
Abstract
microRNAs (miR) can potentially be used for categorizing the various subtypes of colorectal cancer (CRC) and predicting a patient's response to treatment with traditional anti-CRC therapies. We investigated how miR-1297 and its potential target molecule cyclin D2 (CCND2) might affect the progression of CRC. Thirty-two pairs of CRC specimens and corresponding samples of para-tumor tissue were collected and examined for their levels of miR-1297 and CCND2 expression. We also examined miR-1297 and CCND2 expression in cultured SW480 cells. The effects of modulated levels of miR-1297 and CCND2 on cell viability, anchorage-independent growth ability, proliferation, apoptosis, cell cycle distribution, migration, and invasion were detected using specific techniques. The possible regulatory effect of miR-1297 on CCND2 was investigated using dual luciferase assays. Our results showed that miR-1297 expression was downregulated in clinical CRC specimens, and such downregulation was associated with upregulated levels of CCND2 expression. Upregulation of miR-1297 and downregulation of CCND2 reduced the proliferation and metastasis potential of SW480 cells, but did not affect the apoptotic process. In addition, miR-1297 regulated CCND2 function by directly binding to the promoter sequence of the CCND2 gene, which would block CCND2-related signaling at the transcription level. Our findings validate the anti-CRC function of miR-1297 and pro-CRC function of CCND2. Our findings may assist in developing miR-based therapies against CRC.
Introduction
M
In recent years, some epigenetic studies have suggested that microRNAs (miRs) have the potential to better categorize CRC subtypes and predict the effectiveness of traditional anti-CRC therapies. miRs are single-stranded RNA molecules no longer than ∼22 nucleotides. By binding to the 3′-untranslated region (3′-UTR) of a targeted mRNA molecule, miRs initiate mRNA degradation, suppress RNA translation, and thereby play key roles in numerous biological processes (Bartel, 2004). To date, several miR members have been proven to participate in the onset and progression of CRC. miR-21-5p was reported to be closely associated with lymph node positivity and the distance metastasis of CRC (Slaby et al., 2007). In addition, several studies have suggested that miR-29a-3p expression was significantly higher in samples of primary CRC tissue than in samples of matched adjacent normal tissue (Bandrés et al., 2006; Arndt et al., 2009; Slattery et al., 2011; Fu et al., 2012). Furthermore, the levels of miR-148a-3p, miR-10b-5p, and miR-18a-5p were all proven to be dysregulated in CRC tissue (Dong et al., 2014). In this study, we examined how miR-1297 may contribute to the growth and metastasis of CRC.
miR-1297 has distinct functions in different cancer types. In hepatocellular carcinoma and laryngeal squamous cell carcinoma, miR-1297 can promote tumor progression by targeting HMGA2 and RB1 (Li et al., 2012; Liu et al., 2015, 2016), while in prostate cancer, miR-1297 suppresses the proliferation and invasion of cancer cells by inhibiting the AEG-1-Wnt signaling pathway (Liang et al., 2016). Regarding CRC, miR-1297 was reported to be downregulated in CRC-derived cell lines and clinical CRC specimens, and overexpression of miR-1297 was reported to impair the growth, migration, and invasion of CRC cells by inhibiting cyclo-oxygenase-2 (Chen et al., 2014). The above studies suggest miR-1297 as a potential target for new anti-CRC therapies. Therefore, in this study, we examined whether miR-1297 could be used to identify different CRC subtypes and influence CRC progression.
A bioinformatics analysis showed that miR-1297 has numerous potential targets. In this study, cyclin D2 (CCND2) was selected for study because it is a proven contributor to the onset of multiple cancer types (Takano et al., 2000; Dong et al., 2012; Zhu et al., 2013). The levels of miR-1297 and CCND2 expression were determined in 32 pairs of CRC specimens and matched adjacent tissue specimens. The association between miR-1297 and different clinicopathological categories was analyzed to assess the potential value of using miR-1297 for subtyping CRC patients. Moreover, we also examined how miR-1297 and CCND2 expression are regulated in CRC cells, and the effects of different treatments on CRC cell proliferation and metastasis. We found that miR-1297 was closely related to tumor grade, size, and metastasis in clinical samples, and inhibited CRC progression by directly suppressing CCND2 transcription in CRC cells.
Materials and Methods
Chemicals, agents, and plasmids
Antibodies against CCND2 (ab207604) and GAPDH (ab8245) were purchased from Abcam (Cambridge, MA). Secondary goat anti-rabbit IgG-HRP antibodies were purchased from Beyotime (Shanghai, China). Specific mimics for miR-1297 and negative control (NC) mimics were obtained from Genepharma (Suzhou, China). siRNA (5′-UUAGGUAGCAGCUACUUUAGUCAGC-3′) for CCND2 knockdown and a scrambled version of siRNA (5′-UUACUGGAUGCGACUCAUGAUUAGC-3′) were purchased from Genepharma. Wild-type and mutant sequences of the CCND2 promoter were synthesized by Sangon (Shanghai, China) and ligated to a pcDNA3.1 plasmid. Lipofectamine™ 2000 (cat. no. 52887) was obtained from Invitrogen (Carlsbad, CA). An Annexin V/PI apoptosis kit (cat. no. CCS012) was purchased from MultiSciences (Quebec). Hoechst 33258 staining reagent was purchased from Sigma (St. Louis, MO), a Cell-Light™ EdU Apollo®488/567 In Vitro Imaging Kit (cat. no. C10327) was purchased from RIBOBIO (Guangzhou, China), and RNAiso Plus (cat. no. 9109) was purchased from Takara (Japan). A reverse transcription kit and real-time PCR agents were purchased from DBI (Newark, DE). A Total Protein Extraction Kit was purchased from Beyotime, a Protein Concentration Determination kit (cat. no. 23227) was purchased from Thermo Fisher Scientific (Waltham, MA), and a Dual Luciferase Assay kit (cat. no. E1980) was purchased from Promega (Madison, WI).
