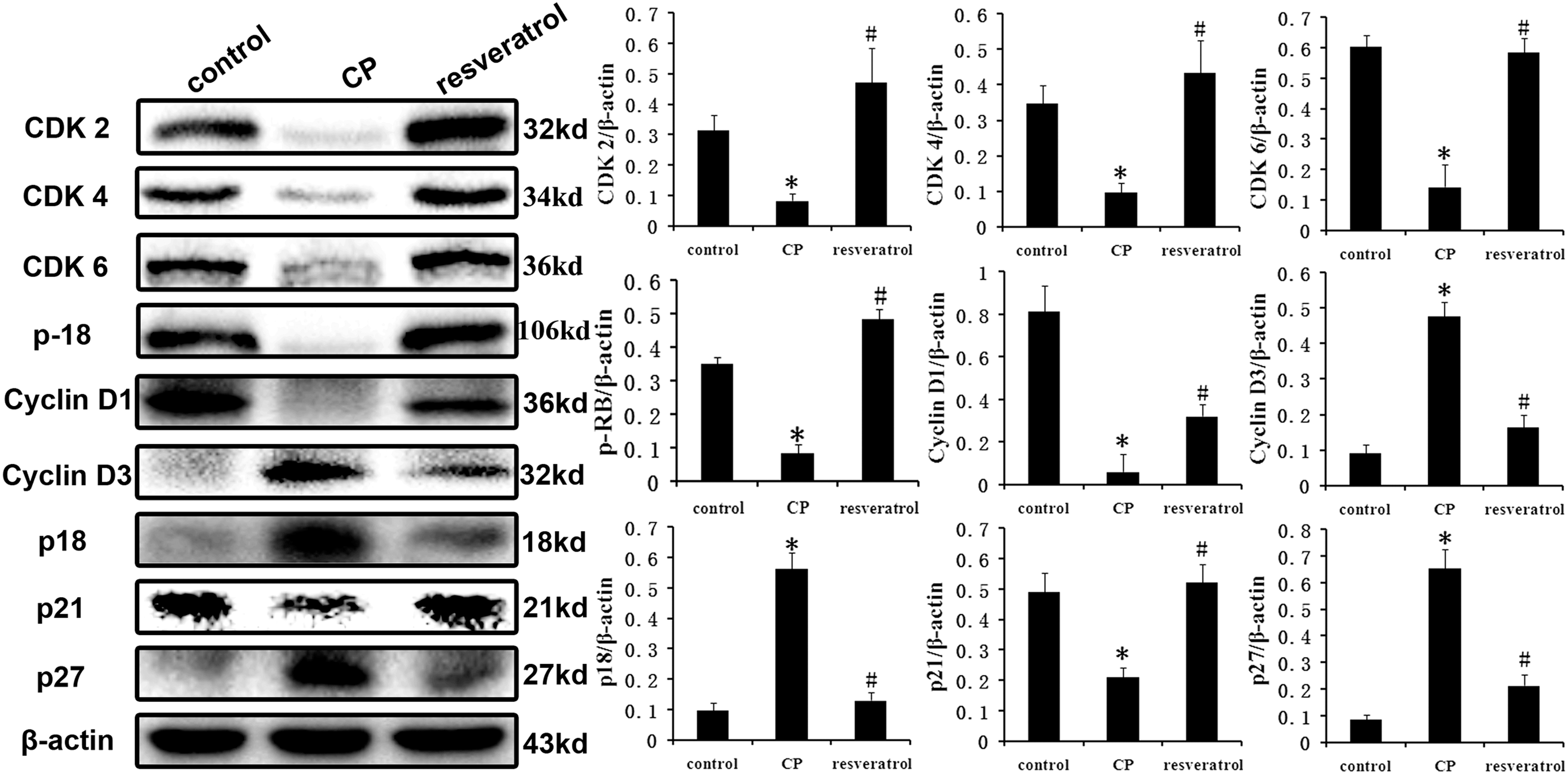
Effect of resveratrol on the protein expression of cell cycle proteins in the prostates of CP rats. Compared with the normal group, their expressions were all significantly dysregulated in the CP group. However, the dysregulation was significantly improved by resveratrol treatment. The bar graph shows the relative expression ratio of each protein calculated after normalization to β-actin. Data are expressed as mean ± SD. (* p < 0.01 vs. control group; # p < 0.01 vs. CP group; n = 8). CDK, cyclin-dependent kinases.
