
Abstract
Cardiac fibrosis is closely related to multiple cardiovascular system diseases, and noncoding RNAs (ncRNAs), including long noncoding RNA (lncRNA) and microRNA (miRNA), have been reported to play a vital role in fibrogenesis. The present study aims to investigate the potential regulatory mechanism of lncRNA H19 and miR-455 on fibrosis-associated protein synthesis in cardiac fibroblasts (CFs). miRNA microarray assay revealed 34 significantly dysregulated miRNAs, including 13 upregulated miRNAs and 21 downregulated miRNAs. Among these aberrantly expressed miRNAs, we paid attention to miR-455, which was significantly downregulated in diabetic mouse myocardium and Ang II-induced CFs. Loss- and gain-of-function experiments showed that miR-455 expression levels were negatively correlated with collagen I and III expression in Ang II-induced CFs. Bioinformatic prediction programs (TargetScan, miRanda, starBase) predicted that miR-455 targeted connective tissue growth factor (CTGF) and H19 with complementary binding sites at the 3′-untranslated region, which was validated by luciferase reporter assay. Functional validation assay demonstrated that H19 knockdown could enhance the antifibrotic role of miR-455 and attenuate the CTGF expression and further decrease fibrosis-associated protein synthesis (collagen I, III, and α-SMA). The present study reveals a novel function of the H19/miR-455 axis targeting CTGF in cardiac fibrosis, suggesting its potential therapeutic role in cardiac diseases.
Introduction
M
ncRNAs are huge nonprotein-coding sequences generated from transcription, exerting vital physiological action in each level. miRNAs have been powerfully validated to modulate the fibrosis process and remodeling of cardiomyocytes. For instance, miR-21 is overexpressed in fibroblasts and enhances cardiac fibrotic remodeling and fibroblast proliferation through the CADM1/STAT3 pathway (Cao et al., 2017). lncRNA is a novel type of ncRNA, ranging 200–100 kb nucleotides in length without protein-coding capacity, participating in serious physiological processes and pathological events (Ayers and Vandesompele, 2017). Except for the affirmed key role of lncRNAs in tumorigenesis and neurological disorders, recently their multidimensional regulation in cardiovascular disorders has been recognized (Haemmig and Feinberg, 2017). For example, lncRNA myocardial infarction-associated transcript (MIAT) is remarkably upregulated and identified as the first profibrotic lncRNA in the heart in a mouse myocardial infarction model accompanied by cardiac interstitial fibrosis, revealing the role of MIAT in the pathogenesis of myocardial infarction (Qu et al., 2017). Thus, increasing evidence and findings demonstrate the emerging regulating role of lncRNAs in cardiovascular pathology.
LncRNA H19 has been verified to wildly participate in various pathological processes in tumorigenesis and tumor metastasis (Chen et al., 2016; Matouk et al., 2016). H19, an ncRNA with 3 kb length and expressed in the nucleus and cytoplasm, is activated in embryonic cells and highly expressed in embryogenesis. However, its expression is significantly decreased after birth and increased in the pathological state (Nordin et al., 2014). H19 has been shown to be involved in liver fibrosis, lung fibrosis, and renal fibrosis, including cardiac fibrosis (Tang et al., 2016; Tao et al., 2016; Xie et al., 2016). Thus, we hypothesize that H19 participates in the pathological process of MF through an uncertain pathway.
To verify the hypothesis and investigate the in-depth pathogenesis involved in miRNAs and lncRNA in MF, we detected the endogenous expression of miRNAs and lncRNAs in mouse myocardial fibroblasts and then assessed their effect on extracellular matrix accumulation.