Specimen collection
From April 2015 to June 2016, samples of primary CRC tissue were collected from 32 CRC patients at the Fifth Affiliated Hospital, Sun Yat-sen University. Samples of corresponding adjacent carcinoma tissue were also collected from the same patients and used for determining the expression status of miR-1297 and CCND2. All patients involved in the study were provided with detailed information concerning the clinical and pathological aspects of their disease and its prognosis. The study protocol was reviewed and approved by the Fifth Affiliated Hospital, Sun Yat-sen University Ethnics Committee, which also approved the related screening, inspection, and data collection procedures. Each subject signed a written informed consent document stating that all study procedures would be performed in compliance with provisions in the Declaration of Helsinki.
Cell cultures and transfection
SW480 cells were obtained from the American Type Culture Collection (ATCC) (Manassas, VA) and maintained in a medium consisting of 45% F12 medium, 10% FBS, and 1% mixed antibiotics (v/v) (penicillin/streptomycin) at 37°C in an atmosphere of 5% CO2 and 95% air. The 293T cells were provided by the Chinese Academy of Science (Shanghai, China) and cultured in a routine manner. Cells from passages 3 to 5 were used for assays in this study. All transfections done for research purposes were performed using Lipofectamine 2000 according to the manufacturer's instructions.
RNA extraction and reverse transcription quantitative PCR
Total RNA was extracted from cells using RNAiso Plus according to the manufacturer's instructions, after which, cDNA templates were obtained by reverse transcription reactions performed using an RNA reverse transcription kit according to the manufacturer's instructions. The final 20 μL reaction mixture contained 10 μL of quantitative polymerase chain reaction (qPCR) mix, 0.5 μL of each primer [miR-1297 forward: 5′-ACACTCCAGCTGGGTTCAAGTAATTC-3′, reverse: 5′-GTGCAGGGTCCGAGGT-3′; CCND2 forward: 5′-CTACCGACTTCAAGTTTGCC-3′, reverse: 5′-GCTTTGAGACAATCCACATCAG-3′; GAPDH (internal reference) forward: 5′-TGTTCGTCATGGGTGTGAA-3′, reverse: 5′-ATGGCATGGACTGTGGTCAT-3′; and U6 (internal reference) forward: 5′-CTCGCTTCGGCAGCACA-3′, reverse: 5′-AACGCTTCACGAATTTGCGT-3′], 1 μL of the cDNA template, and 8 μL of RNase-free H2O. The thermal cycling parameters used for amplification were as follows: denaturation at 94°C for 2 min, followed by 40 cycles at 94°C for 20 s, 58°C for 20 s, and 72°C for 30 s. The reaction was stopped by incubation at 25°C for 5 min. A melting curve was constructed and analyzed between 62°C and 95°C. The relative expression levels of the target genes were calculated by a Real-time PCR Detection System (Mx3000P; Agilent, Santa Clara, CA) and using the formula, 2−ΔΔCt.
Western blotting assay
Total protein was extracted using the Total Protein Extraction Kit according to the manufacturer's instructions. The protein concentration in each sample was determined using a Protein Concentration Determination kit according to the manufacturer's instructions. Western blot assays were performed using standard procedures. Briefly, the cell membranes were incubated with one of the primary antibodies against CCND2 (1:1000) and GAPDH (internal reference) (1:2000) according to the respective conditions, after which, they were incubated with peroxidase-conjugated secondary antibodies (1:10,000). The blotting images were analyzed with a Gel Imaging System (Life Science), and the relative expression levels of different proteins were calculated using Bio-Rad Quantity One software version 4.6.3 (Bio-Rad, Hercules, CA).
Dual luciferase assay
To determine how miR-1297 regulates CCND2 expression, the miRNA target predication databases TargetScan (
Cell viability and proliferation assays
Aliquots of exponentially growing SW480 cells (5 × 105 cells/mL) were seeded into the wells of a 96-well plate. The cells were then incubated for 72 h, after which, their viability was detected by adding 10 μL/mL of Cell Counting Kit-8 (CCK-8) solution to selected wells and then incubating the plate at 37°C for 60 min. OD values at 450 nm were detected with a Microplate Reader (GloMax; Promega) and used as indicators of cell viability. The proliferation of SW480 cells was assessed after 48 h of culture using an EdU assay according to the manufacturer's instructions, and the results were observed under a fluorescence microscope (Olympus, Japan).
Colony formation assay
The capability SW480 cells for anchorage-independent growth was measured using the colony formation assay. In brief, the SW480 cells were suspended in a medium containing 10% serum and 0.35% agarose, and then seeded into 35 mm plates at a density of 200 cells per plate. After 1 week of culture, the cell colonies were stained with Wright-Giemsa stain, and the number of colonies in each plate was recorded. The colony formation rate equaled the ratio of the colony number to the inoculated cell number on each plate.
Hoechst 33258 staining
The deformation of cell nuclei due to apoptosis was detected using a Hoechst staining kit according to the manufacturer's instructions. The results were observed under a fluorescence microscope at 460 nm.
Flow cytometry
The cell cycle distributions of SW480 cells given different treatments were determined using flow cytometry. Briefly, 300 μL of propidium iodide (PI)-FITC was added to different wells to stain the DNA of cells in the dark. After 20 min of incubation at room temperature, the DNA content of the cells was analyzed using a FACS flow cytometer (Accuri C6; BD, Franklin Lakes, NJ).
