
Abstract
Excision repair cross-complementation (ERCC) enzymes are key members of the nucleotide excision repair pathway. Dysregulation of ERCC family members has been shown to be involved in chemoresistance in several malignancies. However, the function of ERCC6 in regulating chemo response has not been evaluated in colorectal cancer (CRC). We stably knocked down ERCC6 expression using short hairpin RNA (shRNA) in HCT116 and DLD1 human colon cancer cell lines, followed by chemosensitivity assay. In vivo chemosensitizing effects of ERCC6 were examined in xenograft experiments. Downregulation of ERCC6 conferred sensitivity to 5-fluorouracil (5-FU) in HCT116 and DLD1 cells. Stable knockdown of ERCC6 significantly enhanced antitumor activity of 5-FU in HCT116 xenograft mouse model. ERCC6 was upregulated in CRC tissues compared to matched noncancerous adjacent tissues and was also upregulated in patients who were resistant to 5-FU treatment. In addition, high expression of ERCC6 was associated with poor overall survival in CRC patients with or without receiving 5-FU therapy. Elevated expression of ERCC6 contributes to chemoresistance in CRC cells. Low ERCC6 expression is associated with better chemo response and survival in CRC patients. Therefore, this protein represents a novel therapeutic target for improvement of chemotherapeutic efficacy and predictive biomarker for patient survival.
Introduction
C
Protein members of nucleotide excision repair (NER) process are related to responses to platinum-based chemotherapy. Excision repair cross-complementation group 1 enzyme (ERCC1), which is the major component of the NER pathway, plays a central role in the removal of platinum-induced DNA adducts (Sancar 1994). Numerous studies suggest that there is a relationship between ERCC1 expression and chemotherapy response in various cancer types, including ovarian, lung, cervical cancer, and others (Dabholkar et al., 1994; Bellmunt et al., 2007; Chen et al., 2010; Hoffmann et al., 2010; Park et al., 2011). Although being a close family member to ERCC1, ERCC6 is less well studied. To our knowledge, experimental or clinical data regarding the role of ERCC6 in chemoresistance are limited.
In the current study, we investigated the involvement of ERCC6 in regulating chemoresistance in CRC using both cell-based assays and xenograft mouse models. We also assessed the expression level of ERCC6 and its association with patient survival in CRC.
Methods
Cell culture and treatment
CRC cells (HCT116, DLD1) were cultured in DMEM medium supplemented with 10% serum at 37°C with 5% CO2.
Study subjects
The study was approved by the Ethical Committee of Tianjin Medical University General Hospital, and informed consents were obtained from all patients. Thirty-eight pairs of CRC tissue samples and matched nontumor adjacent tissues were obtained from CRC patients who went through surgical resection at Tianjin Medical University General Hospital, between February 2013 and March 2015. All the tissues were snap-frozen in liquid nitrogen and stored at −80°C until use. In addition, an independent cohort of 74 patients who received 5-fluorouracil (5-FU)-based chemotherapy was enrolled. Clinical response to chemotherapy was evaluated as follows: complete resolution of the tumor (CR); partial response (PR), that is 50% decrease in tumor volume; stable disease (SD), that is 50% decrease or a 25% increase in tumor volume; and progressive disease (PD), that is 25% increase in tumor volume. Pathological response was determined by the final postoperative pathological analysis. Patients were classified as responders (CR or PR) and nonresponders (SD or PD). The prognosis was assessed in all CRC patients in December 2015.
Chemosensitivity assay
Cells were seeded in 96-well microtiter plates at a density of 3 × 103 cells/well 24 h later, chemo drugs were then added and incubated for 72 h. Cell viability was evaluated with CellTiter-Glo® assay (Promega, Madison). Each value was normalized to control wells with DMSO. IC50 value was determined by Graphpad Prism software.
Viral transductions and stable selections
About 1 μg of ERCC6 harpin RNA ([shRNA] Origene), 0.4 μg pMD2G, and 0.6 μg psPAX2 were transfected into 293FT cells using Effectene reagent (Qiagen, Valencia, CA). Viral supernatants were collected 48 h after transfection and added into cells after being cleared through a 0.45-μm filter with addition of 4 μg/mL polybrene (Sigma, St. Louis, MO).
