
Abstract
The zinc metalloprotease ZMPSTE24 is a constitutively and ubiquitously expressed host restriction factor that is responsible for limiting infection by a broad spectrum of enveloped viruses, including influenza A, vesicular stomatitis, zika, ebola, Sindbis, cowpox, and vaccinia viruses, but not murine leukemia or adenovirus. Antiviral function is independent of ZMPSTE24 enzymatic activity. Protein interaction and genetic complementation studies indicate that ZMPSTE24 is a component of a common antiviral pathway that is associated with interferon-induced transmembrane proteins. In vivo studies with zmpste24-deficient mice demonstrate the importance of ZMPSTE24 for antiviral defense.
Introduction
E
During viral and vertebrate co-evolution, hosts developed a battery of antiviral effectors to impede infection. Many of these effectors are amplified by interferon. The interferon-induced transmembrane (IFITM) proteins (IFITM1, IFITM2, and IFITM3) are a family of antiviral effectors that restrict endosomal entry of multiple pH-dependent enveloped viruses (Brass et al., 2009; Perreira et al., 2013; Bailey et al., 2014). The antiviral function of murine IFITM genes was subsequently confirmed by in vivo studies with influenza A, West Nile, Venezuelan equine encephalitis, and chikungunya virus (Bailey et al., 2012; Everitt et al., 2012; Gorman et al., 2016; Poddar et al., 2016). Furthermore, patients hospitalized for severe influenza A infection show enrichment for a minor ifitm3 allele (Everitt et al., 2012; Yang et al., 2015). Thus, the significance of IFITM proteins for innate defense has been well established both in vitro and in vivo. However, the mechanisms governing antiviral activity remain poorly understood.
The IFITM3 Proteome
Affinity purification coupled with mass spectrometry (AP-MS) was performed to discover potential co-factors that contribute to IFITM3 antiviral activity. HEK293 cells stably expressing C-terminal FLAG-tagged IFITM3 were treated with interferon or control medium. Affinity purification was used to enrich IFITM3 protein complexes (Fu et al., 2017). The SAINT algorithm (Choi et al., 2011) was adopted to analyze AP-MS data in combination with a reference database of 194 protein complexes purified under identical conditions (Fu et al., 2015). SAINT identified 21 high-confidence interacting proteins, most of which are associated with endoplasmic reticulum or endosome (Fig. 1). Interactions between IFITM3 and ZMPSTE24 are enhanced by interferon stimulation, whereas interactions with TTC7B (a regulator of phosphatidylinositol 4-phosphate synthesis) are diminished. The results reflect the dynamic interferon-driven modulation of the IFITM3 proteome. The enhanced association with ZMPSTE24 may reflect interferon-induced posttranslational modifications in either binding partner.