The apoptotic process in SW480 cells was detected using an Annexin V-FITC apoptosis detection kit according to the manufacturer's instructions. The apoptotic rates were analyzed using a FACScan flow cytometer (Accuri C6; BD). The apoptotic cell rate (UR + LR—the total apoptotic cell percentage) was equal to the sum of the late apoptotic rate (UR, upper right quadrant-advanced stage apoptosis cell percentage) and the early apoptotic rate (LR, lower right quadrant-prophase apoptosis cell percentage).
Transwell experiments
Transwell experiments were performed to evaluate the invasion and migration capabilities of CRC cells in the different treatment groups. Briefly, a 200 μL aliquot of incubation medium (1 × 104 cells) was seeded into the upper compartment of each transwell chamber (Corning Star, Cambridge, MA) and incubated for 24 h at 37°C to allow for cell migration through the porous membrane. Following this incubation, cells remaining on the upper surface of the chamber were completely removed, while the lower surfaces of the membranes were fixed with 4% paraformaldehyde for 20 min, and then stained in 0.5% (w/v) crystal violet solution for 5 min. After being washed in ddH2O, the numbers of cells in the different groups were determined using Image-Pro Plus 6.0 software (Nikon, Japan). Next, the invasion capability of the CRC cells was measured as already described. These assays were performed using polycarbonate membranes previously coated with 40 μL of Matrigel (1.5 mg/mL; BD Biosciences, San Jose, CA) at 37°C for 2 h to form a reconstituted basement membrane.
Statistical analysis
All the statistical analyses were performed using IBM SPSS Statistics for Windows, version 19.0 (IBM, Armonk, NY).
All data are expressed as the mean ± standard deviation. The relationship between miR-1297 and CCND2 expression at the mRNA level was analyzed by Pearson's correlation analysis. Differences between two groups were analyzed using the t-test, and two-tailed p-values ≤0.05 were considered statistically significant.
Results
The level of miR-1297 expression was correlated with the CRC subtype
The levels of miR-1297 expression were investigated in clinical CRC specimens. When compared with samples of adjacent carcinoma tissue, the levels of miR-1297 expression were significantly downregulated in tumor tissues (p < 0.05) (Fig. 1). Moreover, we found that the levels of miR-1297 were dramatically different in tumors with different subtypes. The relative level of miR-1297 expression was significantly higher in Grade I/II tumors, tumors with a size >5 cm, distant metastasis, and in tumors with higher degrees of differentiation than in Grade III/IV tumors, tumors sized <5 cm, nondistant metastasis, and poorly differentiated tumors (Table 1). These results suggested that miR-1297 could be used for classifying CRC subtypes in clinical practice.
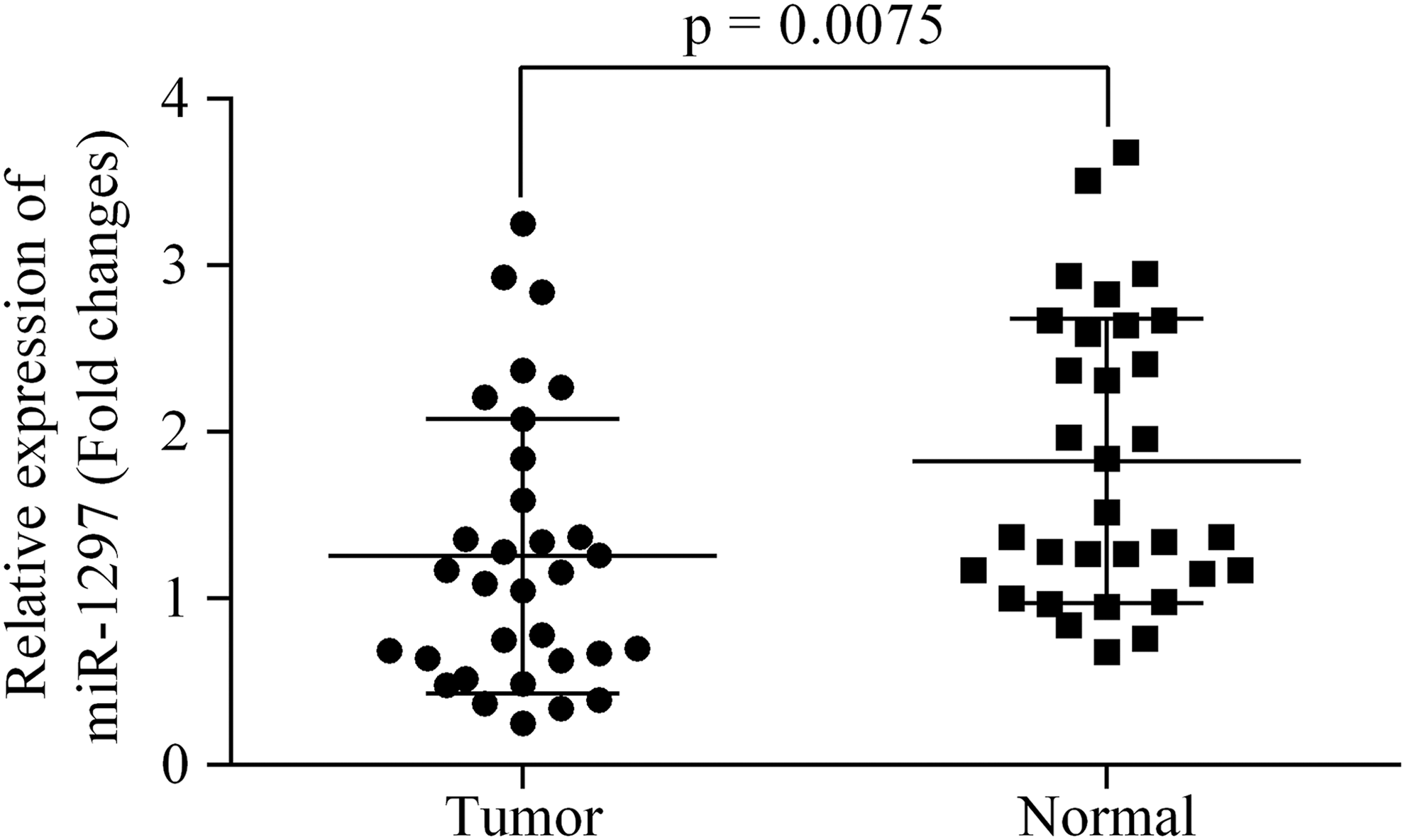
The expression of miR-1297 was downregulated in CRC specimen than that in normal tissues. CRC, colorectal cancer.
