
Abstract
The enrichment of CD20+ and CD138+ immune cells were previously associated with improved survival in colorectal cancer (CRC). We previously discovered that the resected tumors in CRC patients were highly enriched with interleukin (IL)-10-producing CD19loCD27hi plasmablasts with potential suppressor functions. It is still unknown what roles the CD19loCD27hi plasmablasts play in CRC patients. In this study, we first demonstrated that B cells from peripheral blood mononuclear cells (PBMCs) could be stimulated to resemble tumor-infiltrating plasmablasts using a coculture containing Caco-2 and heat-killed bacteria. The PBMC-derived CD19loCD27hi plasmablasts and tumor-infiltrating plasmablasts contained comparable frequencies of IL-10-expressing cells and secreted similar levels of IL-10. We later found that these CD19loCD27hi plasmablasts significantly suppressed the mRNA and cytokine expression of IL-17A in PBMCs, as well as the expression of RAR-related orphan receptor gamma t (RORγt) in CD4+ T cells. This suppressive effect did not involve the induction of Foxp3+ regulatory T cells, since no upregulation of Foxp3 level was observed. Through IL-10/IL-10R blocking and exogenous IL-10 experiments, we found that these CD19loCD27hi plasmablasts primarily mediated IL-17A suppression through IL-10 production. Other B cell-related mechanisms might also contribute to this inhibitory effect. In our cohort of patients, patients with high frequency of tumor-infiltrating IL-10+ CD19loCD27hi plasmablasts presented low IL-17A+ CD4+ T cell frequency and better survival. Altogether, these results suggested that CD19loCD27hi plasmablasts with Breg functions were associated with better prognosis in CRC, possibly by suppressing harmful Th17 inflammation.
Introduction
R
A number of studies have examined the role of B cells and Breg cells in colorectal cancer (CRC). A study by Shimabukuro-Vornhagen et al. (2014) demonstrated that the circulating and tumor-infiltrating B cells in CRC patients were enriched with memory B cells (IgD−CD27+) as well as plasma cells (CD20−CD38hi). One of the regulatory B cell subsets, with characteristic CD24hiCD38hi surface marker expression (Blair et al., 2010), presented decreased numbers in the CRC patients' peripheral blood, but was elevated with progressive tumor stages in resected tumor samples. These cells were also found at significantly higher levels in metastatic tumors compared with primary tumors (Shimabukuro-Vornhagen et al., 2014), suggesting an involvement of Breg cells in CRC. We, in a previous study (Mao et al., 2017), examined tumor-infiltrating B cells in CRC patients. We found that a group of distinct Breg cells, with CD19loCD27hi plasmablast phenotype (Matsumoto et al., 2014), made up a large percentage of lymphocytes in resected tumors from CRC patients. These B cells produced high levels of IL-10, but not TGF-β, could suppress interferon (IFN)-γ and tumor necrosis factor (TNF)-α from T cells, but did not promote Foxp3 expression. The effect of this B cell subset in CRC prognosis is unknown. In another study by Berntsson et al. (2016), immune cell-specific CD20 and CD138 expression was significantly correlated with overall survival in CRC patients, but the underlying mechanisms associated with this observation remain unclear.
In this study, we followed up our previous examinations on the role of CD19loCD27hi plasmablast in CRC.
Materials and Methods
Subjects
The study was approved by the Institutional Ethics Board of the Chinese PLA General Hospital. After receiving informed consent, we collected peripheral blood and tumor samples from all 10 male CRC patients between the ages of 55 and 71 years. The diagnosis, treatment, and sample collection of all patients were performed at the Chinese PLA General Hospital using DICAT (Vancouver, Canada). All patients during the sample collection stage were treated by surgical removal only, without concurrent chemotherapy or radiation therapy before surgery. Follow-ups started immediately after surgery and continued for at least 3 years or until death.
Sample processing
Peripheral blood samples were collected 1–2 weeks before surgery. Peripheral blood mononuclear cells (PBMCs) were collected by Ficoll centrifugation at 350 g for 30 min without braking, followed by two washings at 350 g for 5 min in standard culture medium (RPMI 1640 with
Cell culture
The Caco-2 cell–PBMC coculture was setup as previously described with minor modifications (Lea, 2015). Specifically, 300 μL per well of a 1 × 106/mL Caco-2 cell suspension were seeded on a 24-well 3.0-μm pore-size insert (Corning). The insert was then placed in a plate and incubated for 14 days at 37°C, 5% CO2, and 100% humidity for 14 days. One hundred fifty microliters of the upper culture medium was replaced every other day. At day 14, 1 × 106/mL PBMCs were added to the bottom chamber. Staphylococcus aureus Cowan (SAC) cells (EMD Millipore) were washed three times in phosphate-buffered saline and added to PBMCs at 1:10,000 dilution factor.
For all other cocultures, each cell type was added at 1:1 ratio with each other, to a final concentration of 1 × 106/mL. All cell cultures were maintained at 37°C, 5% CO2, and 100% humidity until analysis. For cultures containing CD4+ T cells, anti-human CD3/CD28 antibodies were added at 3 μg/mL each.