Xenograft experiments
All animal experiments were approved by the Institutional Animal Care and Use Committee of the National Cancer Center. HCT116 cells with nonsilencing shRNA or ERCC6 shRNA (3 × 106 cells/injection) were subcutaneously injected into both flanks of 4–6 weeks old female nude mice (n = 5/group). 5-FU was administered when the tumor reached ∼20–30 mm3 every other day at 1 mg/kg. Tumor volumes were measured with caliper and calculated by a formula (volume = [length × width2]/2). The results represent as mean tumor volumes with SD.
Immunohistochemistry staining
The paraffin-embedded sections were immersed in 0.1 M citric acid buffer (pH = 6.0) and heated using microwave at 100°C for 10 min for antigen retrieval, followed by incubation with primary antibodies at 4°C overnight. After 1 h incubation with secondary antibody incubation at room temperature, the slides were developed using 0.05% diaminobenzidine with 0.01% hydrogen peroxidase. For negative controls, normal goat serum was incubated at 4°C overnight instead of antibody of interest.
Total RNA extraction
Tissue sections were minced into small fragments with scissors and homogenized using TRIzol™ reagent (Takara Bio, Inc., Otsu, Japan), followed by addition of 200 μL of chloroform. The mixture was then centrifuged at 12,000 × g for 15 min at 4°C, and the upper layer was transferred to a clean Eppendorf tube, supplemented with an equal volume of isopropanol. The mixture was centrifuged at 12,000 × g for an additional 15 min at 4°C. After washing the precipitation with 70% ethanol, total RNA was dissolved in RNase-free water and RNA concentrations were measured by optical density.
Quantitative reverse transcription polymerase chain reaction analysis
cDNA was reverse transcribed using Bio-Rad S1000 Thermal Cycler (Bio-Rad Laboratories, Hercules, CA). Briefly, 1 μg of the total RNA for each sample was reverse transcribed in a 20 μL reaction volume, containing 0.5 μg of oligo (dT). All samples were amplified as follows: 95°C for 2 min, 36 cycles of 95°C for 15 s, 60°C for 30 s, and 72°C for 20 s. RT-qPCR reaction was performed on the Bio-Rad Fluorescence Thermal Cycler (Bio-Rad Laboratories, Hercules) in triplicates using the following conditions: 95°C for 10 min; amplification for 35 cycles, denaturation at 95°C for 30 s; annealing at 56°C for 30 s; and elongation at 72°C for 30 s, followed by a final extension at 72°C for 10 min. GAPDH mRNA level was used for normalization.
Statistical analysis
Data represent mean ± SD. Statistical significance was evaluated by Student t-test. For patient sample data, statistical analysis was conducted using IBM SPSS Statistics Version 16 (SPSS, Inc, Chicago, IL). The Wilcoxon test was used to compare ERCC6 expression in paired tumor samples and NATs. Survival curves were constructed with the Kaplan–Meier analysis and assessed using log-rank test. Differences were considered statistically significant when p value is less than 0.05.
Results
Knockdown of ERCC6 confers sensitivity to 5-FU in CRC cells
To investigate the effect of ERCC6 on chemosensitivity, we used short shRNA to stably knockdown ERCC6 in HCT116 and DLD1 human CRC lines. Two independent shRNAs efficiently decreased ERCC6 mRNA levels (Fig. 1A, B). As shown in Figure 1C, D, downregulation of ERCC6 sensitized both HCT116 and DLD1 cells to 5-FU significantly.
