
Abstract
Schizophrenic patients tend to have reduced incidence of some cancers due to the treatment of antipsychotic drugs with antitumor effects, such as chlorpromazine and trifluoperazine (TFP). Forkhead Box O1 (FOXO1) as tumor suppressor in many malignancies is often inactivated by nuclear export, which could be inhibited by TFP. However, the antitumor efficiency of TFP and related role of FOXO1 in hepatocellular carcinoma (HCC) are unclear. Thus, two HCC cell lines SMMC-7721 and Bel-7402 were treated with different concentrations of TFP and the IC50 was determined. We found that TFP could inhibit the vitality of two cell lines and induce cell cycle arrest at G0/G1. Meanwhile, the apoptosis was also increased and the ability of migration or invasion was found to be impaired by TFP. Interestingly, TFP reversed the cytoplasmic localization of FOXO1 to nuclear and increased its expression in nuclear, and increased the ratio of Bax/Bcl-2. However, knockdown of FOXO1 significantly abrogated the TFP-induced apoptosis by decreasing the Bcl-2 expression. Furthermore, we found that TFP in vivo could effectively restrict the angiogenesis and tumor growth with reduced expression of VEGF, Bcl-2, and PCNA, and increased the nuclear localization of FOXO1, which indicated its antitumor role in HCC.
Introduction
A
Although the patients with schizophrenia have high likelihood of smoking, alcohol abuse, obesity, and poor diet, the incidence of cancer tends to be not significantly higher than those in the general population (Catts et al., 2008). Importantly, many studies reported that patients with schizophrenia showed a reduced incidence of some types of cancer, especially in prostate cancer, colon cancer, and malignant melanoma (Catts et al., 2008; Preti and Wilson, 2011). The potential factors involved in the reduced cancer risk remain elusive. Genetic factors and antipsychotic drugs had been found to participate to the interaction between cancer and schizophrenia (Mortensen, 1989; Dalton et al., 2006). Dalton et al. (2006) had found that neuroleptic medications, such as chlorpromazine and other phenothiazine compounds, trifluoperazine (TFP), were associated with a reduced risk of rectal cancer in both men and women. Currently, many antipsychotic agents showed antitumor effects in vitro and vivo. The combination of chlorpromazine and an antiparasitic agent (pentamidine) could effectively inhibit the growth of tumor cell lines through the inhibition of mitotic kinesin KSP/Eg5 to decrease tumor cell proliferation (Lee et al., 2007). TFP was found to upregulate the expression of early growth response gene-1 (Egr-1), which could repress the tumor cell growth by the inhibition of cyclin D1 (Shin et al., 2004). Besides, Polischouk et al. (2007) reported that TFP might be capable of inhibiting one or more elements of the DNA repair machinery, which consolidated the bleomycin cytotoxicity in lung cancer cells. However, the overall mechanisms of TFP-mediated tumor killing remain to be further investigated.
Forkhead Box O1 (FOXO1) is a transcriptional regulator of the G1/S checkpoint and apoptosis, and has been identified as a tumor suppressor in cervical cancer (Hou et al., 2014), prostate cancer (Zhang et al., 2011), and gastric cancer (Park et al., 2017). Several studies identified that FOXO1 was functionally inactivated by AKT-mediated phosphorylation, leading to results in CRM-1-dependent nuclear export, proteasomal degradation, and reduced transcriptional activity during carcinogenesis (Aoki et al., 2004). Interestingly, TFP had been demonstrated to inhibit FOXO1 nuclear export, restored sensitivity to AKT-driven chemotherapy resistance through the reactivation of KLF6/FOXO1 signals in both cell culture and xenograft models of lung adenocarcinoma (Sangodkar et al., 2012). However, whether TFP could inhibit the progression of HCC by FOXO1 remains unknown.
Previous bioinformatics analysis on two cohorts of patients with primary HCC or Hepatitis B virus-positive HCC patients (
Materials and Methods
Cell culture and reagents
The human HCC cell line SMMC-7721 and Bel-7402 cells were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS; Life Technologies) and ampicillin and streptomycin at 37°C and 5% CO2 conditions. VP16 was purchased from Sigma. TFP hydrochloride was acquired from Sigma-Aldrich (St. Louis, MO). Anti-FOXO1, Bax and Bcl-2, and VEGF antibodies were obtained from Abcam (USA) and Cell Signaling Tech (Denver, MA). The siRNA targeting FOXO1 was purchased from RiboBio (Guangzhou, china), siRNA-FOXO1: 5′-GGAGGUAUGAGUCAGUAUATT-3′.