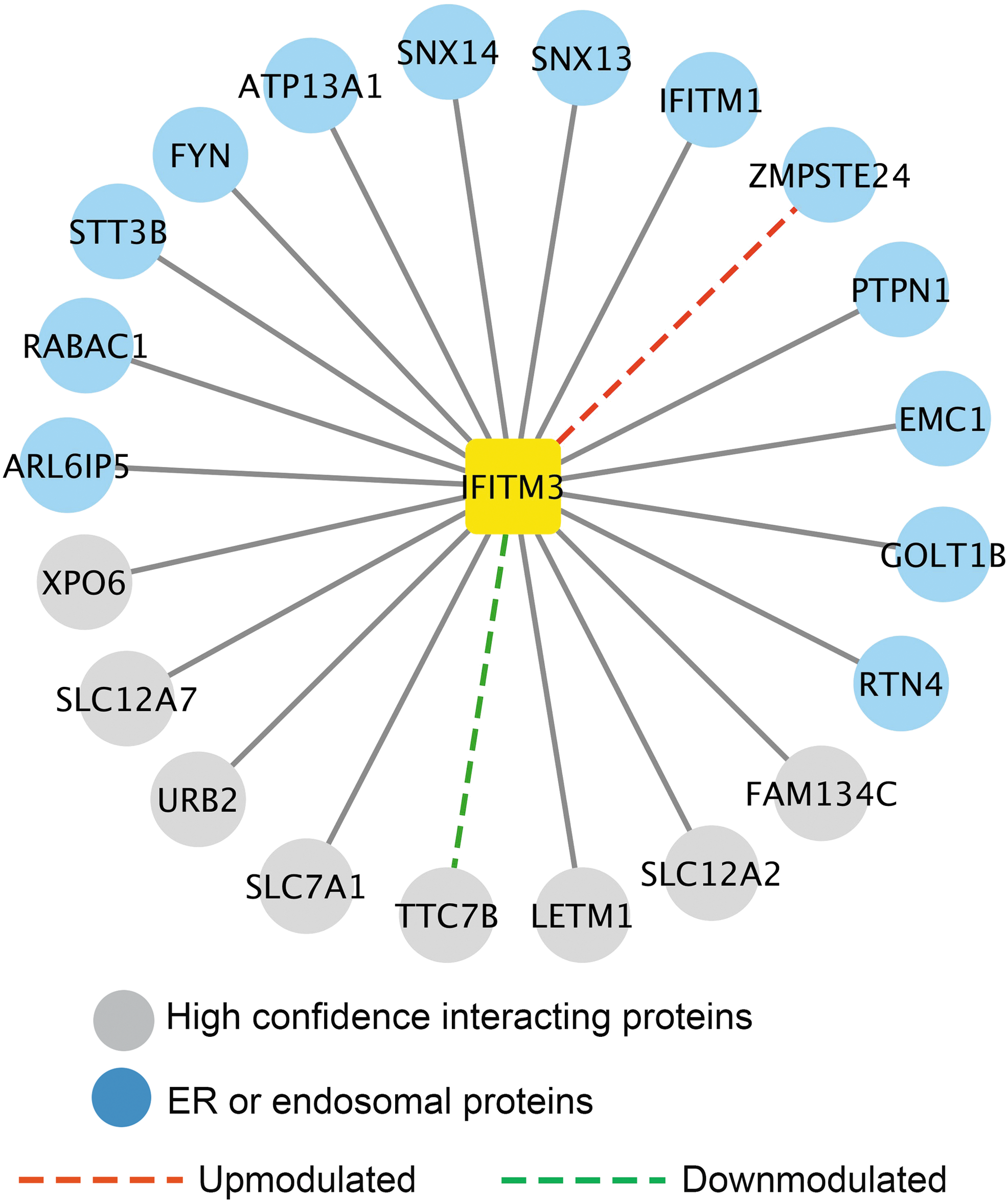
Overview of the IFITM3 interactome. SAINT algorithm was used to analyze mass spectrometry data (Fu et al., 2017). High-confidence interacting proteins are shown in circles. ER and endosomal proteins are highlighted in blue. Red dashed line indicates interactions that are upmodulated (>5-fold increase) by type 1 interferon, and dashed green line represents >5-fold downregulation after interferon treatment. Complete dataset can be downloaded from
ZMPSTE24 contains seven transmembrane domains and a canonical HEXXH metalloprotease motif. As opposed to IFITM1-3 proteins, ZMPSTE24 expression is not upregulated by type 1 interferon or viral infection (Fu et al., 2017). ZMPSTE24 is evolutionarily conserved; orthologues are present in metazoans and yeast. Human ZMPSTE24 and its yeast orthologue have a nearly identical crystal structure (Pryor et al., 2013; Quigley et al., 2013). ZMPSTE24 has promiscuous substrate specificity, cleaving prenylated and non-prenylated substrates. ZMPSTE24 is expressed on intracellular membranes and assists with the clearance of clogged translocons in the endoplasmic reticulum (Ast et al., 2016; Hildebrandt et al., 2016). In mammals, ZMPSTE24 is required for maturation of the lamin A nuclear scaffold protein. Mutations in the ZMPSTE24 gene that result in decreased enzyme function cause a spectrum of diseases that share certain features of Hutchinson-Gilford Progeria Syndrome (HGPS), including premature aging. Reduced ZMPSTE24 enzymatic activity correlates with disease severity (Barrowman et al., 2012). Zmpste24-deficient mice display defects in prelamin A processing and develop a progeriod phenotype that phenocopies the main clinical manifestations of HGPS (Bergo et al., 2002; Pendas et al., 2002).
Specificity of ZMPSTE24 Antiviral Activity
Overexpression of ZMPSTE24 inhibited production of infectious influenza A virus (IAV) particles, repressed transcription of IAV proteins, and reduced IAV reporter activity (Fu et al., 2017; Wang et al., 2017). Inhibition of viral transcription is observed within 2 h, suggesting an intrinsic defense mechanism, as there is little time for protein synthesis. Ectopic ZMPSTE24 expression also impeded infection by other fusogenic enveloped RNA viruses, including vesicular stomatitis virus (VSV), ebola, zika, and Sindbis virus. However, infection with murine leukemia virus (MLV) was not impacted. Although MLV also utilizes a fusogenic entry process, it is pH independent and can directly penetrate the plasma membrane (Rasmussen and Vilhardt, 2015).