Materials and Methods
Animal model and tissue specimens
For microarray screening analysis, 18-week-old male diabetic C57BLKS/J db/db mice and control db/m mice (paired five mice) were purchased from Qingzilan Science and Technology Ltd. (Nanjing, China). For cellular function experiments, newborn C57BL/6 mice (1–3 days) were provided by the Model Animal Research Center of Guangdong Medical University-Affiliated Hospital. Animals were housed in a humidity- and temperature-controlled room with food and water provided ad libitum. All experiments and procedures were performed according to the guidelines of the Institutional Animal Care and Use Committee of Guangdong Medical University-Affiliated Hospital. Besides, the experiment was performed in accordance with the guidelines by the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
RNA isolation and microarray analysis
Mice were anesthetized with pentobarbital sodium (50 mg/kg, intraperitoneal injection), and then their hearts were excised. RNA was isolated from each specimen using the mirVana miRNA Isolation Kit (Ambion, Austin, TX). The isolated RNA was purified with an RNeasy mini kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. The miRNA microarray was performed according to the MicroRNA Expression Profiling Assay Guide (Illumina, Inc.). The miRNAs were labeled with the miRCURY Hy3/Hy5 Power labeling kit and hybridized with the miRCURY LNA Array (version 16.0), then the labeled miRNAs were purified and hybridized to a GeneChip miRNA 4.0 Array (Affymetrix, Santa Clara, CA). After hybridization, results were analyzed using Agilent Feature Extraction software (Agilent Technologies, Waldbronn, Germany).
Cardiac fibroblast isolation and treatment
To obtain the primary cardiac fibroblasts (CFs), newborn C57BL/6 mice were sacrificed by cervical dislocation. Instantly, CFs were isolated from the myocardium as previously described (Tillmanns et al., 2015). Briefly, CFs were separated from cardiomyocytes using gravitational separation and digested with 0.1% pancreatin. CFs were cultured in DMEM supplemented with 10% FBS, 1% penicillin, and 1% streptomycin at 37°C in humid air with 5% CO2. CFs grew to confluency in the culture dish. The third-passage CFs were more suited for our further experiments. To structure the MF phenotype, partial CFs were induced with angiotensin-II (10−5 mM) for 24 h.
Cell transfection
Cell transfection was performed using small interfering RNA sequences that were supplied and synthesized by GenePharma (Shanghai, China). The transfection was performed using Lipofectamine 2000 (Invitrogen, MA) according to the manufacturer's procedure. The sequences were listed as follows: si-H19, 5′-GUACGUAACCGAAACAAUU-3′; and miR-455 inhibitor, 5′-CCCCAAGGCCAAUGACAGAUU-3′.
Quantitative real-time polymerase chain reaction analysis
Total RNA was extracted from cultured CFs using TRIzol reagent (Invitrogen, Carlsbad, CA) and quantified by NanoDrop 2000 (Thermo Fisher). Then, RNA was reverse transcribed into cDNA using the Reverse Transcription Reagent Kit (Invitrogen) according to the manufacturer's instructions. Subsequently, cDNA was amplified by SYBR Premix Ex Taq (TaKaRa, Dalian, China). All RNA levels were standardized to the GAPDH levels. The primers used in this study were listed as follows: H19, forward, 5′-CACAACGTGCTCTGCGTTGA-3′, reverse, 5′-GACTCAAGCCCTACTTGGGTA-3′; miR-455, forward 5′-GCAATAGCGTCACGCTGTCTATG-3′, reverse, 5′-GTGGTTCGGGGTAGACCGT-3′; CTGF, forward, 5′-TGGTGACCCAACCCCTATGAT-3′, reverse, 5′-GCACTTGCTTTCCTTTCTATAGTT-3′; and GAPDH, forward, 5′-GAAGGTCGGAGTCAACGGATT-3′, reverse, 5′-CGCTCCTGGAAGATGGTGAT-3′. The relative fold change was analyzed using the 2−ΔΔCt method.