CCK-8 assay
Cells were harvested after the treatment of TFP as indicated dose and washed with phosphate-buffered saline (PBS), and then cell counting kit-8 (Kumamoto, Japan) mixed with DMEM was used for cell viability assay, and the absorbance was measured at 450 nm by a microplate reader.
Hoechst staining assay
SMMC-7721 and Bel-7402 cells treated with TFP were cultured at 37°C for 48 h, stained with 0.1 μg/mL Hoechst 33342 (Sigma, St Louis, MO), and added to the culture medium. Fluorescence microscopy (OLYMPUS IX71; Olympus Corporation, Tokyo, Japan) with a filter for Hoechst 33342 (365 nm) was used to detect the changes of nuclear morphology.
Flow cytometer assay
For the apoptosis analysis, the cells were fixed in cold 70% ethanol at −20°C for 2 h. Ten milligram per milliliter RNase were added and stained with 2 μL of annexin V mixed with 2 μL of propidium iodide (PI, eBioscience), were used according to the manufacturer's instructions. For the cell cycle analysis, cells were stained with PI staining solution (10 μg/mL RNase A and 50 μg/mL PI) at 37°C for 30 min in dark. The cell cycle distribution was analyzed using a flow cytometry provided with the Cell-Quest software.
Scratch wound assay and transwell assay
For the scratch wound assay, the cells were wounded with a plastic tip that was dragged across the cell monolayer upon cells reaching confluence. Cells were incubated with a different dose of TFP for 24 h. Five fields were randomly selected and the distances of migrated cells were measured under a light microscope. In the transwell assay, 2 × 104 SMMC-7721 and Bel-7402 cells were cultured with TFP for 24 h in the upper chamber of a noncoated transwell insert. For the invasion assay, the upper chamber of the transwell inserts was coated with Matrigel, and tumor cells were plated in the upper chamber of the Matrigel-coated transwell insert. Cells that did not migrate or invade were removed using a cotton swab and were stained by crystal violet and counted under an inverted microscope. Five random views were selected to count the cells and the independent experiments were repeated thrice.
Western blots
According to the manufacturer's protocol, cells for Western blots were collected and total protein was isolated from the cell samples. Detailed procedures for immunoblotting are described elsewhere (Aoki et al., 2004). For nuclear cytoplasmic fractionation, cell lysates were extracted using NE-PER Nuclear and Cytoplasmic Extraction Reagents (Thermo Scientific) according to the manufacturer's protocol.
Cell transfection
8 × 104 SMMC-7721 and Bel-7402 cells were seeded into 12 plates and cultured in DMEM containing 10% FBS. For the knockdown assay, the cells were washed by PBS and transfected with siRNA-FOXO1 or negative control for 24 h at a concentration of 100 nM by Lipofectamine 2000 (Invitrogen) and treated with TFP for indicated times.
Immunofluorescence for FOXO1
SMMC-7721 cells were fixed with 4% formaldehyde in PBS for 15 min and rinsed with PBS thrice. Then cells were permeabilized with 100% methanol for 10 min at −20°C and blocked with 3% bovine serum albumin in PBS for 60 min, and incubated with primary antibodies overnight at 4°C. After rinsing thrice in PBS, incubated coverslips in fluorochrome-conjugated secondary antibody for 1–2 h at room temperature in dark and then stained the nucleus with diamidino-phenyl-indole. The coverslips were mounted onto the glass slides with neutral gum and observed by FV10i confocal microscope (OLYMPUS, Japan).
Immunohistochemistry for CD31
The microvessel density (MDV) in tumor tissues was analyzed as previously reported by IHC on 2-μm-thick, formalin-fixed, and paraffin-embedded specimen sections (Ding et al., 2017). Slides were incubated in xylene for 5 min and followed by two washes of 100% ethanol for 10 min and 95% ethanol for 10 min. Antigen unmasking was performed and then the slides were blocked with 3% hydrogen peroxide for 30 min at room temperature. Then, the primary antibody for CD31 was incubated with the formalin fixed paraffin-embedded specimen sections at 4°C overnight, and then the EnVision Detection System kit (DAKO, Denmark) was used for the diaminobezidin chromogen followed by nuclear staining using hematoxylin.