The ability of ZMPSTE24 to restrict infection of human cells by enveloped DNA viruses was also examined. Neither IFITM3 nor ZMPSTE24 restricts adenovirus (AdV) (Warren et al., 2014; Fu et al., 2017). Although AdV also enters cells via a pH-dependent endocytic route, it uses a lysogenic rather than a fusogenic strategy to escape the endosome (White and Whittaker, 2016). Orthopoxviruses (e.g., Vaccinia virus [VACV] and cowpox) exploit pH-dependent fusogenic strategies for endocytic entry (Schmidt et al., 2012). Overexpression of ZMPSTE24 or IFITM3 inhibits infection with VACV or cowpox virus (Fu et al., 2017). Thus, IFITM and ZMPSTE24 can suppress the infection of human cells by DNA and RNA viruses.
To substantiate the role of ZMPSTE24 in antiviral defense, zmpste24−/− mouse embryonic fibroblasts (MEF) were examined for susceptibility to IAV infection. ZMPSTE24 deficiency resulted in increased viral protein expression, enhanced viral reporter activity, and higher titers of infectious viral particles in zmpste24−/− compared with zmpste24+/+ MEF (Fu et al., 2017). Zmpste24−/− cells were also more susceptible to infection with VSV, Sindbis, MLV, VACV, and cowpox. However, genetic deficiency of ZMPSTE24 did not impair host restriction to MLV or AdV (Fu et al., 2017). Reconstitution of zmpste24−/− MEF with human ZMSPTE24 restored antiviral activity. The effects of ZMPSTE24 deficiency were validated with human cells. The silencing of ZMPSTE24 increased IAV, VSV, Sindbis, cowpox, and VACV reporter activity in primary human respiratory epithelial cells (Fu et al., 2017). The combined gain- and loss-of-function experiments demonstrate a role for ZMPSTE24 in host defense to selected RNA and DNA viruses that rely on pH-dependent fusogenic strategies for endosomal entry. The specificity of ZMPSTE24 restriction generally mirrors that of IFITM. However, quantitative differences in potency occur; VSV is more susceptible than IAV to blockade by ZMPSTE24, whereas the reverse is reported for IFITM proteins (Diamond and Farzan, 2013). More viruses need to be examined to better define the interrelationship between ZMPSTE24 and various IFITM molecules.
ZMPSTE24 Protects Mice Against Influenza A Infection
ZMPSTE24 is required for endoproteolytic cleavage of prolamin A. In the absence of ZMPSTE24, animals produce excess progerin, leading to phenotypes that resemble HGPS. To experimentally overcome this phenotype, one must reduce the level of prelamin A. This can be accomplished by using animals that are heterozygous for the prelamin A gene (lmna) (Fong et al., 2004, 2006; Varela et al., 2005; Marino et al., 2008). Thus, to evaluate the in vivo relevance of ZMPSTE24 in viral restriction, lmna +/− zmpste24−/− , lmna+/− zmpste24+/− , and lmna+/− zmpste24+/+ animals were infected with a limiting dose of PR8 strain IAV. ZMPSTE24-deficient animals present signs of severe clinical illness. Overall, 65% of zmpste24 −/− animals become moribund (defined as 25% loss of body mass) compared with 20% and 0% in the zmpste24 +/− and zmpste24 +/+ groups, respectively (Fu et al., 2017). Thus, ZMPSTE24 restricts IAV infection both in vitro and in vivo.
Lungs from IAV-infected zmpste24−/− animals displayed evidence of severe interstitial pneumonia and carried an increased viral load. The histologic findings were supported by augmented levels of cytokine (interferon β, tumor necrosis factor α, interleukin 6) and chemokine (CCL2, CXCL1) transcripts in the lungs of ZMPSTE24-deficient animals (Fu et al., 2017). Thus, ZMPSTE24 deficiency is associated with enhanced pathogenesis, morbidity, and viral proliferation. The findings are consistent with an inability of zmpste24−/− mice to control influenza infection and closely parallel results observed with IFITM-deficient animals (Everitt et al., 2012; Bailey et al., 2014).