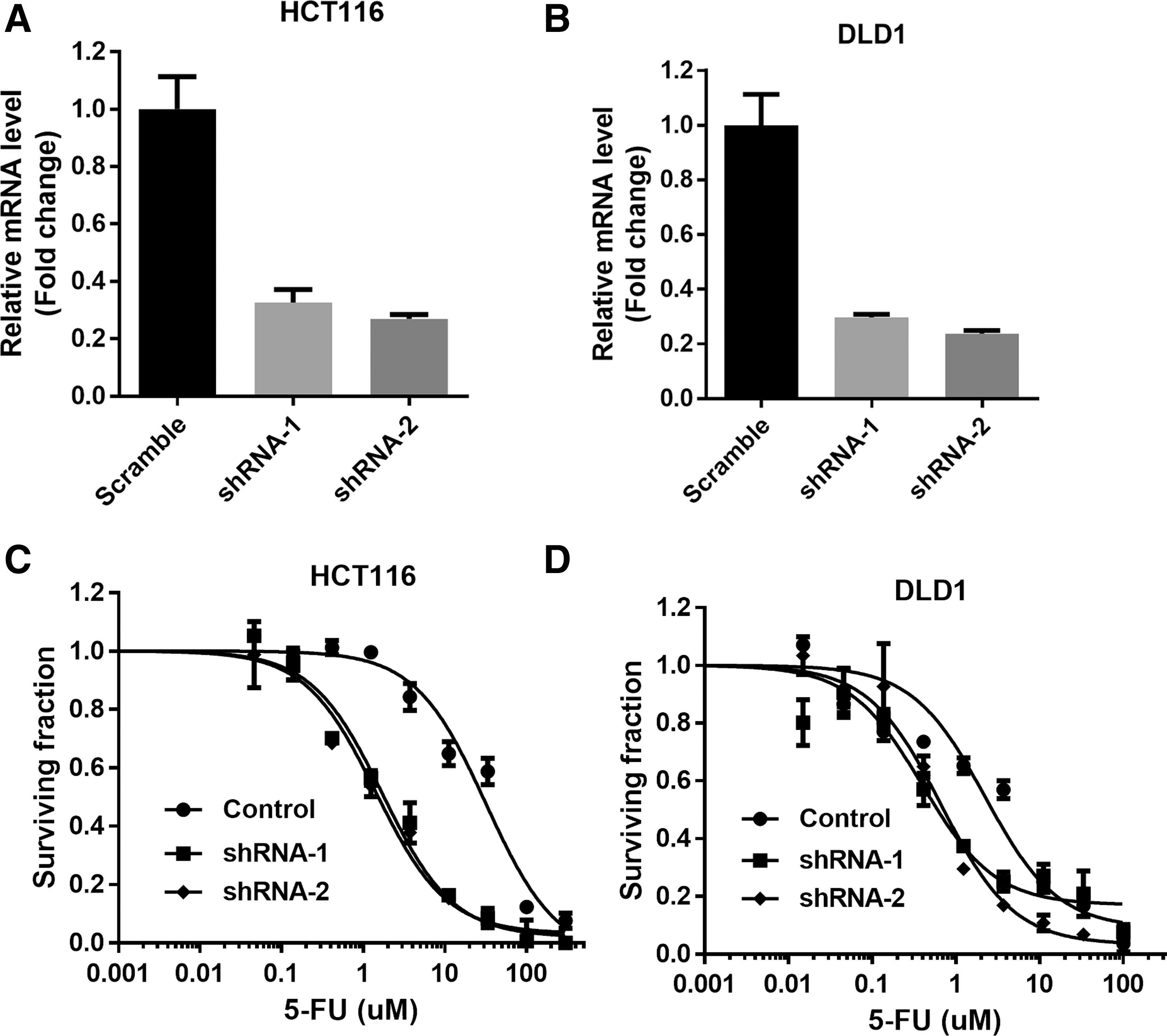
Knockdown of ERCC6 confers sensitivity to 5-FU in CRC cells.
Knockdown of ERCC6 enhances antitumor activity of 5-FU in xenograft mouse model
To evaluate whether ERCC6 modulates chemosensitivity in vivo, we established xenograft mouse models with HCT116 cells expressing Nsi shRNA or ERCC6 shRNA-2, which showed better knockdown efficiency. Mice bearing control or ERCC6 knockdown xenografts were injected with vehicle or 5-FU. Knockdown of ERCC6 slightly decreased tumor growth rate compared to control xenografts. Administration of 5-FU significantly reduced tumor growth rate and tumor weights in HCT116 control xenografts and this reduction was further enhanced in ERCC6 knockdown xenografts (Fig. 2A, B), indicating that downregulation of ERCC6 can sensitize HCT116 cells to chemo drugs. Consistently, 5-FU treatment led to induction of cleaved caspase 3 in control tumor sections and this induction was more dramatic in ERCC6 knockdown tumor sections (Fig. 2C).