RNA isolation and quantitative real-time polymerase chain reaction
Total RNA from tumor tissues in each group was extracted using Trizol reagent (Invitrogen) according to the standard RNA isolation protocol. Quantitative real-time polymerase chain reaction (qRT-PCR) was performed and the expression levels of VEGF, Bcl-2, and PCNA were normalized to GAPDH for gene expression. The primers are listed in Table 1.
Tumor model
To investigate the antitumor role of TFP in vivo, 2 × 106 SMMC-7721 cells were subcutaneously injected in rear flank of nude mice (four per group). After volumes reached an average of 200 mm3, two doses of TFP (5 and 10 mg/kg, intraperitoneally) were administrated to each group five times, 3 days apart.
Statistical analyses
For statistical analyses, the Statistical Package for Social Sciences version 16.0 (SPSS 16.0; SPSS, Inc., Chicago, IL) and the Prism statistical software package (Version 5.0; Graphpad Software, Inc.) were used. Unpaired t-tests or Mann–Whitney U tests were used to compare the two groups, and multiple group comparisons were analyzed with one-way ANOVA. p < 0.05 was considered statistically significant. All experiments were performed at least thrice.
Results
TFP effectively inhibits cell vitality and induces G1 arrest in two HCC cell lines
To investigate the antitumor effects of TFP in HCC, a series of doses of TFP was prepared and two HCC cell lines SMMC-7721 and Bel-7402 were selected. The IC50 of TFP for two cell lines was determined and the results showed that the IC50 for SMMC-7721 was 25.25 μM and for Bel-7402 was 20.83 μM. Also, 5 μM TFP had showed inhibition effects on both cell lines (Fig. 1A, B). Therefore, 5 and 10 μM were chosen for further study.