Flow cytometry
PBMCs or tumor mononuclear cells were incubated with FcX FcR blocker (BioLegend) for 5 min at room temperature, and then with Violet Dead Cell Stain (Invitrogen) and various combinations of the following anti-human antibodies: CD3, CD4, CD19, and CD27 (all from BioLegend), for 30 min on ice. Cells were washed twice to remove excess antibodies. For intracellular staining, cells were incubated in Cytofix/Cytoperm (BD) for 20 min on ice, washed in 1× Perm Wash (BD), and then incubated with various combinations of the following anti-human antibodies: IL-10, IL-17A, TGF-β, Foxp3 (all from eBioscience), and RORγt (R&D Systems) for 30 min on ice. Isotype control antibodies were used for the staining and gating of each intracellular/nuclear antigen. Cells were washed twice in 1× Perm Wash, and fixed with 2% formaldehyde. Sample acquisition was performed in FACS Canto cytometer (BD) and the results were analyzed in FlowJo.
Cell subset isolation
The isolation of B cells/plasmablasts or CD4+ T cells from PBMCs, tumor mononuclear cells, and cocultures were performed using magnetic selection with materials from the Human B Cell Enrichment Kit or the Human CD4+ T Cell Enrichment Kit (both from STEMCELL), respectively. For the removal of plasmablasts, the anti-human CD27 monoclonal antibody was added to the antibody cocktail in the B Cell Enrichment Kit.
Quantitative real-time PCR
Total RNA was extracted from isolated CD4+ T cells using TRIzol (Invitrogen). cDNA was synthesized using the SuperScript IV reverse transcriptase system (Thermo Fisher). Quantitative real-time PCR was performed using the QuantStudio 3 Real-Time PCR System (Applied Biosystems) and the SYBR Green Master Mix (Thermo Fisher) following the manufacturer's instructions. Primer pairs used were human Foxp3 forward CTGCCCCTAGTCATGGTGG, human Foxp3 reverse CTGGAGGAGTGCCTGTAAGTG, human RORγt forward GTAACGCGGCCTACTCCTG, human RORγt reverse GTCTTGACCACTGGTTCCTGT-3, and commercial primer pairs for IL-17A, IL-21, and IL-22 from SinoBiological. The 2–ΔΔCT method was used to calculate mRNA levels for each gene (Livak and Schmittgen, 2001).
Statistical analyses
All statistical analyses were performed in the Prism 6 software (GraphPad). Lines and error bars represent mean ± standard deviation. p Values <0.05 were considered significant.
Results
SAC-induced plasmablasts partially resembled tumor-infiltrating plasmablasts in cytokine secretion
We previously found that colorectal tumors were enriched with CD19loCD27hi plasmablasts with high IL-10, but low TGF-β secretion and potential to suppress inflammation in the tumor microenvironment (Mao et al., 2017), but the lack of cells prohibited further phenotypical and functional characterization. Therefore, in this study, we first examined whether the tumor-infiltrating plasmablasts could be recreated using B cells from PBMCs. To simulate the microenvironment of a tumor, we cocultured PBMCs from CRC patients with Caco-2 cells, a common human CRC line, with additional heat-killed SAC cells added in cell culture. At day 0 of coculture, little plasmablasts were found in PBMCs. However, after 6 days, we were able to recover a large fraction of B cells differentiating into CD19loCD27hi plasmablasts (Fig. 1A). Similar to tumor-infiltrating plasmablasts, the PBMC-derived plasmablasts also demonstrated high IL-10, but low TGF-β expression (Fig. 1B, C). No significant differences in the percentage of IL-10- or TGF-β-expressing cells were found between tumor-infiltrating plasmablasts and PBMC-derived plasmablasts (Fig. 1C). We also examined the amount of IL-10 and TGF-β secretion from sorted B cells/plasmablasts from tumor-infiltrating mononuclear cells and SAC-stimulated PBMCs by enzyme-linked immunosorbent assay (ELISA). At the same cell number, the tumor-infiltrating B cells/plasmablasts and B cells/plasmablasts from SAC-stimulated PBMCs secreted comparable levels of IL-10 (Fig. 1D). Both cell types did not express high levels of TGF-β. The removal of SAC diminishes the formation of CD19loCD27hi plasmablasts to near-zero levels, and the removal of Caco-2 cell line in coculture significantly reduced the level of IL-10 secretion by B cells/plasmablasts (data not shown).