The relative expression of miR-1297 was calculated using 2−ΔΔCq method.
Bold values represent p < 0.05 with a significant level.
Induced expression of miR-1297 suppressed the growth of SW480 cells
Based on the information derived from our clinical investigation, we specifically induced miR-1297 expression in human SW480 CRC cells (Fig. 2A) to detect the effect of miR-1297 overexpression on the growth ability of those cells. After transfection of miR-1297 mimics, the viability of SW480 cells, as detected by the CCK-8 assay, was significantly decreased at the last two recorded time points (p < 0.05) (Fig. 2B). The anchorage-independent growth ability of tumor cells is an indicator of their tumorigenesis potential. In this study, fewer cells were detected in the colony formation assay after transfection of the miR-1297 mimics (Fig. 2C, D). SW480 cell proliferation was also inhibited by the miR-1297 mimics (Fig. 2E), as fewer proliferating cells (stained red) were recorded in the miR-1297 mimics group. The above results suggest that miR-1297 overexpression suppresses CRC cell proliferation.
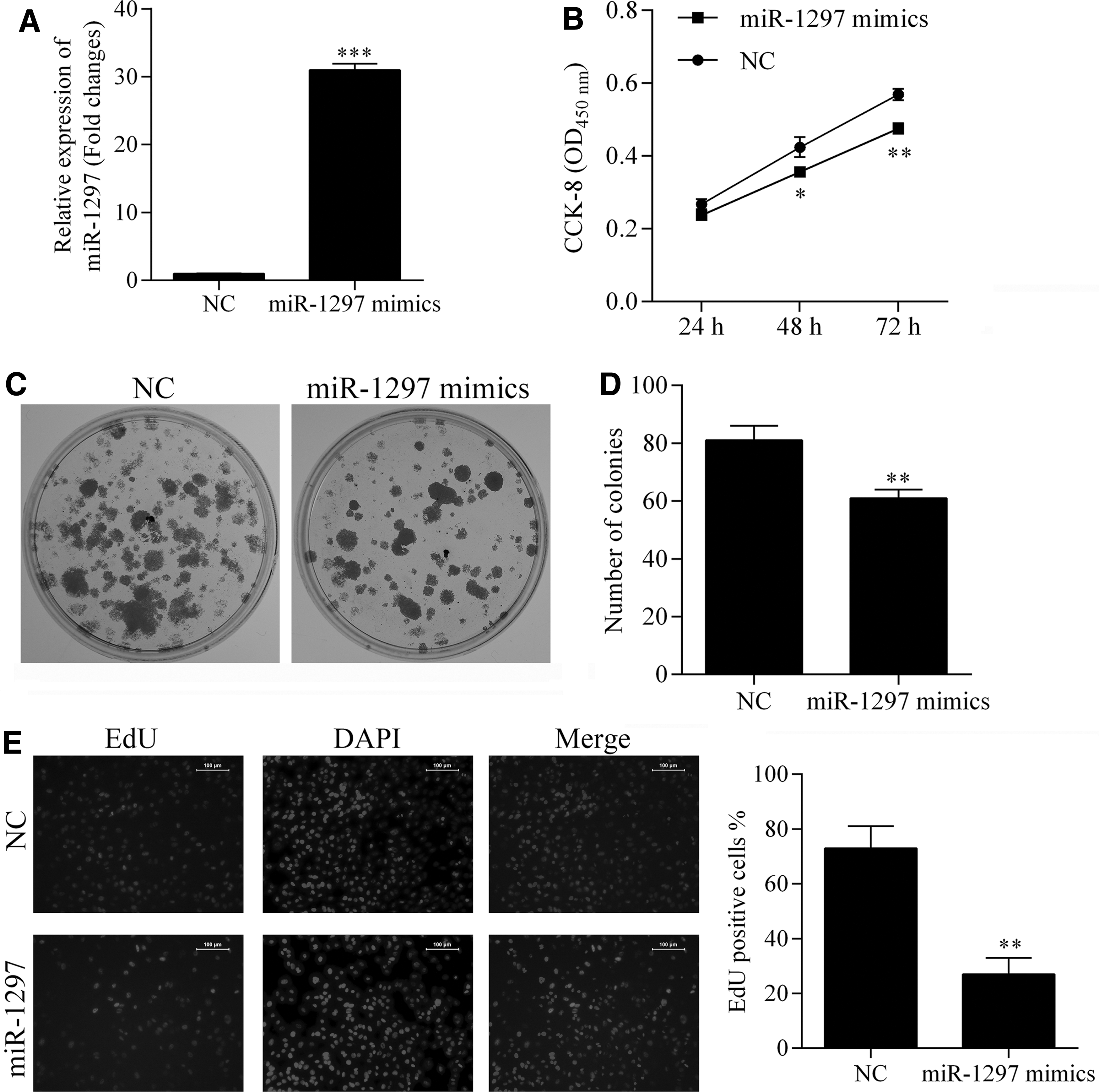
Detection of the effect of miR-1297 overexpression on cell viability, anchorage-independent growth, and proliferation ability of SW480 cells. The expression of miR-1297 was induced by transfection of miR-1297-specific mimics
Induced expression of miR-1297 initiated G1 cell cycle arrest and inhibited invasion/migration of SW480 cells
We next investigated the effect of miR-1292 overexpression on the cellular apoptosis process and cell cycle distribution. As shown in Figure 3A and B, transfection of miR-1297 had no effect on the apoptotic process in SW480 cells. However, upregulated levels of miR-1297 induced G1 cell cycle arrest, which subsequently inhibited the proliferation of SW480cells. Because CRC is a highly aggressive type of cancer, we also investigated the role played by miR-1297 in CRC metastasis. After transfection of miR-1297 mimics, the invasion and migration capabilities of SW480 cells were both impaired, as fewer cells penetrated membranes in the Transwell assay (Fig. 3D).