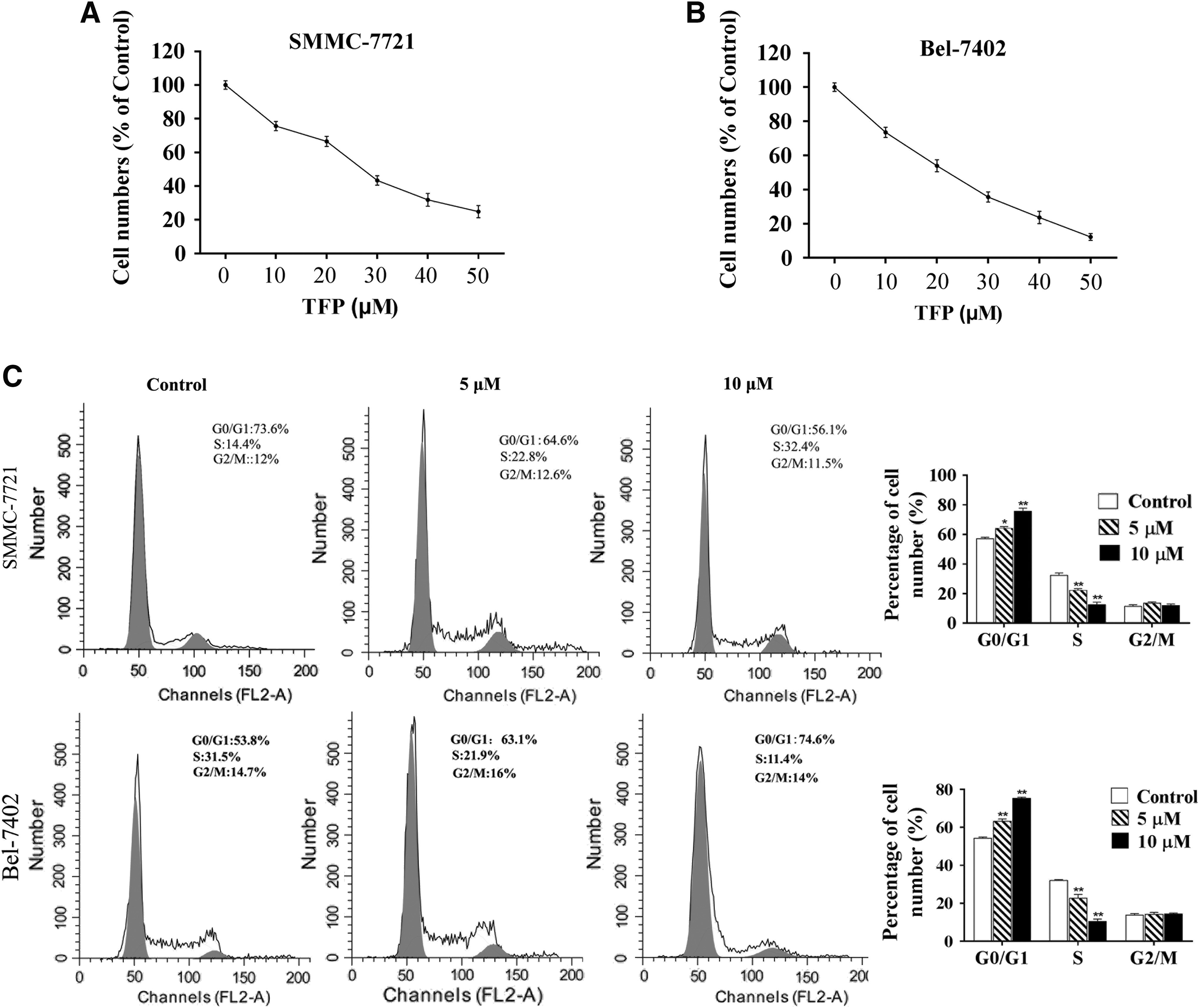
The IC50 of SMMC-7721
We proposed a hypothesis that the reduced cell vitality was attributed by cell cycle arrest. After the treatment of TFP, we found that two cell lines showed G0/G1 arrest and the S phase (%) was significantly decreased, which was dependent on the concentration of TFP (Fig. 1C). These data indicated that TFP could induce G1 arrest to inhibit cell vitality in two HCC cell lines SMMC-7721 and Bel-7402.
TFP promotes apoptosis in dose-dependent manner
The inhibition of cell cycle could activate the apoptosis pathway and initiate the apoptosis; thus we further analyzed the impacts of TFP on apoptosis. Two cells were treated with 5 and 10 μM TFP and the apoptosis was determined. The results showed that TFP improved the apoptosis in two doses and 10 μM TFP induced higher apoptosis than 5 μM TFP, which suggested that TFP could promote SMMC-7721 and Bel-7402 apoptosis in a dose-dependent manner (Fig. 2A). These findings were confirmed by Hochest stain assay, which showed similar results (Fig. 2B).
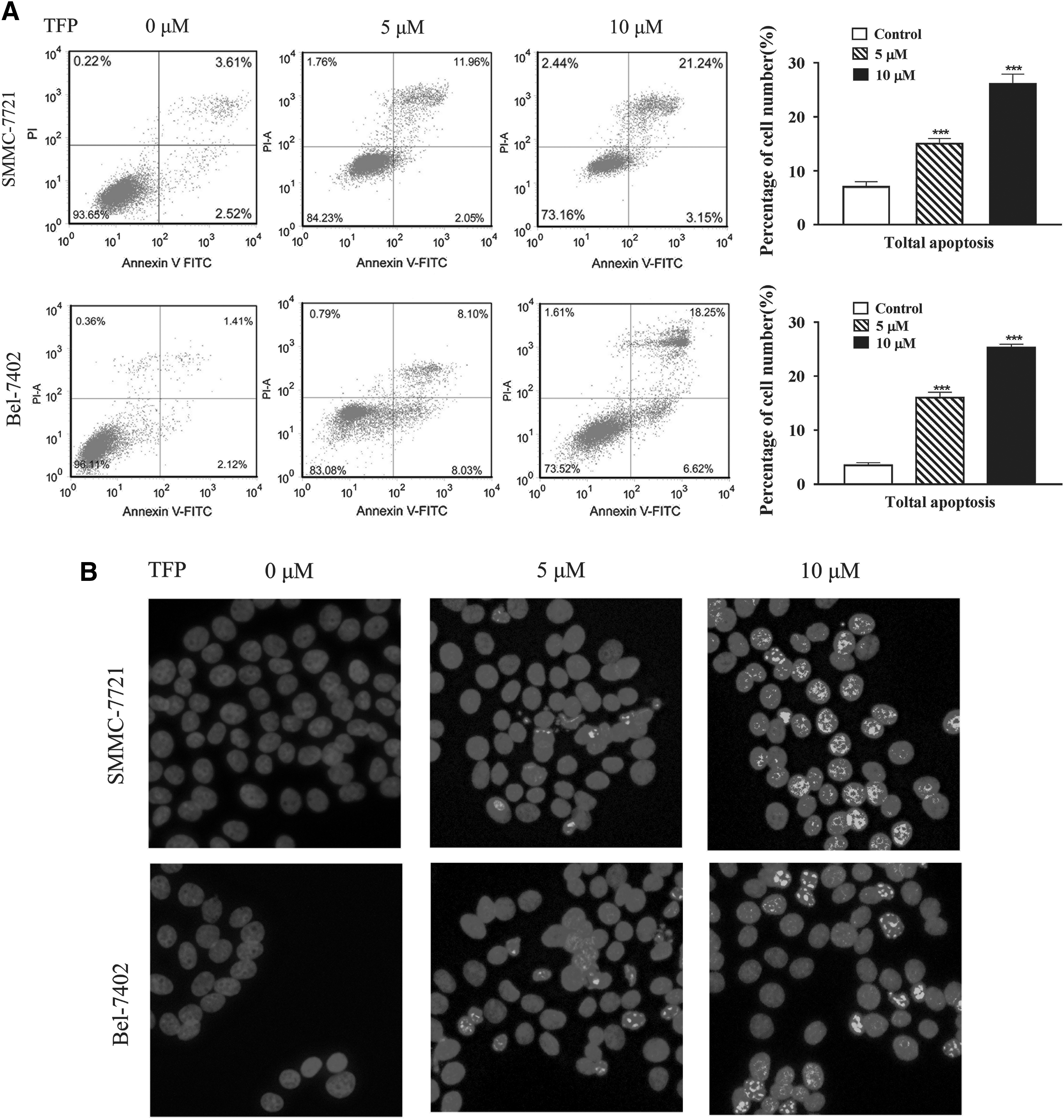
TFP could promote the apoptosis of SMMC-7721 and Bel-7402 cells. The apoptosis was determined by
TFP inhibits HCC cell migration and invasion