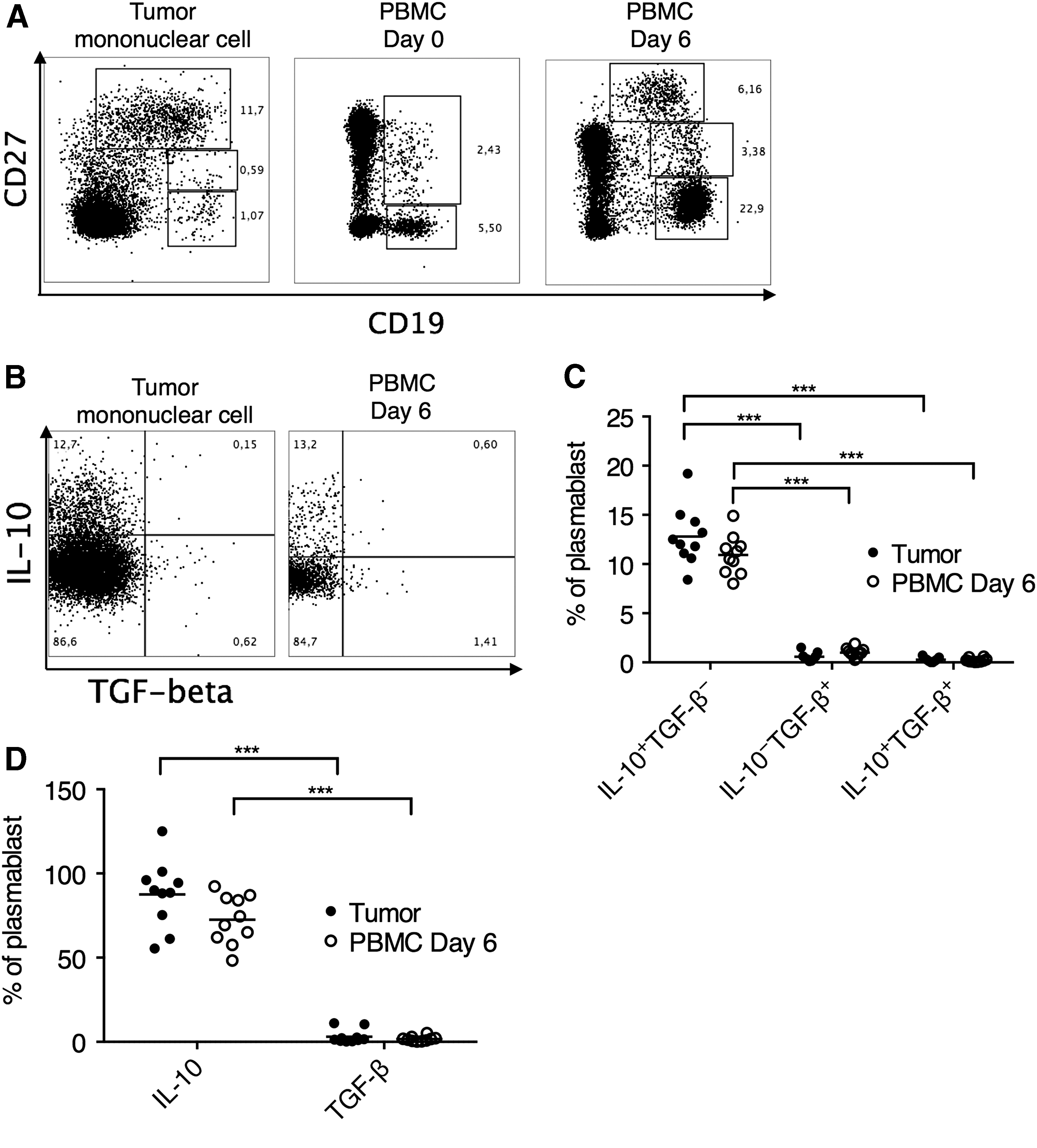
SAC-induced PBMC-derived plasmablasts partially resembled tumor-infiltrating plasmablasts in cytokine secretion.
Plasmablast suppressed IL-17A expression
Having shown that SAC stimulation and Caco-2 coculture enabled the derivation of CD19loCD27hi plasmablasts, which resembled tumor-infiltrating plasmablasts in functional cytokine secretion, we next examined the potential effects of these plasmablasts on CRC-related inflammation. It has been shown by many studies that enrichment of Th17 cells in CRC patients is associated with poor prognosis (De Simone et al., 2013). The cytokines secreted by Th17 cells, including IL-17, IL-21, and IL-22, have been associated with various tumor-promoting roles. IL-17 and IL-22 signaling activate proliferative pathways in epithelial cells through STAT3 and NF-κB (West et al., 2015). IL-17 also promotes tumor vascularization through increasing vascular endothelial growth factor (VEGF) and ProstaglandinE 2 (PGE2) production (Numasaki et al., 2003), whereas high level of IL-22 promotes the growth of human CRC cell lines in immunodeficient mice (Jiang et al., 2013). IL-21, a promoter of Th1 and Th17 responses, is increased in the inflamed tissues of ulcerative colitis and patients with colitis-associated CRC.