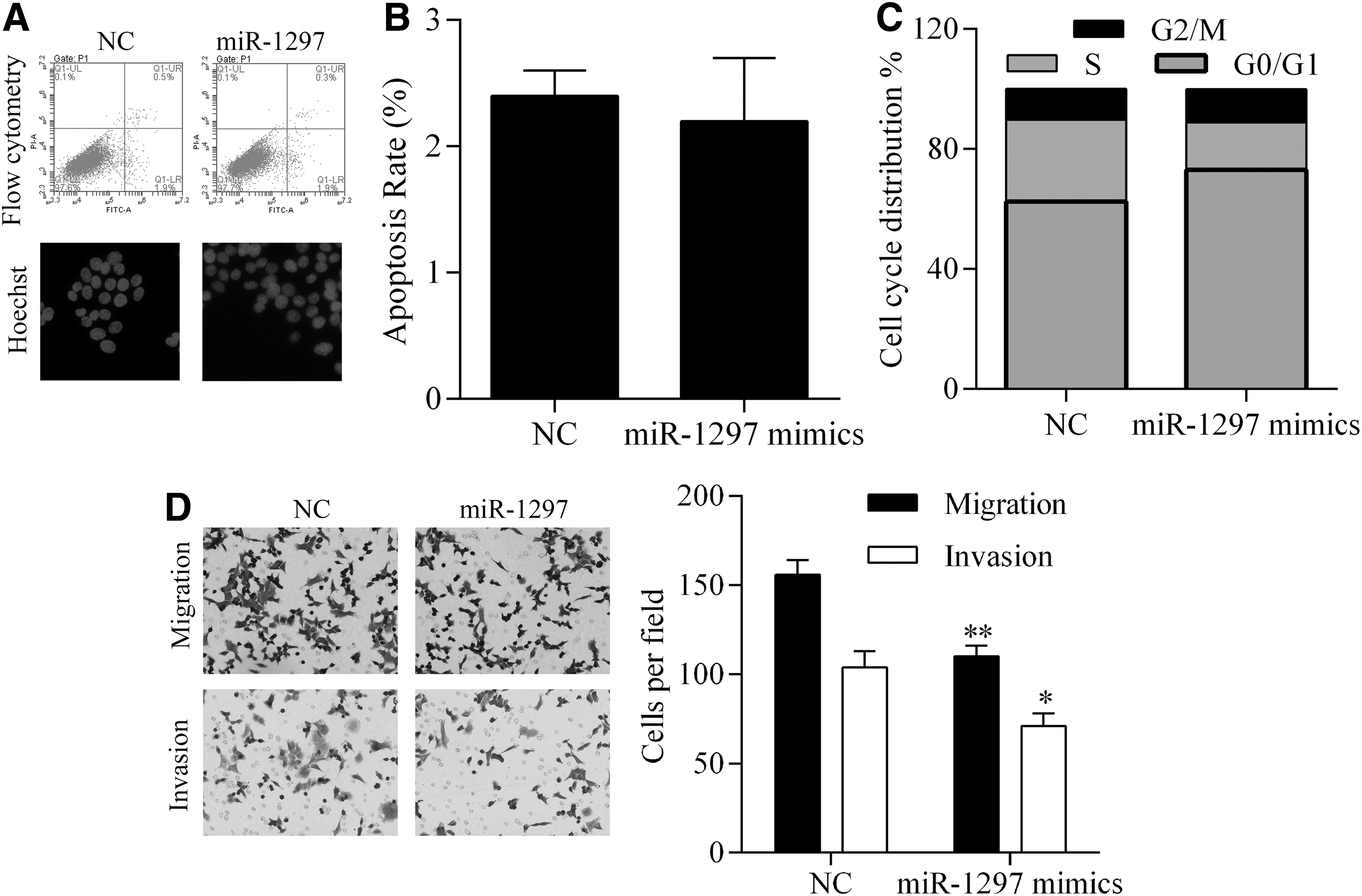
Detection of the effect of miR-1297 overexpression on apoptosis, cell cycle distribution, invasion, and migration of SW480 cells. Induced expression of miR-1297 had no effect on cell apoptosis rates
CCND2 expression was upregulated in CRC specimens, and miR-1297 interacted with CCND2 by directly binding to the CCND2 gene promoter region
Our bioinformatics analysis identified CCND2 as a potential target of miR-1297. CCND2 contributes to the progression of multiple cancer types. We examined the levels of CCND2 expression in CRC specimens at both the mRNA and protein levels and found that an onset of CRC increased the levels of CCND2 expression (Fig. 4A, B). Moreover, our Pearson's correlation analysis identified a negative relationship between miR-1297 mRNA levels and CCND2 (Fig. 4C). Furthermore, our in vitro assay showed that transfection of miR-1297 mimics suppressed the expression of CCND2 (Fig. 4D, E). To explicitly elucidate the interaction between miR-1297 and CCND2, vectors carrying wild-type and mutant-type CCND2 promoters were cotransfected with miR-1297 mimics into 293T cells. Data from the subsequent luciferase assays indicated that cotransfection of miR-1297 and the wild-type CCND2 promoter significantly inhibited the luciferase activity by direct binding of miR-1297 to the CCND2 promoter (Fig. 4F). In addition, cotransfection of miR-1297 and the mutant-type CCND2 promoter had no influence on the luciferase activity, which indicated that the binding of miR-1297 to the CCND2 gene promoter region was specific (Fig. 4F).