Western blot analysis
Protein from CF cells was extracted using protein lysis buffer and centrifuged at 12,000 rpm for 30 min. After the protein concentration was measured, protein and sample buffer were subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). Then, the separated protein was transferred onto polyvinylidene fluoride (PVDF) membranes. PVDF membranes were blocked in 5% fat-free milk and blocked in the blocking buffer and then incubated overnight at 4°C with the primary antibody. Primary antibody (Cell Signaling Technology, Inc., Danvers, MA) involved in incubation contained rabbit anti-connective tissue growth factor (CTGF) antibody (1:1000), rabbit anti-collagen I antibody (1:1000), rabbit anti-collagen III antibody (1:1000), rabbit anti-TGF-β1 antibody (1:1000), and rabbit anti-αSMA antibody (1:1000). GAPDH acted as the internal reference. Goat anti-rabbit horseradish peroxidase-conjugated IgG (1:2000) was used as a secondary antibody. Signal was detected by chemiluminescence and images were captured on the XRS ChemiDoc system (BioRad) and analyzed by Quantity One software (BioRad).
Luciferase reporter gene assay
Fragments of the H19 gene and CTGF gene containing miR-455 binding sites, and mutant not containing binding sites, were amplified using PCR and subcloned into pGL3 luciferase promoter plasmid (Promega, Madison, WI), respectively, named pGL3-H19-WT and pGL3-H19-Mut. For luciferase reporter gene assay, HEK293 cells were cultured in 24-well culture plates and transfected with firefly luciferase vector (100 ng) and Renilla luciferase expression vector (10 ng). Renilla luciferase acted as an internal control to normalize transfection efficiencies. The combined plasmid and control plasmid were transfected into HEK293 cells using Lipofectamine-2000 (Invitrogen) according to the manufacturer's instructions. Firefly and Renilla luciferase activities were measured using the Dual-Luciferase Reporter Assay System (Promega).
RNA immunoprecipitation assay
RNA immunoprecipitation (RIP) assay was performed to confirm the relationship between H19 and miR-455. The assay was performed using the EZ-Magna RIP RNA-binding protein immunoprecipitation kit (Millipore, Billerica, MA) according to the manufacturer's instructions. CFs were lysed in RNA lysis buffer containing protease inhibitor and RNase inhibitor. Then, cells were incubated with the RIP buffer containing magnetic beads coated with antihuman argonaute 2 (Ago2) antibodies (Millipore). IgG (Millipore) was used as a negative control (input). After incubation at 4°C for 2 h, real-time polymerase chain reaction (RT-PCR) was performed to detect the enrichment of H19 and miR-455.
Statistical analysis
All data are presented as the mean ± standard error of the mean. Comparisons between groups were calculated using one-way analysis of variance and Student's t-test. A value of p < 0.05 between groups was considered statistically significant.
Results
miRNA expression profiles in diabetic mouse myocardium
To assess the miRNA expression profiles in mouse myocardium, microarray analysis was performed within diabetic C57BLKS/J db/db mice and control db/m mice. Heat maps exhibited the 34 dysregulated miRNAs, including 13 upregulated miRNAs and 21 downregulated miRNAs with twofold changes (Fig. 1A). Among these aberrantly expressed miRNAs, we paid attention to miR-455, whose function in fibroblasts was unclear, and chose it as our target. Afterward, the expression of miR-455 in diabetic mouse myocardium and extracted CFs was detected using RT-PCR to validate the microarray analysis results. As expected, miR-455 expression was significantly decreased in both diabetic mouse myocardium and Ang II-induced CFs (Fig. 1B, C). The above results revealed the downregulation of miR-455 and verified the consistent phenotype in diabetic mouse myocardium and CFs.

miRNA expression profiles detected by microarray analysis showed the dysregulated miRNAs in diabetic mouse myocardium. miRNA-455 was significantly downregulated.