Several studies had reported that TFP could inhibit the ability of invasion in human prostate cancer cells. We, in this study, investigated the role of TFP in tumor migration and invasion. After the treatment of 5 and 10 μM TFP, the scratch experiment demonstrated that TFP could inhibit the ability of migration in SMMC-7721 and Bel-7402 (Fig. 3A). To confirm this result, transwell assay was performed and found that the migration and invasion of two cells were remarkably abrogated by TFP, and the inhibition efficiency was positively associated with TFP concentration (Fig. 3B, C).
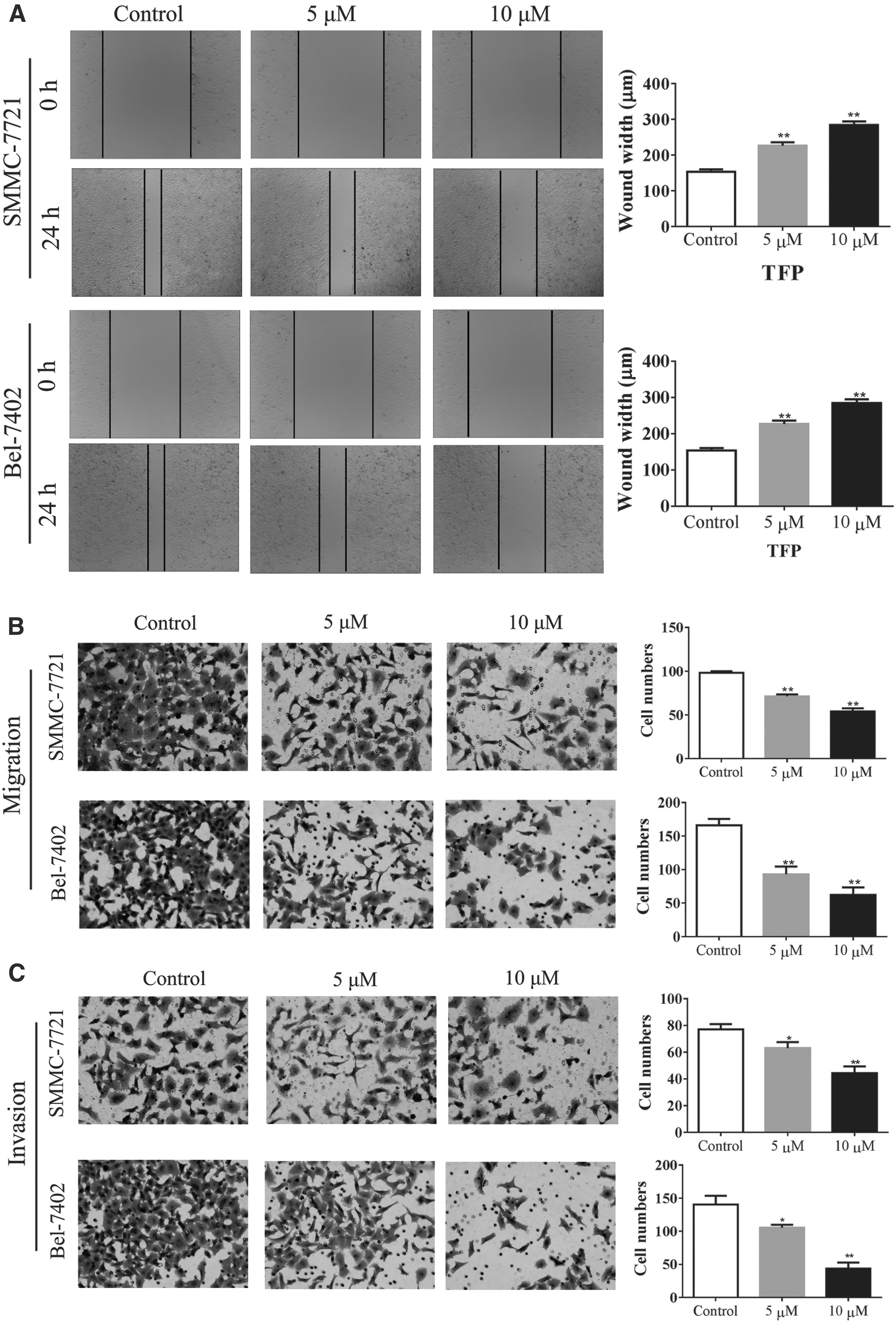
TFP could inhibit the migration and invasion of SMMC-7721 and Bel-7402 cells. The ability of migration was determined by
FOXO1 is reactivated by TFP and induces apoptosis by Bax/Bcl-2
The nuclear localization of FOXO1 promotes many transcription factors involving cell cycle DNA repair and apoptosis, such as p27, TRAIL, and Bcl-2. (Lam et al., 2006). As tumor suppressors, we investigated the role of FOXO1 in TFP-induced apoptosis. We found that the expression of FOXO1 in nuclear was significantly upregulated, whereas the cytoplasmic expression was downregulated by TFP (Fig. 4A). Also, TFP could promote the nuclear localization of FOXO1 (Fig. 4B). The expression of FOXO1 was knocked down by siRNA transfection in SMMC7721 and Bel-7402. Knockdown efficiency was determined by qRT-PCR and Western blot (Fig. 4C). Meanwhile, the level of antiapoptosis factor Bcl-2 was decreased, but the proapoptosis factor Bax was enhanced by TFP (Fig. 4D). Of note, the ratio of Bax/Bcl-2, which