Knockdown of ERCC6 enhances antitumor activity of 5-FU in xenograft mouse model.
ERCC6 is upregulated in CRC patient and patients resistant to 5-FU treatment
Quantitative PCR was performed to evaluate ERCC6 mRNA level in 38 pairs of CRC tumor tissues and their matched NATs. As shown in Figure 3A, ERCC6 was significantly upregulated in CRC tumor tissues. In addition, ERCC6 mRNA level was increased in tumor tissues from CRC patients who were resistant to 5-FU therapy compared to those who responded (Fig. 3B).
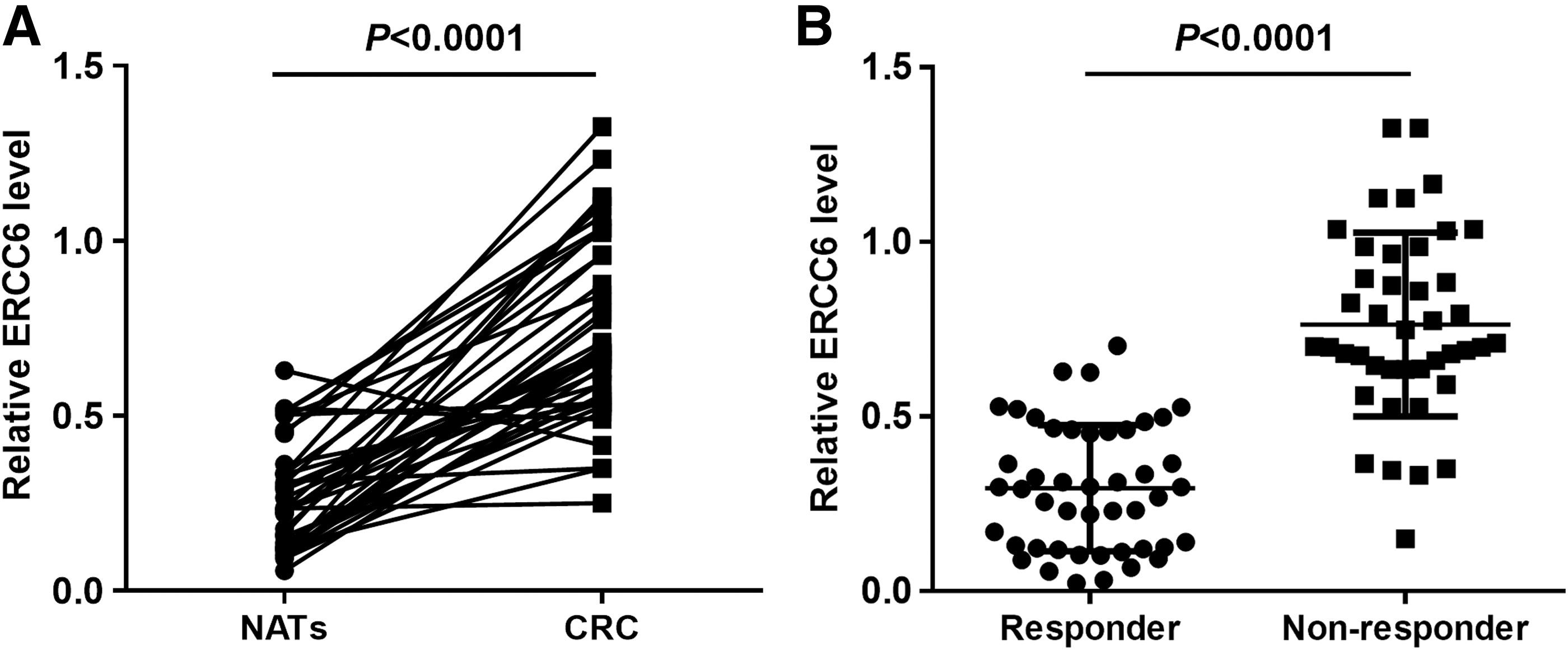
ERCC6 is upregulated in CRC patient and patient who are resistant to 5-FU treatment.