We, therefore, examined the effect of plasmablasts on Th17 inflammation. We incubated CD4+-sorted T cells in the presence of SAC-stimulated B cells/plasmablasts or unstimulated B cells from PBMCs. A significantly lower level of IL-17A mRNA expression was found in CD4+ T cells when SAC-stimulated B cells/plasmablasts were present (Fig. 2A). We also found that in the supernatant, the level of total IL-17A secretion was significantly reduced when SAC-stimulated B cells/plasmablasts were present (Fig. 2B). To confirm that the regulatory functions were mediated by CD19loCD27hi plasmablasts and not by other B cell subsets in the SAC-stimulated B cells/plasmablasts, we used an anti-CD27 monoclonal antibody to deplete CD19loCD27hi plasmablasts in the SAC-stimulated B cell/plasmablast mixture, and the inhibitory effect on IL-17A was no longer present (Fig. 2A, B). Another possible explanation to these observations was that unstimulated B cells presented higher IL-17A expression than SAC-stimulated B cells/plasmablasts, resulting in higher total IL-17A in the supernatant (Fig. 2B). To exclude this possibility, we repeated this experiment with unstimulated B cells and SAC-stimulated B cells/plasmablasts only, and found that neither B cell group presented substantial expression of IL-17A, IL-21, or IL-22. Contrary to the alternative explanation, unstimulated B cells had lower IL-17A expression than SAC-stimulated B cells/plasmablasts (Fig. 2C). No significant difference in IL-21 and IL-22 expression between the two groups was observed. Altogether, these data demonstrated that compared with B cells directly from PBMCs, the SAC-stimulated plasmablasts presented significant suppressive function in IL-17A expression.
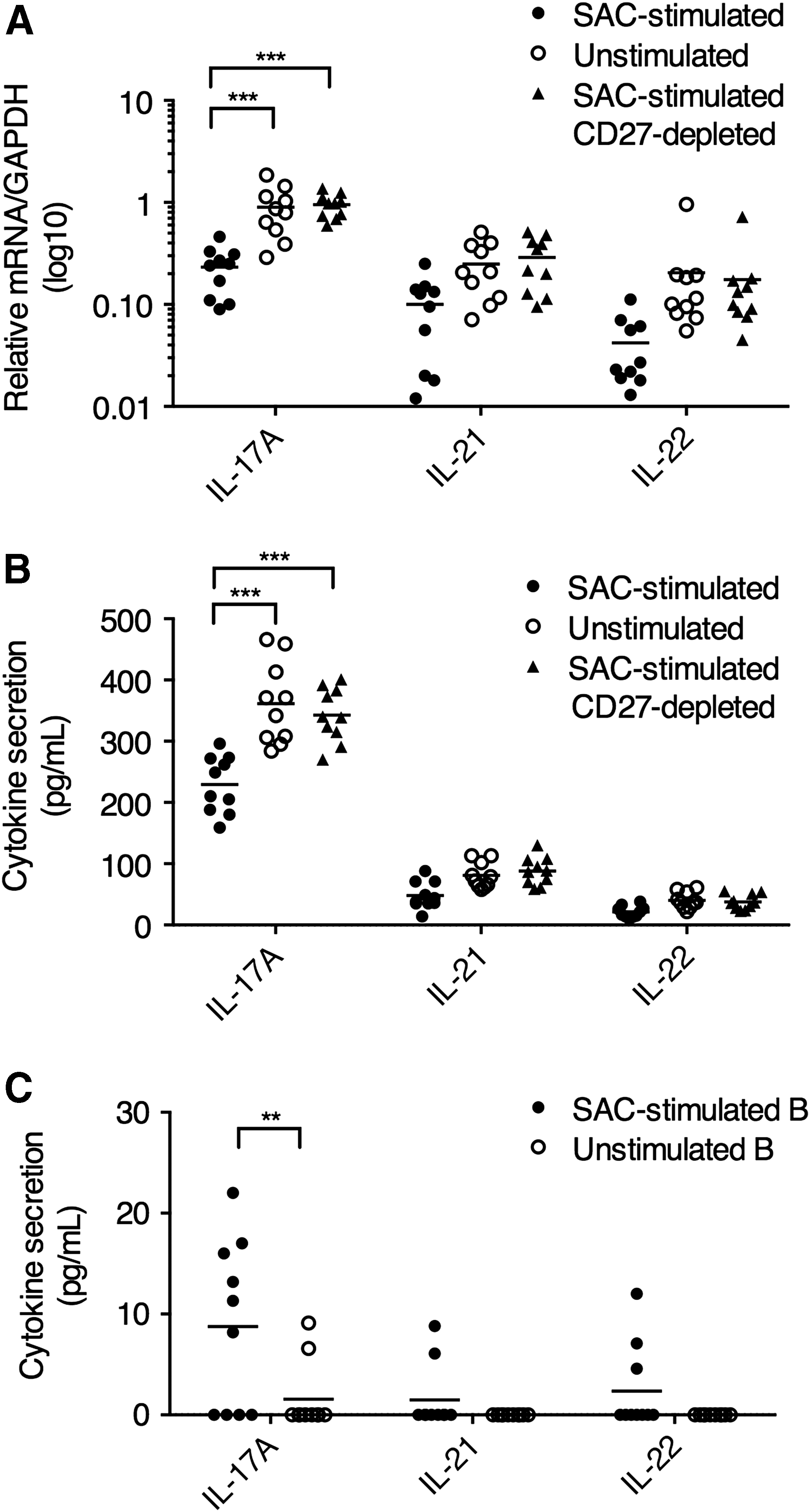
SAC-stimulated plasmablasts suppressed IL-17A expression. By negative enrichment, CD4+ T cells were isolated from PBMCs and B cells/plasmablasts were isolated from PBMCs (unstimulated) or SAC-stimulated Caco-2-cocultured PBMCs (SAC stimulated). For CD19loCD27hi plasmablast depletion (SAC stimulated CD27 depleted), an anti-CD27 monoclonal antibody was added to the antibody cocktail during negative enrichment. The CD4+ T cells and B cells/plasmablasts were then cocultured at 1:1 ratio for 72 h. The CD4+ T cells were then isolated from the coculture by magnetic selection for
Plasmablasts suppressed the expression of RORγt in CD4+ T cells
To explain the suppressive effects of plasmablasts on IL-17 production from CD4+ T cells, we examined the effect of plasmablasts on the expression of RORγt, the canonical Th17 transcription factor. By flow cytometry staining, we found that CD4+ T cells with SAC-stimulated plasmablasts demonstrated significantly lower RORγt expression than those with unstimulated B cells (Fig. 3A, B). We also found that CD4+ T cells with SAC-stimulated plasmablasts presented significantly lower RORγt mRNA transcription than those with unstimulated B cells (Fig. 3C).