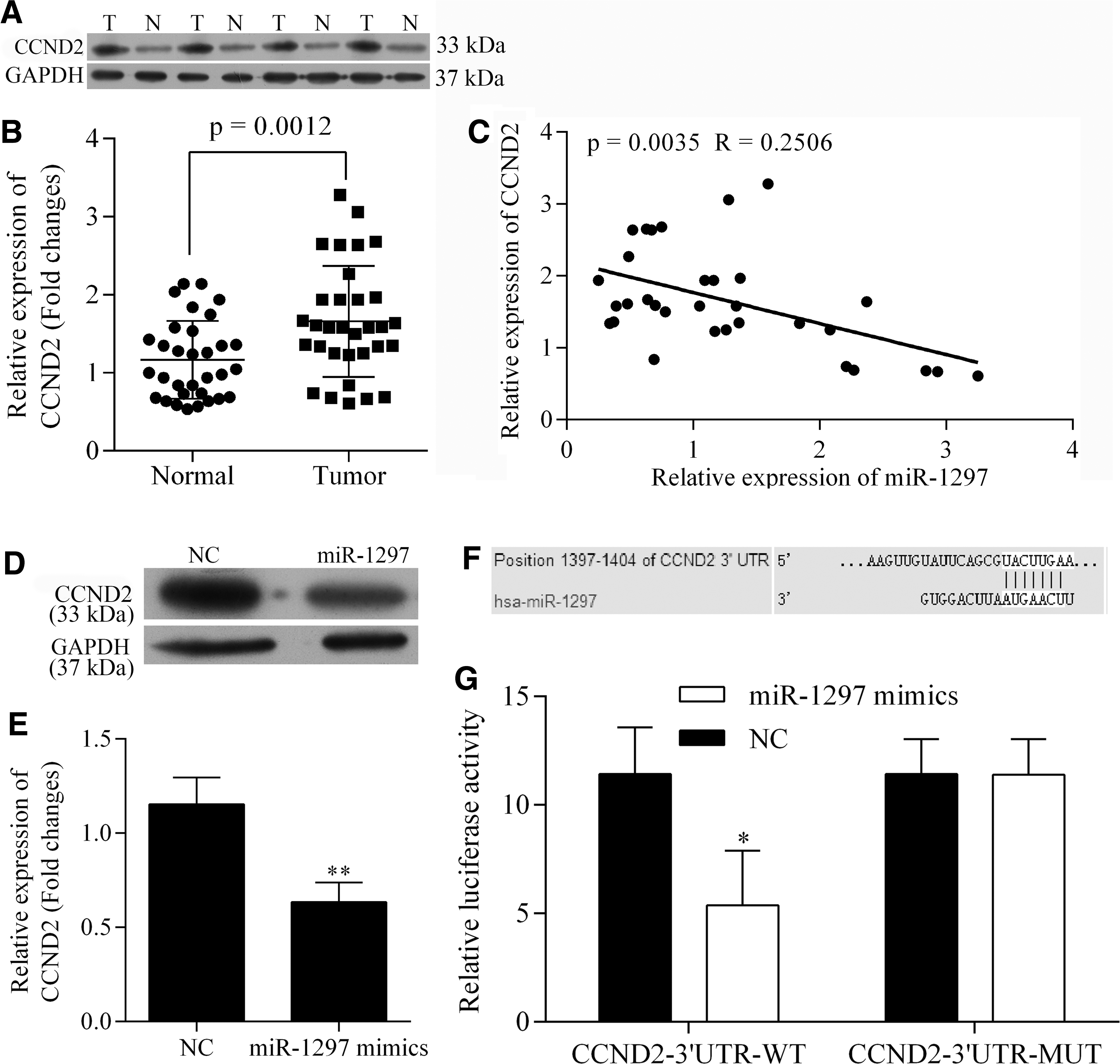
Detection of expression status of CCND2 in CRC specimen and regulating mechanism of miR-1297 on CCND2. The expression of CCND2 was upregulated in CRC specimen both at mRNA and protein levels
Knockdown of CCND2 inhibited CRC by suppressing cell proliferation and metastasis potential
CCND2 demonstrates a protumor function in various types of cancers. In this study, we knocked down the levels of CCND2 in SW480 cells to assess how CCND2 affects the oncogenesis and progression of CRC (Fig. 5A, B). The effect of CCND2 knockdown was similar to that of miR-1297 overexpression, as suppression of CCND2 was associated with decreased levels of cell viability, anchorage-independent growth ability, and proliferation (Fig. 5). In addition, knockdown of CCND2 had no impact on cell apoptosis, but resulted in G1 cell cycle arrest and impaired invasion and migration capabilities of SW480 cells (Fig. 6), which was also identical with the effect of miR-1297 overexpression.