Elevated expression of ERCC6 is correlated with worse patient survival in CRC
Next, we examined whether ERCC6 expression level was correlated with CRC patient survival. Kaplan–Meier survival curves were constructed for two cohorts of patients who did or did not receive 5-FU chemotherapy. Figure 4A, B showed that patients expressing higher level of ERCC6 had worse survival in both cohorts, suggesting that ERCC6 can serve as a biomarker for CRC patient survival.

Elevated expression of ERCC6 is associated with worse overall survival in CRC patients with or without 5-FU chemotherapy.
Discussion
It is known that many commonly used chemotherapy drugs, such as 5-FU, exert cytotoxic effects in highly proliferating cancer cells by inducing intrastrand and interstrand cross-links with resultant DNA damage (Koberle et al., 2010; Galluzzi et al., 2012). These lesions trigger the activation of a series of DNA damage repair (DDR) responses with concomitant increases in the transcription of genes that are directly involved in NER to remove those lesions (Rieger and Chu 2004; Boerma et al., 2005). Therefore, expression levels and activity of the components of DDR pathways play critical roles in regulating the cellular response to chemo drugs.
ERCC enzymes are the key components of NER pathway. Accumulating evidences suggest that several ERCC family members are implicated in chemoresistance in various cancer types with ERCC1 as the most well studied. For example, low ERCC1 expression is reported to be associated with better survival in bladder cancer patients receiving neoadjuvant chemotherapy (Ozcan et al., 2013). Overexpression of ERCC1 is found to contribute to the resistance of lung cancer cells toward cetuximab combined with cisplatin (Li et al., 2009). In addition to ERCC1, dysregulation of ERCC2 and ERCC5 has also been reported to affect responses to 5-FU or oxaliplatin-based chemotherapy in ovarian cancer and CRC, respectively (Gornati et al., 1997; Chen et al., 2009). However, few, if any, studies have been reported regarding the role of ERCC6 in regulating chemoresistance. In the current study, we observed that downregulation of ERCC6 conferred sensitivity to 5-FU in CRC cells both in vitro and in vivo (Figs. 1 and 2), suggesting the direct involvement of ERCC6 in regulating response to 5-FU chemotherapy.
Polymorphisms of multiple genes in NER pathway, including ERCC6, have been detected in bladder, prostate, and more recently gastric cancer patients (Ramaniuk et al., 2014; Zhang et al., 2014; Liu et al., 2016). In addition, mutations in ERCC6 gene has been detected in premalignant human colonic epithelial cells, suggesting its involvement in early stage of CRC development (Zhang et al., 2013, 2015). Dysregulation of ERCC6 has been correlated with patient survival and proposed as a potential biomarker in several malignancies (Garcia-Closas et al., 2006; Matakidou et al., 2007; Abbasi et al., 2009). In our analysis, we observed upregulation of ERCC6 in CRC tissues compared with NATs and in particular, it was upregulated in those patients who were resistant to 5-FU chemotherapy (Fig. 3). Importantly, low expression of ERCC6 was associated with better survival in CRC patients with or without receiving 5-FU treatment (Fig. 4), indicating that ERCC6 expression may be used as a potential predictor for chemo response and biomarker for patient survival in CRC.
Conclusions
In conclusion, we have identified the role of ERCC6 in regulating chemo response in CRC cells. ERCC6 is upregulated in CRC patients and particularly in those who are chemoresistant. Elevated expression of ERCC6 is associated with poor patient survival. Therefore, ERCC6 represents a novel therapeutic target to improve chemotherapeutic efficacy and a potential biomarker for CRC patient survival.
Footnotes
Acknowledgment
This study was supported by the Science and Technology Fund of Tianjin Health Bureau (2014KZ123).
Disclosure Statement
No competing financial interests exist.