miR-455 overexpression decreased fibrous protein secretion in CFs and CTGF was the target gene of miR-455
Microarray analysis had revealed the aberrantly expressed miRNA expression profiles; moreover, we selected the lower expressed miR-455 as the research target. Besides, the miR-455 expression level was markedly upregulated or downregulated in CFs transfected with miR-455 mimics or inhibitors (Fig. 2A). Because miR-455 was significantly decreased in CFs induced by Ang II, to assess its regulation of extracellular matrix accumulation, we performed loss- and gain-of-function experiments to measure the expression levels of collagen I and III. As shown in Figure 2B–D, mRNA and protein expression levels of collagen I and III were significantly increased in CFs induced by Ang II. Nevertheless, miR-455 mimics and inhibitors effectively inhibited or enhanced the mRNA and protein expression levels of collagen I and III. Bioinformatic analysis and computational prediction programs (TargetScan, miRanda, PicTar, and miRGen) were performed to predict the potential binding sites for miR455 in the 3′-UTR (untranslated region) of CTGF (Fig. 2E). Afterward, the luciferase reporter assay validated the predictive 3′-UTR binding within miR455 and CTGF (Fig. 2F). Moreover, miR-455 mimics significantly suppressed the mRNA and protein expression levels of CTGF, which were recovered by miR-455 inhibitors. In summary, comprehensive data revealed that miR-455 acted as an antifibrogenic factor in MF, and the regulatory pathway might target CTGF.
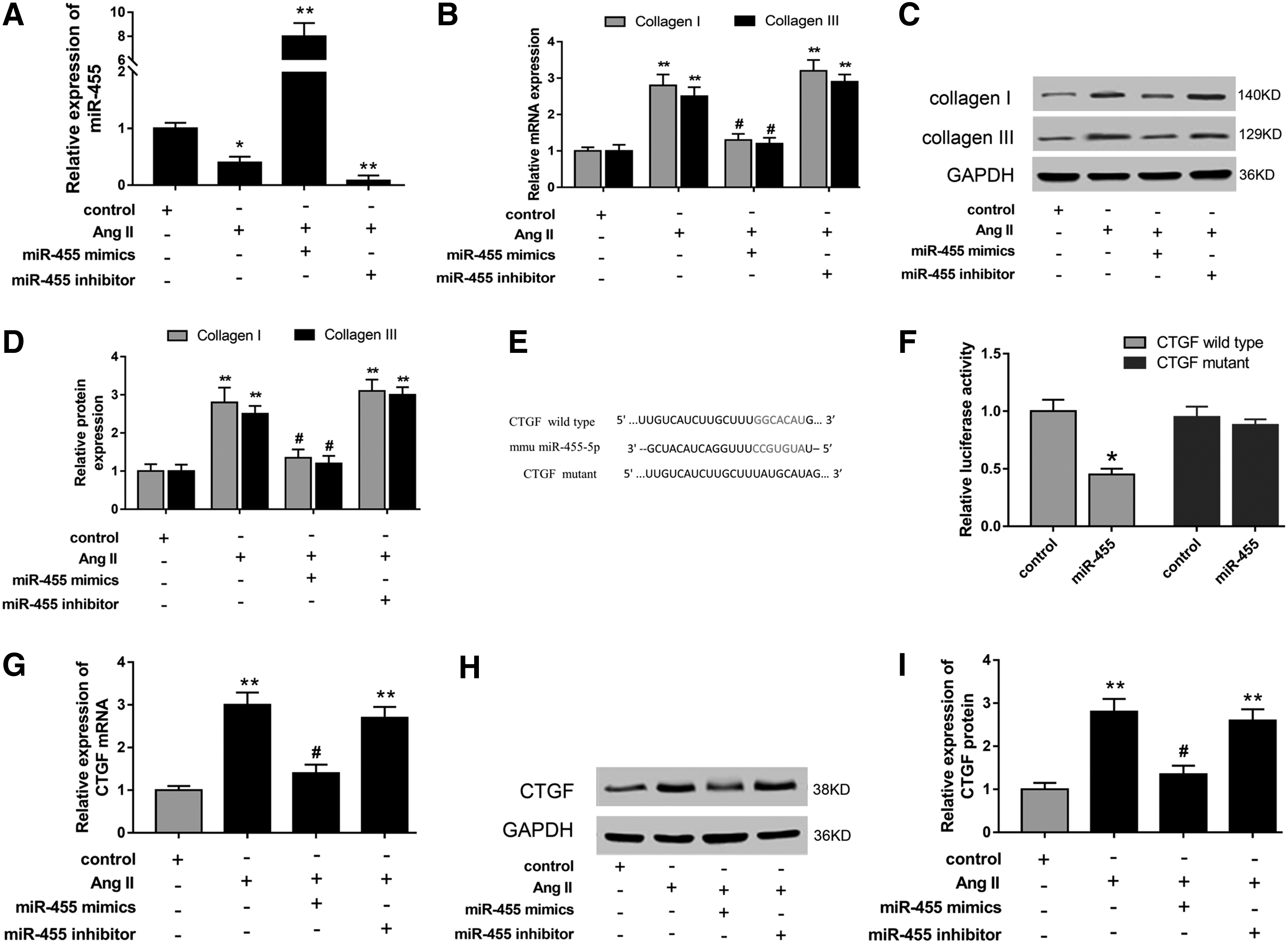
miR-455 overexpression decreased fibrous protein secretion in CFs. Besides, CTGF was the target gene of miR-455.