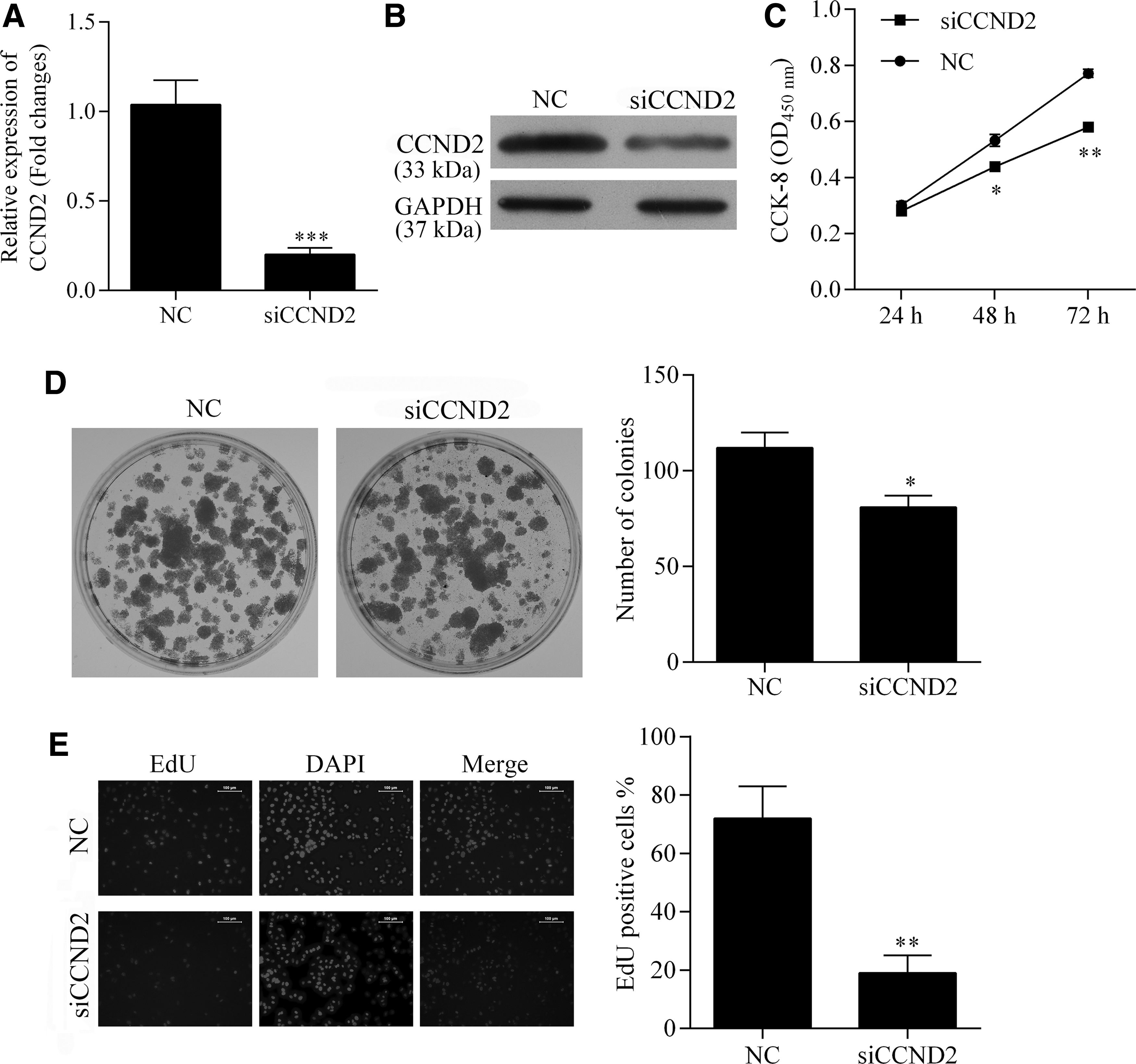
Detection of the effect CCND2 knockdown on cell viability, anchorage-independent growth, and proliferation ability of SW480 cells. The expression of CCND2 was knocked down by transfection of CCND2-specific siRNA
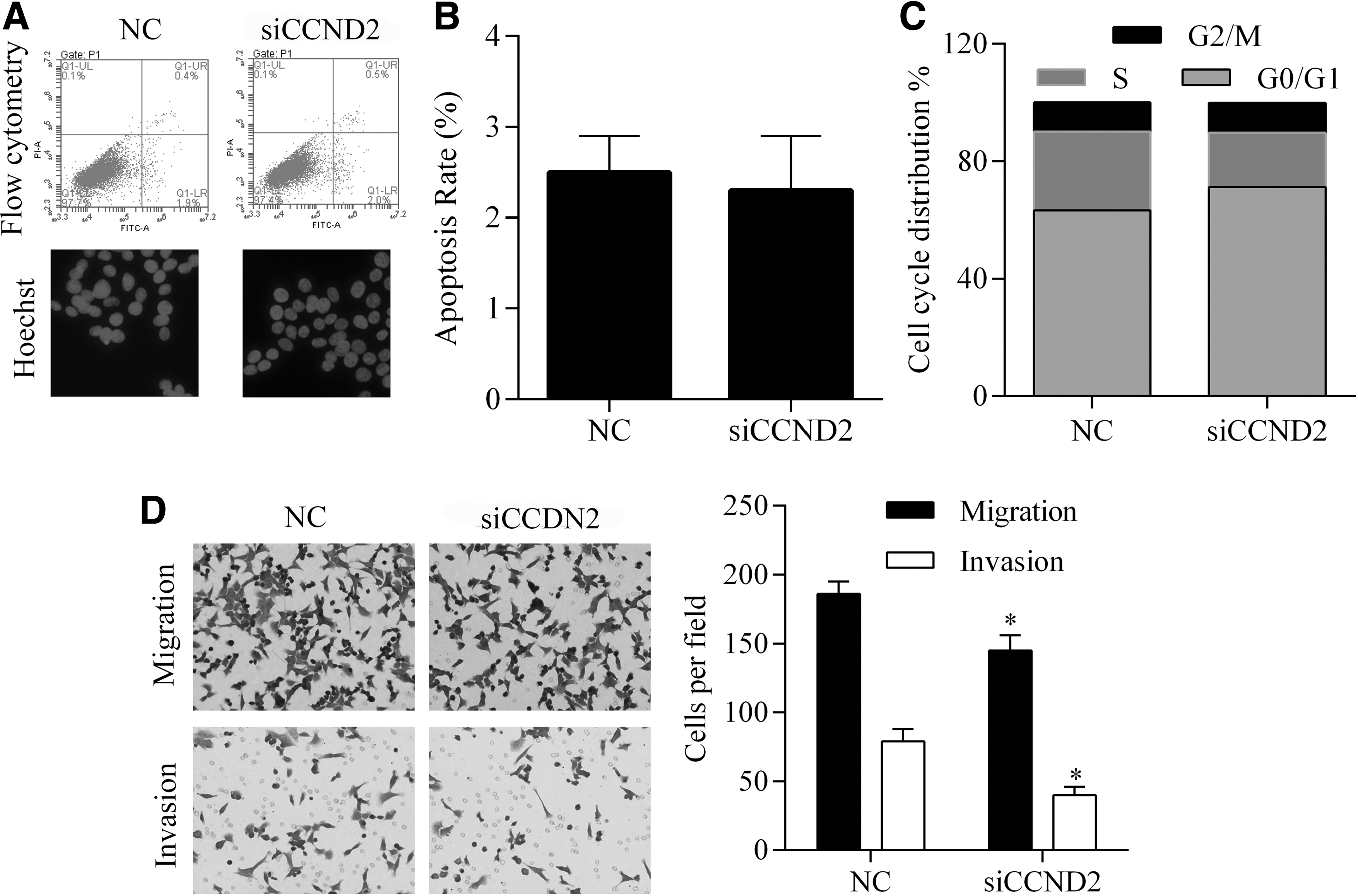
Detection of the effect of CCND2 knockdown on apoptosis, cell cycle distribution, invasion, and migration of SW480 cells. Knockdown of CCND2 had no effect on cell apoptosis rates
Discussion
In this study, miR-1297 expression was proven to be downregulated in clinical samples of CRC tissue, and downregulation was associated with an upregulation of CCND2. By using an in vitro research model, we found that miR-1297 overexpression and suppression of CCND2 both exerted an anti-CRC function, which manifested as decreased rates of CRC cell proliferation, migration, and invasion, and the induction of G1 cell cycle arrest. Furthermore, the direct binding of miR-1297 to the promoter sequence of the CCND2 gene was verified with a dual luciferase assay. These findings affirmed the potential of miR-1297 as an anti-CRC target for development of novel prevention and treatment modalities that exert their effects by suppression of CCND2.
CRC is characterized by its slow development from premalignant lesions, which makes it even more important to detect and treat the malignancy in its phase to reduce the burden of the disease. In recent years, with the development of molecular and bioinformatics techniques, gene expression studies of tumor samples have revealed the dysregulation of numerous noncoding genomic molecules in CRC (Luo et al., 2011), including miRs. MiR members such miR-21-5p (Slaby et al., 2007), miR-29a-3p (Bandrés et al., 2006; Arndt et al., 2009; Slattery et al., 2011; Fu et al., 2012), miR-148a-3p, miR-10b-5p, and miR-18a-5p were previously reported to be involved in the oncogenesis and progression