Discussion
The mammalian cell cycle is a highly organized and regulated process that ensures duplication of genetic material and cell division. This regulation involves growth-regulatory signals as well as signals from proteins that monitor the genetic integrity to ascertain the absence of any genetic damage. The cell cycle is governed by a family of cyclins, CDKs (like CDK2, CDK4, and CDK6) and CDKIs (like p18, p21, and p27) through activating and inactivating phosphorylation events (Carbone et al., 2002). Alteration in molecular mechanisms involved in cycle signaling contributes to a vast range of diseases (Gahan, 2001). This study was performed to evaluate the role of some cell cycle regulatory genes in the development and progression of CP. The results showed a significant decrease or loss in CDK2, CDK4, CDK6, and cyclin D1 and p21 protein expression in CP compared to normal controls, while cyclin D3, p18, and p27 were significantly increased. Different classes of cyclins and their complex formation with CDKs control the different phases of cell cycle progression (Giacinti et al., 2006).
G0/G1 phase provides the materials for mitosis, and DNA replication occurs during S phase. The G1 phase checkpoint is regulated by association between CDK 4/6, and cyclins D1 and D3 (Bockstaele et al., 2009; Aggarwal et al., 2010). The formation of cyclin D1-CDK4/6 complexes regulates the synthesis of DNA to prepare for cell division. In CP rats, protein levels of G1-S phase regulators CDK 4 and 6 and cyclin D1 were significantly decreased. Increased cyclin D3 may compensate for the loss of cyclin D1 (Qian et al., 2011). These results (Fig. 2B) revealed that CP induced prostate cells to arrest in the G0/G1 phase.
Similar to the results from G1-phase, CP played suppression effects on S phase checkpoint in the prostate. The association of cyclin E with CDK2 is active at the G1/S transition and directs entry into S phase. During the G1-S phase transition, cyclin E binds to CDK-2, and thus phosphorylates Rb, resulting in the activation of transcription factors and S-phase proteins. S-phase progression is directed by the cyclin A/CDK2 complex (Schwartz et al., 2005.). To understand the effects of CP on the S-phase in prostate, CDK-2 and p-RB levels were estimated. As noted in the G1 phase checkpoint, CP downregulated p-RB and CDK-2 in the prostate in a similar pattern.
Furthermore, we detected the effect of CP on CDKIs p18, p21, and p27 (Roy et al., 2009; Perisanidis et al., 2012.). The p27 negatively regulates cyclin E/CDK2 and cyclin A/CDK2 complexes, and is a predominant CDKI. There are other CDKIs such as p18 and p27 that inhibit CDK-4/-6 and CDK-2, respectively. In CP rats, expression levels of p18 and p21 were increased, but p21 was decreased. Collectively, these data suggest that p18 and p27 could play major roles in the CDK inhibition in the progression of CP.
The mechanism of development of the CP is largely unknown. The SCF/c-kit system plays a relevant role in cell fate decision, and its activity is widely known to control cell proliferation, differentiation, survival, and apoptosis (Lennartsson and Ronnstrand, 2012; Liang et al., 2013). As reported, the c-kit is required in cell cycle regulation (Arun et al., 2008; Kathrina et al., 2013). Cycle arrest is always induced by c-kit supression. In this study, to investigate the molecular mechanism by which the cell cycle dysregulation in CP was dependent on, the c-kit/SCF signaling pathway was investigated. Interestingly, the results showed that the expression of SCF and c-kit proteins in the prostate of the CP group increased significantly compared to the control group, which suggested that the c-kit/SCF pathway was activated during CP progression, so as to induce prostate cells to arrest in the G0/G1 phase.
In this study, we determine the effect of resveratrol on CP. Also, the results of histological examinations showed that resveratrol significantly improved the morphological changes of prostates in CP rats. CP increased the expression of c-kit and SCF in prostate tissues, but had no effect on Sirt1 expression level. After treatment with resveratrol, the c-kit/SCF was decreased and cell cycle arrest was improved. Resveratrol is always known as the activator of Sirt1, which was indeed increased after resveratrol treatment in CP rats. These reasoned us to hypothesize that resveratrol improves cell cycle arrest in CP rats by c-kit/SCF suppression, which was dependent on Sirt1 activation.
Conclusions
In conclusion, the results in this study suggested that dysregulation of cell cycle was involved in the pathological processes of CP, which may be associated with the upregulation of c-kit/SCF, independent of Sirt1. Resveratrol treatment improved cell cycle arrest and suppressed the c-kit/SCF by activating Sirt1, thereby improving CP. Therefore, resveratrol should be considered a candidate for the treatment of CP. Sirt1 and SCF/c-kit are not only important for the cell cycle regulation in prostate but also a possible target for therapy of CP.
Footnotes
Acknowledgment
This work is technically supported by the College of Pharmacy of Dalian Medical University and the Second Affiliated Hospital of Dalian Medical University, Dalian, Liaoning, China.
This work was supported by the Natural Science Foundation of Liaoning province under grant No. (201601229).
Disclosure Statement
No competing financial interests exist.