LncRNA H19 was the target of miR-455
Because lncRNA had been reported to play an important role in cardiac fibrosis, we predicted and confirmed the potential functional lncRNA using the bioinformatic computational program. Fortunately, we found that lncRNA H19 shared complementary binding sites with miR-455 at 3′-UTR (Fig. 3A). The dual-luciferase reporter assay showed that the combination of H19 and miR-455 could decrease luciferase activities, indicating the complementary binding at 3′-UTR (Fig. 3B). Based on the interaction, the expression of H19 was detected using RT-PCR and found to be significantly upregulated in db/db diabetic mouse myocardium compared with db/m mice (Fig. 3C). RIP assay was performed to confirm the interaction between H19 and miR-455. RIP assay showed that H19 and miR-455 were significantly enriched in Ago2-containing beads compared with the input group (Fig. 3D). For cell validation experiment in CFs induced by Ang II and transfected with miR-455 mimics and inhibitors, H19 was overexpressed in Ang II-induced CFs and negatively correlated with miR-455 expression levels (Fig. 3E). Then, interference sequence-targeted H19 (si-H19) could markedly knock down the expression (Fig. 3F). Moreover, the expression of miR-455 was significantly recovered after transfection with si-H19 in Ang II-induced CFs (Fig. 3G). Overall, serious experiments discovered and validated the target relationship and negative regulation between miR-455 and H19, providing a feasible approach to probe the modulation of lncRNA in cardiac fibrosis.
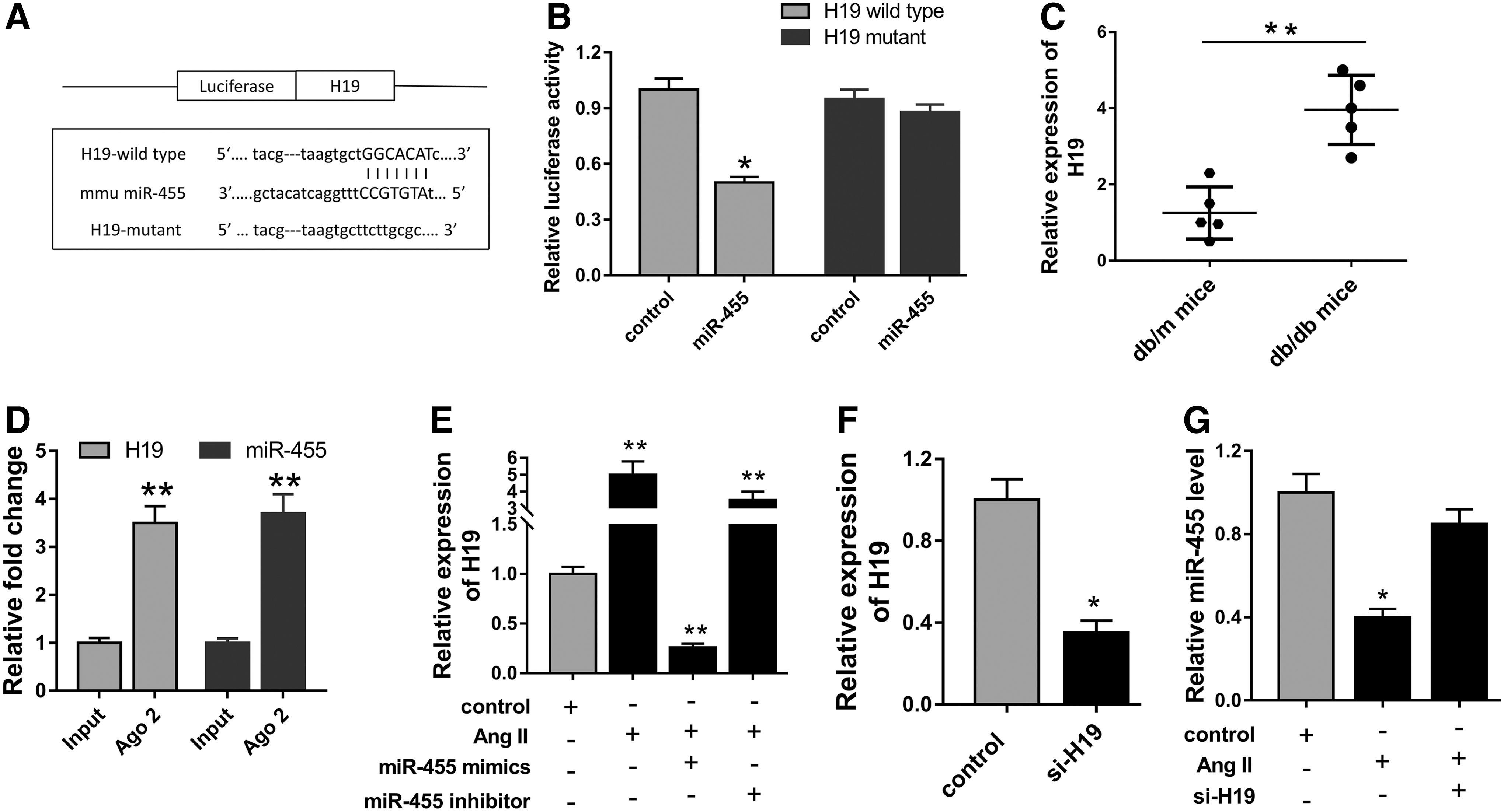
LncRNA H19 was the target of miR-455.