of CRC (Dong et al., 2014). A previous study revealed that mrR-1297 participates in the proliferation, migration, and invasion of CRC cells (Chen et al., 2014), and those findings were further validated in this study. SW480 cells transfected with miR-1297 mimics displayed decreased rates of viability, anchorage-independent growth, and proliferation. In addition, upregulated expression of miR-1297 also induced G1 cell cycle arrest in SW480 cells. Although it was confirmed that miR-1297 mimics had no effect on the apoptosis of CRC cells, these findings evidently represented the suppressing effect of the miRNA on CRC. CRC is also characterized by a high metastasis potential (Siegel et al., 2012), inhibition of which contributes to treatment of the cancer (Isaac and Kochubiei, 2014). The migration and invasion of cancer cells are important factors affecting cancer metastasis; therefore, we investigated how miR-1297 overexpression affects the migration and invasion capabilities of CRC cells. Transfection of miR-1297 inhibited the migration and invasion of SW480 cells when compared with cells in a control group, and this finding was consistent with results reported by Chen et al. (2014).
To further understand the mechanism that drives the function of miR-1297 in CRC, we conducted a bioinformatics prediction search on the TargetScan data base (
In conclusion, the findings outlined in this study validate the anti-CRC function of miR-1297 and pro-CRC function of CCND2 by using data obtained from a clinical investigation and laboratory experiments. MiR-1297 exerted its suppressing effect on CRC cells by directly targeting the CCND2 gene, and this finding can be used to assist in developing new miR-based therapies for CRC.
Footnotes
Acknowledgments
This work was supported by the Fifth Affiliated Hospital of Sun Yat-sen University.
Disclosure Statement
No competing financial interests exist.