indicated the apoptosis, was increased by TFP, but knockdown of FOXO1 could restore the expression of Bcl-2 and inhibit the expression of Bas to downregulate the ratio of Bax/Bcl-2, leading to impaired TFP-induced apoptosis. These results implicated that FOXO1 is required for TFP-induced apoptosis, which might be involved with the balance of Bax/Bcl-2.

TFP increases the expression of FOXO1 in nuclear for Bcl-2-dependent apoptosis.
TFP activates FOXO1 and restricts tumor angiogenesis and growth in vivo
Given the antitumor effects of TFP in vitro, we next estimated the efficiency of TFP for tumor inhibition in vivo. The xenograft model of human SMMC-7721 was established and the nude mice were treated with different doses of TFP. The results showed that the administration of TFP could effectively inhibit tumor growth (Fig. 5A), but had no significant cytotoxicity to mice (Fig. 5B), which might be related to decreased MDV (Fig. 5C). The HE staining suggested that TFP treatment exhibited some areas of necrotic cells (Fig. 5D). The expression of VEGF was found to be reduced in mice treated with TFP and the tumor proliferation indicated that PCNA and antiapoptosis factor Bcl-2 were also inhibited by TFP (Fig. 5E). In addition, the cytoplasmic and nuclear expression of FOXO1 were analyzed and we found that TFP treatment in vivo also could activate FOXO1 and promote its nuclear localization (Fig. 5F). Collectively, TFP is a potential antitumor drug in HCC and this effect involves the activation of FOXO1.