H19 and miR-455 modulated myocardial extracellular matrix accumulation through targeting CTGF
The present study had uncovered the antifibrogenic role of miR-455 and its target CTGF; besides, H19 was also targeted with miR-455. Hence, we hypothesized that there was in-depth function of the regulatory pathway of H19, miR-455, and CTGF in myocardial extracellular matrix accumulation. In further cell verification experiments, we performed loss-of-function experiments to investigate the mRNA and protein expression of CTGF and fibrosis-associated proteins in CFs. As shown in Figure 4A–C, mRNA and protein of CTGF were suppressed in CFs transfected with si-H19, while miR-455 inhibitor restored the expression. To measure the modulation of H19, miR-455, and CTGF of myocardial extracellular matrix accumulation, we detected the fibrosis-associated protein (collagen I, III, and α-SMA) expression in CFs transfected with si-H19, miR-455 inhibitor, and CTGF siRNA. Protein and mRNA expression detected with western blot and RT-PCR showed that collagen I, collagen III, and α-SMA expression levels were significantly upregulated in Ang II-induced CFs (Fig. 4D–I). Moreover, H19 knockdown induced by si-H19 and lower CTGF expression induced by siRNA markedly suppressed the mRNA and protein levels of collagen I, collagen III, and α-SMA, while miR-455 inhibitor and CTGF siRNA could reverse the suppression. Hence, a series of loss-of-function experiments and rescue experiments strongly revealed that H19 modulated myocardial extracellular matrix accumulation through negative regulation of miR-455 by targeting CTGF, validating the hypothesis proposed in the preamble.