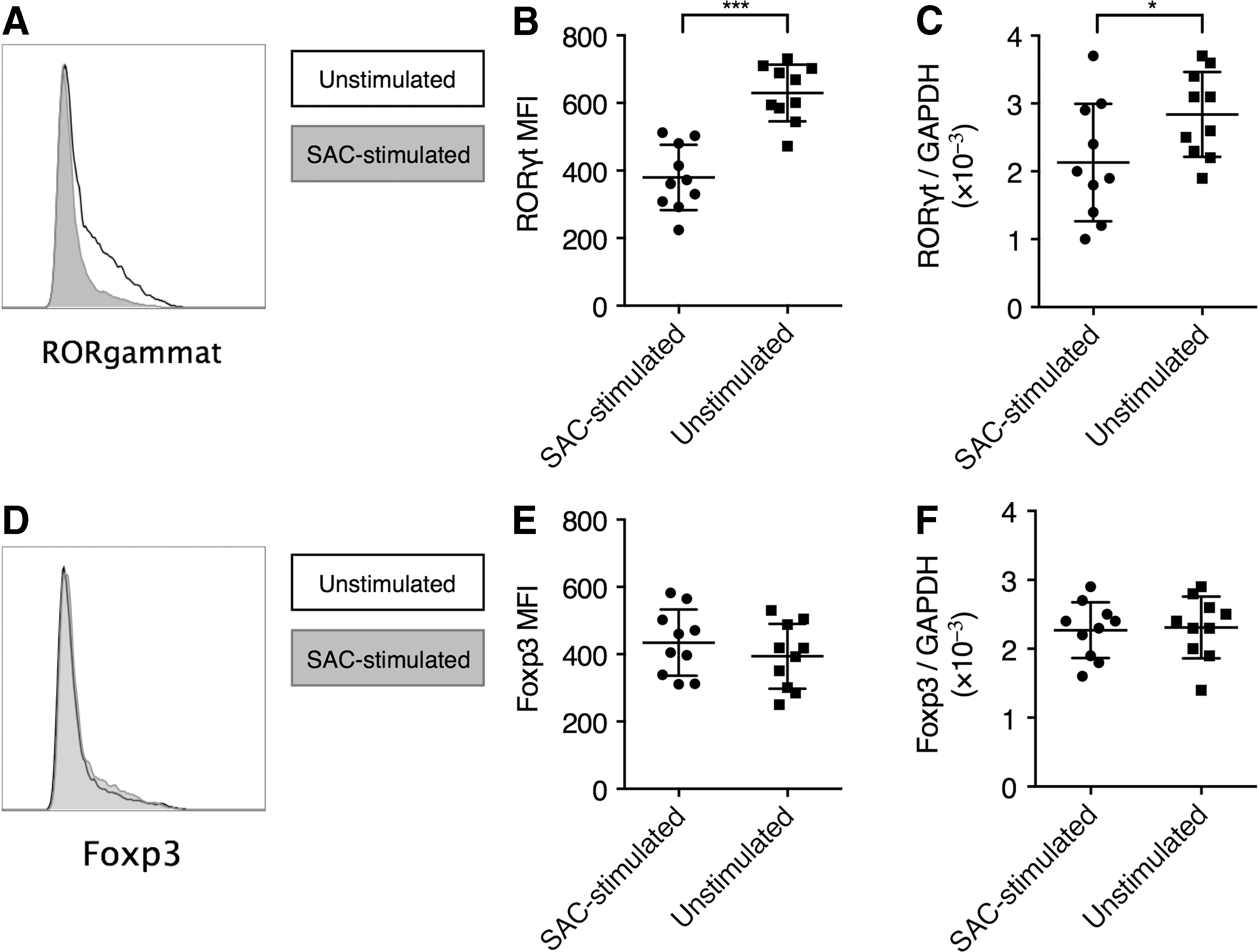
SAC-stimulated plasmablasts suppressed RORγt expression. CD4+ T cells and SAC-stimulated plasmablasts or unstimulated B cells were cocultured as described in Figure 2. The cells were then split into two fractions, with one fraction stained for intracellular ROR c expression in flow cytometry, and the CD4+ T cells isolated from the other fraction and processed for mRNA analyses.