Mechanisms of ZMPSTE24 Antiviral Activity
Protease activity is essential for processing prelamin A and clearing clogged translocons. To evaluate the role of catalytic activity in viral restriction, residues associated with ZMPSTE24 catalytic activity were mutated, including those in the conserved HEXXH zinc metalloprotease motif (Pryor et al., 2013; Quigley et al., 2013). Wild-type and mutant ZMPSTE24 constructs exhibited comparable antiviral activity against IAV, VSV, and VACV infection in human cells. In addition, zmpste24−/− MEF reconstituted with human ZMPSTE24 or the E336A catalytically inactive mutant restored host restriction to IAV (Fu et al., 2017). The combined data suggest that antiviral function is independent of ZMPSTE24 protease activity.
Overexpression of ZMPSTE24 delays IAV infection, and virus accumulates in acidic endosomal-like vesicles (Fu et al., 2017). Entry assays using viral-like particles (VLP) containing the Ebola or MLV viral glycoprotein and the matrix VP40 protein fused to a beta-lactamase reporter protein were used to assess the specificity of ZMPSTE24-dependent restriction. Zmpste24−/− MEF are more susceptible to fusion with Ebola VLP. Zmpste24−/− cells are also more susceptible to infection with VLP containing IAV or VSV glycoproteins. In contrast, ZMPSTE24-deficient and wild-type cells are equally vulnerable to VLP containing the fusogenic MLV glycoprotein (Fu et al., 2017). The combined data indicate that ZMPSTE24-mediated restriction is at the level of viral entry, and specificity is determined by the fusogenic viral glycoprotein. At least three classes of viral fusion proteins have been characterized, exemplified by IAV, Sindbis, and VSV (Kim et al., 2017). ZMPSTE24, like IFITM, restricts viruses of each architectural class of fusion protein. Thus, it seems unlikely that ZMPSTE24 or IFITM directly interacts with so many structurally diverse molecules.
Genetic complementation experiments were used to establish the functional relationship between ZMPSTE24 and IFITM3. Importantly, ectopic expression of IFITM3 does not restore antiviral function in zmpste24−/− cells. However, overexpression of ZMPSTE24 can restore antiviral activity against IAV, VSV, and VACV in IFITM-deficient MEF (Fu et al., 2017). The findings imply that ZMPSTE24 is required for IFITM antiviral action. To determine whether ZMPSTE24 still exerts antiviral activity in the absence of IFITM, ZMPSTE24 was silenced in IFITM-deficient cells. Under these conditions, ZMPSTE24 knockdown uncovered an IFITM-independent restriction for VSV and VACV. The data are compatible with a model in which ZMPSTE24 serves as a downstream effector in the IFITM pathway, although an IFITM-independent pathway may also exist.
Our working model proposes that ZMPSTE24 acts in a common pathway with IFITM and is the downstream antiviral effector (Fig. 2). Without an interferon signal, basal levels of IFITM and ZMPSTE24 are expressed on endosomes to provide limited antiviral defense. In the absence of ZMPSTE24 (even with transfected IFITM), cells become more vulnerable to infection. Treatment of control cells with interferon stimulates IFITM expression, which, in turn, recruits or retains additional ZMPSTE24 to the endosomes. ZMPSTE24 alters endosomal function to impede entry of pH-dependent fusogenic viruses.