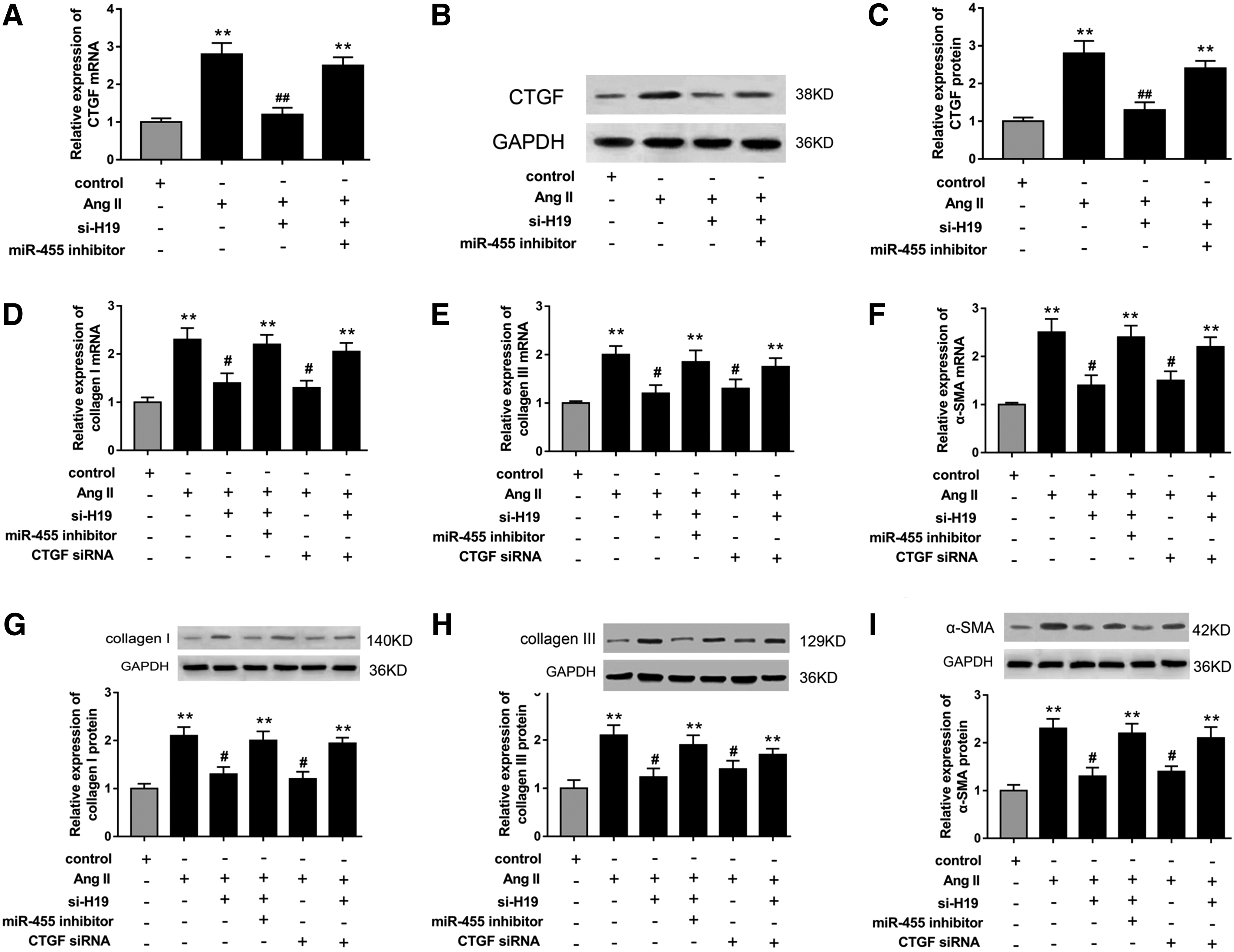
H19 and miR-455 modulated myocardial fibrosis-associated protein accumulation through targeting CTGF.
Discussion
ncRNAs have been verified to participate in the modulation of cardiovascular system disease, including atherosclerosis, cardiac remodeling, and MF (Lorenzen et al., 2015; Piccoli et al., 2016). MF is the ultimate outcome of multiple myocardial damage progression. Being different from miRNA, whose roles in the pathological process of MF have been widely probed, lncRNAs have been increasingly recognized for their vital roles in pathogenesis of cardiovascular diseases, especially MF (Huang et al., 2016). In the present study, we investigate the in-depth roles of lncRNA and miRNA in MF.