Several previous studies demonstrated that Breg cells play a critical role in the maintenance of Foxp3+ Treg cells. Therefore, it is also possible that plasmablasts could inhibit Th17 inflammation through promoting Treg development. We analyzed this possibility by examining the expression of Foxp3 in CD4+ T cells. We did not find that SAC-stimulated plasmablasts significantly altered the expression of Foxp3, either by flow cytometry or by mRNA analysis (Fig. 3D–F).
Plasmablast mediated suppressive effects primarily through the expression of IL-10
Given that the SAC-stimulated plasmablasts highly expressed IL-10 but not TGF-β, and did not induce Foxp3 expression in vitro, we postulated that the plasmablasts possibly mediated IL-17A and RORγt suppression through IL-10-mediated pathways. To examine this possibility, we repeated the SAC-stimulated plasmablast CD4+ T cell coculture experiment with conditions to manipulate the IL-10 pathway. Blocking IL-10 and IL-10 receptor (IL-10R) resulted in significantly elevated IL-17A expression (Fig. 4A). Using unstimulated B cells with IL-10 supplement in place of SAC-stimulated plasmablasts also caused significantly reduced IL-17A, when IL-10 was added at moderate (10 ng/mL) or high (100 ng/mL) concentrations (Fig. 4B). In the absence of B cells/plasmablasts, IL-10 supplement could significantly reduce IL-17A secretion only at high (100 ng/mL) concentrations (Fig. 4C). Altogether, these data demonstrated that SAC-stimulated plasmablasts primarily mediated IL-17A inhibitory effects through IL-10, but other B cell/plasmablast-associated mechanisms could also contribute to IL-17A suppression.
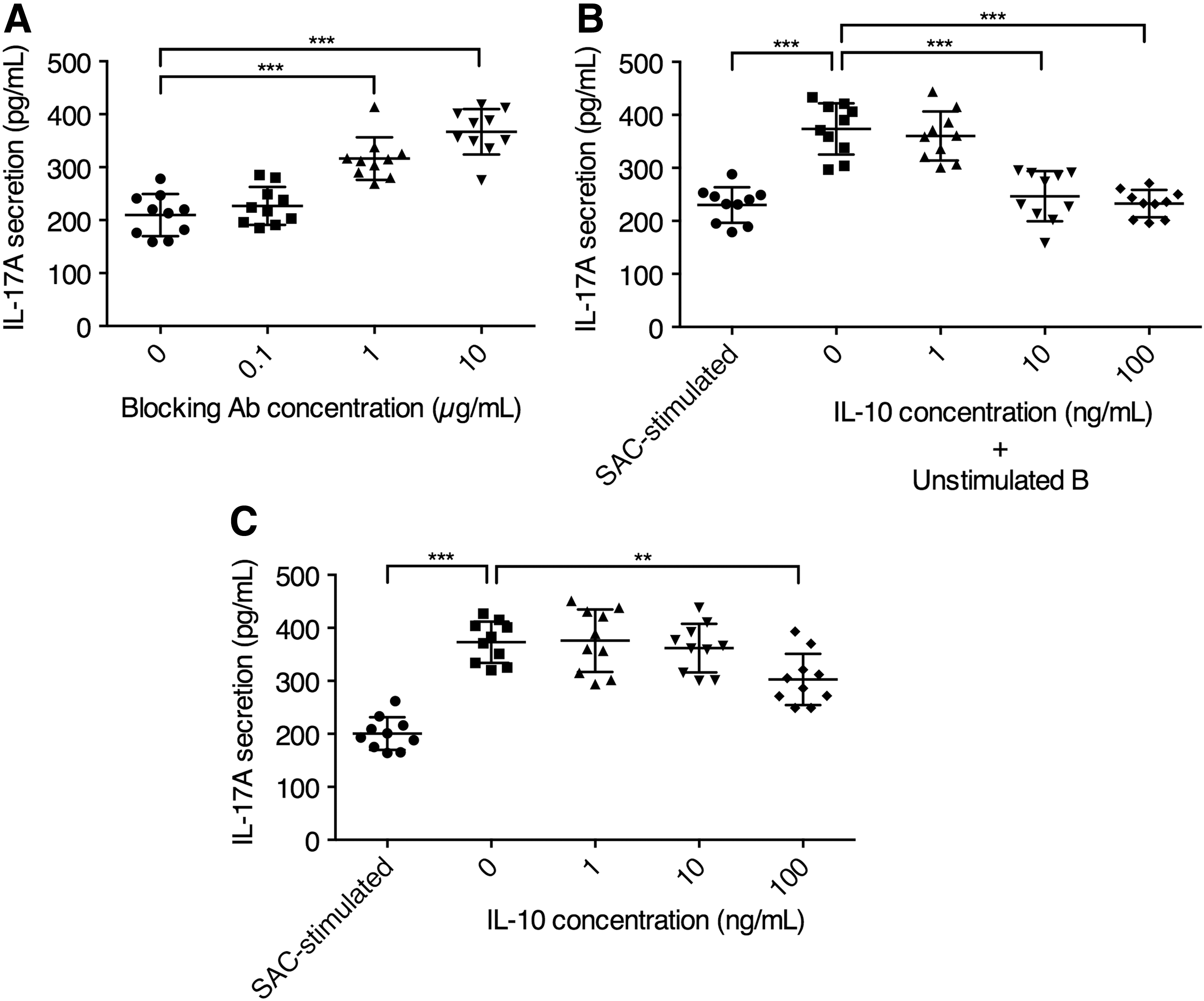
SAC-stimulated plasmablasts primarily suppressed IL-17A expression through IL-10.