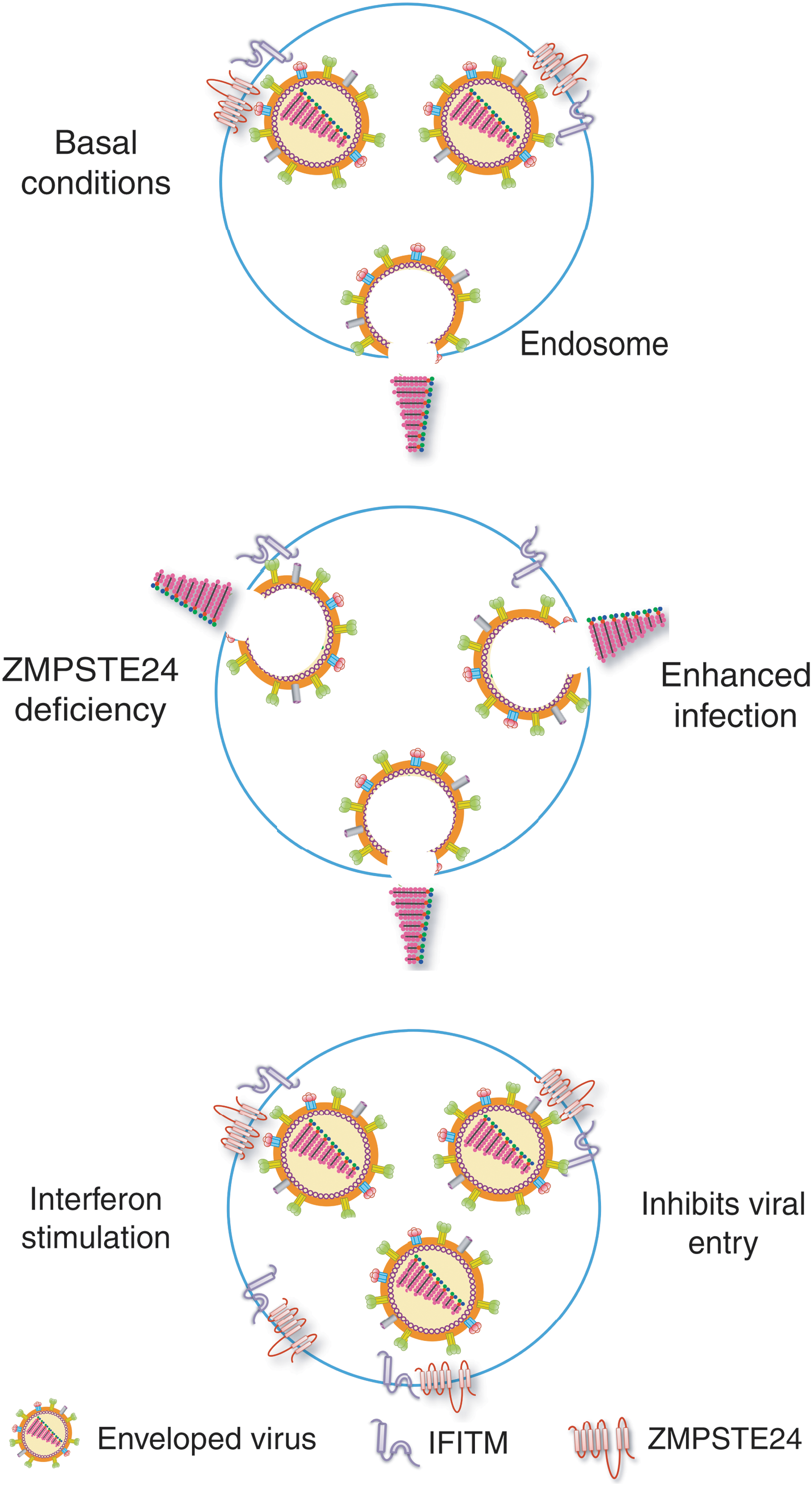
ZMPSTE24 controls IFITM antiviral pathway. In the absence of interferon, basal levels of IFITM and ZMPSTE24 are expressed in the endosome, providing limited protection against infection by fusogenic viruses. ZMPSTE24-deficient cells express IFITM but are rapidly infected, supporting a role for ZMPSTE24 as an effector of the IFITM-ZMPSTE24 antiviral pathway. Stimulation of control cells with type 1 interferons recruits additional IFITM and ZMPSTE24 to the endosome, inhibiting viral entry and trapping virus in the endosomal compartment.
Open Issues and Conclusions
(1) ZMPSTE24 is detected in IFITM protein complexes, but we do not know whether these interactions are direct. If so, the structural basis for these protein associations needs to be defined.
(2) How is ZMPSTE24 trafficking to the endosome regulated? Have some pathogens evolved strategies to subvert the IFITM-ZMPSTE24 host defense mechanism?
(3) How does the IFITM-ZMPSTE24 pathway control endosomal barrier function? Can such understanding be translated to protect against infection.
Although critical questions remain, ZMPSTE24 represents a novel intrinsic host defense molecule that impedes infection with a broad spectrum of pathogenic viruses. It acts in coordination with IFITM to hinder viruses from breaching the endosomal barrier. In mice, ZMPSTE24 protects against influenza infection. Further investigation may facilitate the development of therapeutics that mimic or stimulate ZMPSTE24 activity.
Footnotes
Authors' Contributions
Designed and analyzed experiments: M.E.D., S.L., B.F., and L.W. Performed experiments: S.L., B.F., and L.W. Wrote the article: M.E.D. Prepared the Figures: S.L.
Acknowledgment
This project is supported by NIH grant AI121288.
Disclosure Statement
No competing financial interests exist.