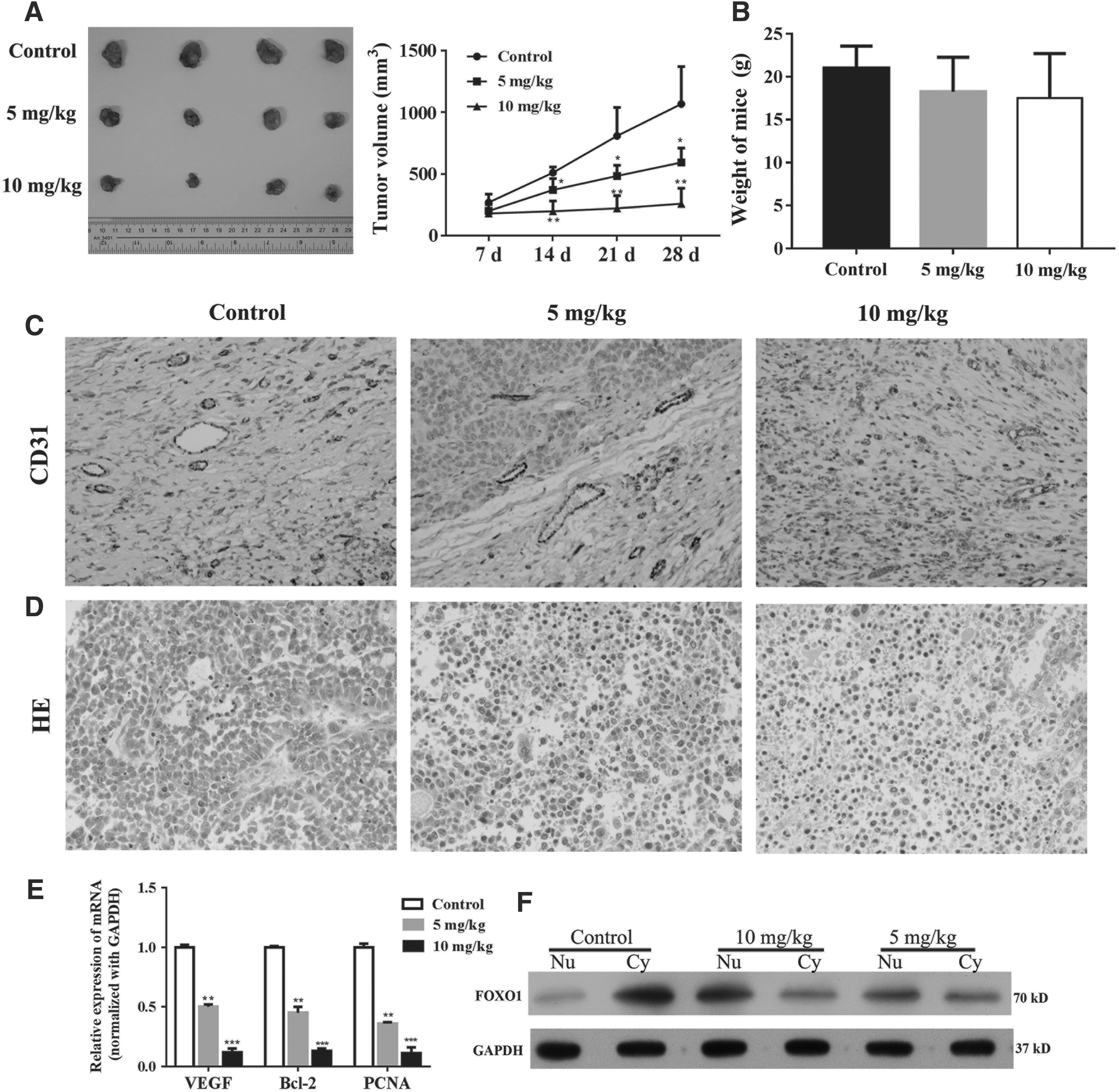
The antitumor role of TFP was assessed in vivo. 2 × 106 SMMC-7721 cells were subcutaneously injected in rear flank of nude mice (four per group) and 5 and 10 mg/kg TFP were administrated to mice.
Discussion
Phenothiazine drugs are widely used antipsychotic drugs that mainly act on central dopamine receptors and have sedative, antiemetic, antipsychotic, and body temperature-lowering effects (Shen, 1999). The representative drugs are chlorpromazine and its derivatives, fluphenazine and TFP. In this study, we found that TFP could promote apoptosis, cell cycle arrest of HCC cells, which was mediated by FOXO1 activation in vitro and vivo.