Patients with high intratumoral plasmablasts presented low Th17 cells and better survival
Previous investigations suggested that the level of immune cell-specific CD20 and CD138 expression was associated with better prognosis in CRC patients (Berntsson et al., 2016). In this study, we examined the association between intratumoral IL-10+ plasmablast frequency and IL-17A+CD4+ T cell frequency. Patients were grouped into high IL-10+ and low IL-10+ groups, using the median frequency of tumor-infiltrating IL-10+TGF-β− plasmablasts as a division standard. We found that high IL-10+ patients presented significantly lower intratumoral frequencies of IL-17A+CD4+ T cells (Fig. 5A). In the six patients that were alive 5 years after initial tumor resection, the frequency of tumor-infiltrating IL-10+ plasmablasts was significantly higher than the four deceased patients (Fig. 5B). Altogether, these results demonstrated that in our cohort, CRC patients with high tumor-infiltrating plasmablasts presented low IL-17A expression in the tumor and better survival.
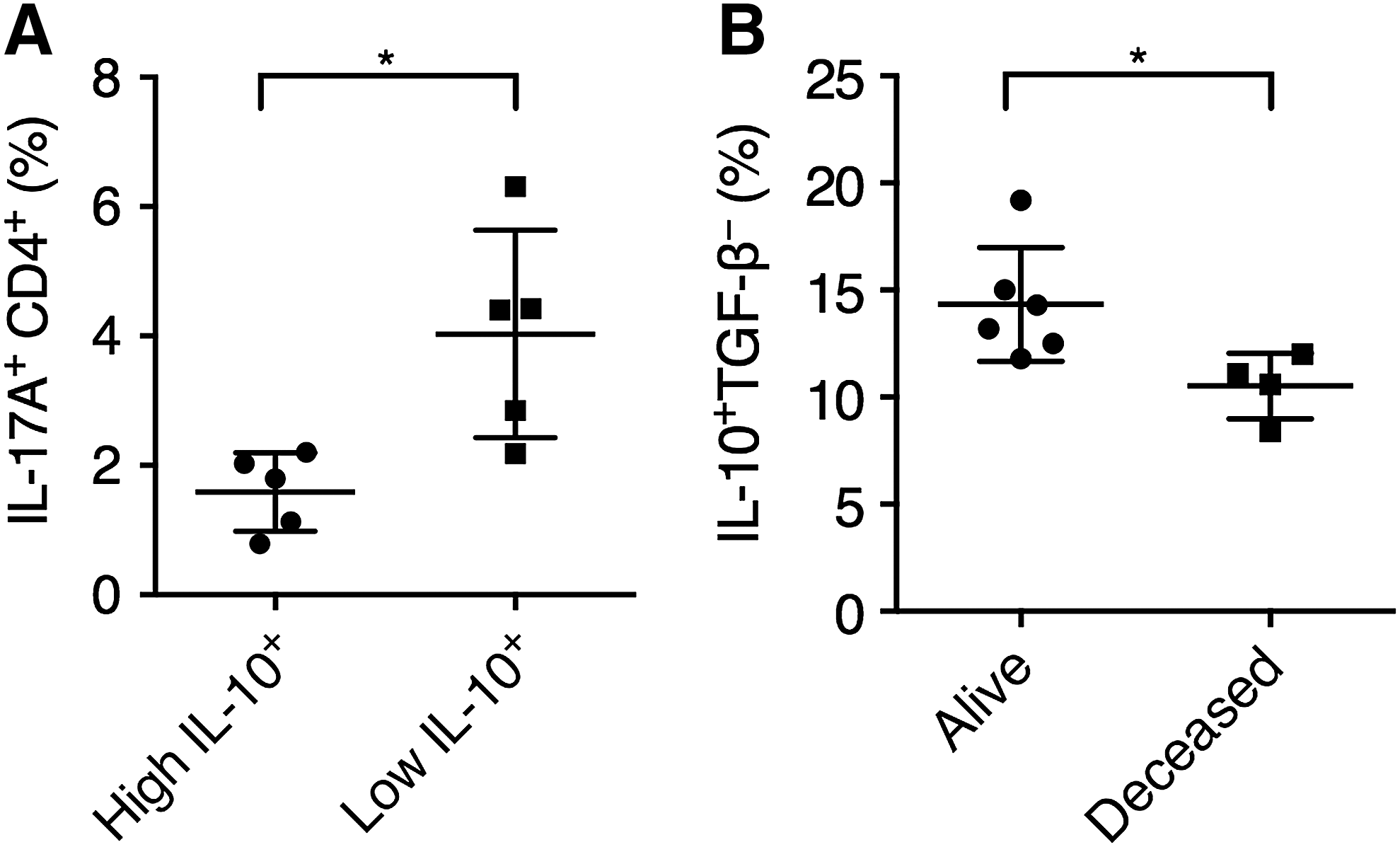
The frequency of tumor-infiltrating IL-10+ plasmablasts was associated with lower IL-17A expression and better survival.