Although numerous miRNAs had been measured and verified in MF pathogenesis, to discover the novel functional miRNA in fibrotic processes, we performed miRNA microarray to screen dysregulated miRNAs and found that miR-455 was significantly downregulated in diabetic mouse myocardium. Loss- and gain-of-function experiments validated that miR-455 overexpression suppressed fibrosis-associated protein synthesis, indicating the antifibrogenic role in MF. As is known to all, miRNAs exert physiological and pathological regulations through targeting functional gene mRNAs to silence protein expression. Thus, we uncovered the potential target mRNA using bioinformatic analysis and found CTGF bound with miR-455 at 3′-UTR, which was validated by luciferase reporter assay. For instance, miR-22 silencing induced with AMO-22 upregulated Col1α1 and Col3α1 expression and increased fibrogenesis; besides, TGFβRI is a direct target for miR-22 and mediated the antifibrotic effect of miR-22 (Hong et al., 2016). In addition, miR-155 deficiency (miR-155 knockout mice) could prevent cardiac fibrosis induced by diabetes and attenuate collagen synthesis through the TGF-β1-Smad 2 signaling pathway in high-glucose-induced CFs (Zhang et al., 2016). Thus, we validate the antifibrosis of miR-455 through targeting CTGF, providing a novel miRNA/mRNA pathway in cardiac fibrogenesis.
The function of miR-455 and its target gene CTGF in MF secretion had been verified. Because lncRNAs have been reported to play an important role in cardiac fibrosis, being encouraged with significant miR-455/CTGF regulation, we sequentially investigate the possible functional lncRNA in CF fibrogenesis. Fortunately, with the assistance of bioinformatic analysis, we found that lncRNA H19 shared complementary binding sites with miR-455, which was validated by luciferase reporter assay. Tao et al. (2016) reported that H19 endogenous expression is overexpressed in CF and fibrosis tissues and controls the DUSP5/ERK1/2 axis to modulate CF proliferation and fibrosis. Furthermore, H19 encoded miR-675 and is upregulated in both normal and diseased hearts in pathological cardiac hypertrophy and heart failure; besides, CaMKIIδ was a direct target of miR-675 and partially mediated the effect of H19 on cardiomyocyte hypertrophy (Liu et al., 2016). Comprehensively, consistent with previous researches, H19 plays an important role in fibrosis-associated protein synthesis in diabetic CFs.
Up to now, lncRNAs have been reported to participate in the regulation of cardiac fibrosis pathogenesis induced with myocardial infarction and myocardial hypertrophy; however, the authentic action mechanism is still unclear (Thum, 2014). One of the major regulating patterns is that they act as miRNA sponges to lower target miRNAs and further alleviate functional protein secretion. In postinfarct myocardium, lncRNA MIAT is upregulated and miR-24 is downregulated; meanwhile, miR-24 downregulation increases expression of Furin and TGF-β1 (Qu et al., 2017). In cardiac hypertrophy, lncRNA cardiac hypertrophy-related factor acts as an endogenous sponge of miR-489 and downregulates miR-489 expression levels and regulates target Myd88 expression (Wang et al., 2014). Besides, lncRNA cardiac hypertrophy-associated transcript is detected using global lncRNA expression profiling, modulating cardiomyocyte hypertrophy (Viereck et al., 2016).
In summary, the present study reveals the crucial regulation of H19 in fibrogenesis in diabetic CFs through binding miR-455 targeting CTGF, suggesting the role of lncRNA/miRNA/mRNA regulation pathway in cardiac fibrosis.
Footnotes
Acknowledgments
This work was supported by Affiliated Hospital of Guangdong Medical University. The experimental resources were provided by Basic Medical Center of Guangdong Medical University. The authors thank Dr. Li, PhD, for technical guidance and English language editing.
Disclosure Statement
No competing financial interests exist.