Actually, antipsychotic drugs have shown antitumor effects through different ways, including directly killing tumor cells, reverse tumor chemotherapy resistance, or inhibiting tumor metastasis. Currently, on the one hand, TFP had direct cytotoxicity for tumor cells. In the malignant peripheral nerve sheath tumors (MPNSTs), TFP monotherapy could inhibit the tumor cell proliferation and abrogate the growth of xenografted MPNST cells (Brosius et al., 2014). In human leukemic cell lines, TFP induced apoptosis of leukemic cells through a “nonclassical” pathway that suppression of mitochondrial DNA polymerase and decrease of ATP production, but TFP had any influence on the viability of normal lymphocytes (Zhelev et al., 2004). Besides, TFP was found to upregulate the expression of tumor suppressor Egr-1 and downregulate cyclin D1 to induce cell cycle arrest in U87MG glioma cells (Shin et al., 2004). On the other hand, TFP had indirect cytotoxicity for tumor cells. Yeh et al. found that TFP inhibited spheroid formation of cancer stem-like cell (CSC) and downregulated CSC markers (CD44/CD133). The combination of TFP with gefitinib or cisplatin could effectively inhibit the lung cancer growth (Yeh et al., 2012). TFP was also found to be capable of inhibiting key factors of the DNA double-strand break repair pathway and promoted the cytotoxicity of bleomycin in lung cancer cells (Polischouk et al., 2007). The drug tamoxifen could inhibit MPNST proliferation and survival, but upregulate the expression of serum/glucocorticoid regulated kinase (SGK) for tamoxifen resistance (Brosius et al., 2014). TFP could improve chemotherapy of tamoxifen by inhibiting the expression of SGK. We, in this study, found that in HCC, TFP could directly decrease tumor cell vitality and induce G0/G1 arrest and apoptosis by increasing the ratio of Bax/Bcl-2 in vitro. Meanwhile, the treatment of TFP also inhibited MDV and VEGF level in vivo, which indirectly restricted tumor growth. Interestingly, Cao et al. also found that TFP significantly reduced angiogenesis with decreased expression of VEGF and prevented cancer cell invasion through decreased phosphorylated AKT and β-catenin (Pulkoski-Gross et al., 2015).
As tumor suppressors, FOXO factors participate into regulation of cell cycle arrest, DNA repair, and apoptosis through cyclin D1/2, Bcl-6, FasL, and TRAIL (Lam et al., 2006). FOXOs activate transcription of the cell cycle inhibitor p27 and p21, while they repress transcription of cyclin D1 and cyclin D2 (Greer and Brunet, 2008). In nonsmall cell lung carcinoma, downregulation of FOXO1 by siRNAs significantly promoted cell proliferation, decreased S phase cell numbers, and increased G2/M cell population (Hou et al., 2016). However, PI3K-/PKB-dependent phosphorylation contributes to the loss of function of FOXO transcription factors and subsequent proteasomal degradation, which has emerged as an important event during carcinogenesis (Aoki et al., 2004). Sangodkar et al. (2012) reported that TFP was able to inhibit FOXO1 nuclear export and restored sensitivity to AKT-driven erlotinib resistance through activation of KLF6/FOXO1 signaling cascade in lung adenocarcinoma, which was consistent with previous findings that TFP could inhibit calmodulin activity to relocalize FOXO1 to nucleus through a calmodulin-dependent regulatory mechanism (Kau et al., 2003). In this study, we found that TFP increased the nuclear expression of FOXO1 through inhibition of its cytoplasmic localization and induced the expression of Bax, but impaired the Bcl-2 level. Therefore, the ratio of Bax/Bcl-2 was elevated by TFP for apoptosis.
In summary, in HCC, TFP has direct antitumor effects in vitro and vivo by impairing cell proliferation, inducing cell cycle arrest and apoptosis through FOXO1-regulated balance of Bax/Bcl-2, and indirect antitumor effects through inhibition of angiogenesis.
Footnotes
Acknowledgments
We here thank professor Huang from Southern Medical University for the microarray data of HBV+HCC patients. This work was supported by a Grant from Social Development Special Project of Science and Technology Department of Hainan Province Key Research (Grant number ZDYF2016123).
Disclosure Statement
No competing financial interests exist.