Discussion
The immune system has critical roles in the initiation and progression of cancer. In CRC, the immune system appears to play both tumor-promoting and tumor-inhibitory roles, depending on the cell types (Galon et al., 2006; Terzić et al., 2010). Most cancer studies so far have focused on the activities of CD4+ and CD8+ T cells in mediating antitumor inflammation and tumor cytotoxicity. The enrichment of Breg cells, however, are thought to suppress antitumor inflammation and promote tumor progression, with only a minor fraction of studies suggesting that B cells might support antitumor inflammation through antigen presentation and cytokine secretion (Olkhanud et al., 2011; Nielsen et al., 2012; Schwartz et al., 2016). However, in CRC, chronic inflammation mediated primarily by Th1 and Th17 cells plays a significant role in tumor formation and progression. Therefore, the existence of regulatory B cells, a cell type known to suppress harmful inflammation in autoimmunity, might exert tissue protection in CRC. We previously found that tumors from CRC patients contained a large number of IL-10-producing plasmablasts, the effect of which could not be ignored. Other studies and this have suggested that B cells/plasmablasts possibly exert beneficial roles in CRC (Shimabukuro-Vornhagen et al., 2014; Berntsson et al., 2016). The results presented here demonstrated that IL-10-producing plasmablasts were potent suppressors of Th17 inflammation, a promoter of vascularization and epithelial cell proliferation, and suggested that plasmablasts possibly protected CRC patients through this mechanism.
There are a couple of limitations in this study. First, the Caco-2 cell line was used to represent the CRC cells, which might not fully resemble the primary tumor cells from each individual. Also, to avoid potential main histocompatibility complex (MHC) incompatibility issues, the Caco-2 cells were physically separated from the PBMCs, and thus, the modulatory effects mediated through direct tumor cell to immune cell contact could not be examined in this system. Second, SAC, but not intestinal bacteria from each patient, was used to stimulate B cells. Although this method standardized the experimental condition across different CRC patients, it might not be the most representative bacterium in each patient and discounted the effects of various intestinal commensals. Hence, the results must be carefully interpreted and more investigations are required to examine these issues.
Multiple B cell subsets were found to possess regulatory functions, with potential overlaps and distinctions among various subsets (Rosser and Mauri, 2015). Previously, Breg cells were found to promote Foxp3 expression and converting resting CD4+ T cells into Treg cells (Lee et al., 2014; Wang et al., 2014). In our study, we did not find that the IL-10+ plasmablasts could promote the expression of Foxp3, possibly due to the absence of TGF-β expression by these plasmablasts. Instead, we observed that IL-10 was indispensible for plasmablast-mediated Th17 suppression. The molecular pathways of IL-10 in CRC still require further investigations.
Another question regarding the role of Breg cells and plasmablasts is raised following conclusions of this research. Since the intestinal tract at homeostasis is enriched with plasmablasts and plasma cells, their activity during the CRC initiation stage should be examined in further detail. It is possible that dysregulations in the B cell compartment also contributes to inflammation and CRC development in the intestinal tract. It has been shown that B cells in patients with Crohn's disease, a major inflammatory bowel disease, present elevated T-bet expression and support Th1 inflammation (Wang et al., 2016). The mucosal antibody subtypes in patients with inflammatory bowel diseases also present significant alterations, with a strong upregulation in IgG secretion (Macpherson et al., 1996). Given that inflammatory bowel diseases are common precursors of colitis-associated cancer, it is possible that alterations in the B cell compartment also contribute to CRC induction. So far, very little is known about the role of B cells, plasmablasts, and plasma cells in CRC (Terzić et al., 2010). More research is urgently needed given that these cells make up a significant proportion of lymphocytes in the normal intestinal tract and intestinal tumors.
Footnotes
Disclosure Statement
No competing financial interests exist.