
Abstract
Exosomes are small vesicles secreted by a variety of cell types under physiological and pathological conditions. When Saccharomyces cerevisiae are grown in low glucose, small vesicles carrying more than 300 proteins with diverse biological functions are secreted. Upon glucose addition, secreted vesicles are endocytosed that requires cup-shaped organelles containing the major eisosome protein Pil1p at the rims. We aim to identify genes that regulate the function of cup-shaped organelles in vesicle endocytosis. In cells lacking either VID27 or VID21, Pil1p distribution was altered and cup-shaped organelles became elongated with narrower openings. Change in shape reduced the number of vesicles in the deeper areas and impaired vesicle endocytosis. Vid21p and Vid27p were localized to vesicle clusters and interacted with other Vid proteins. In the absence of these genes, these vesicles failed to aggregate and were secreted. Vid21p and Vid27p are required for the aggregation and retention of vesicles that contain Vid proteins in the cytoplasm. Increased vesicles near the plasma membrane in mutant strains correlate with an increased Pil1p movement resulting in the fusion of cup-shaped organelles. We conclude that the shape of vesicle-containing organelles is critical for their functions in vesicle endocytosis.
Introduction
E
Saccharomyces cerevisiae serve as an important model system in elucidating the effects of environmental changes on secretion and endocytosis of small vesicles. For example, small vesicles/exosomes (30–50 nm in diameter) are secreted into the periplasm when yeast cells are grown in low glucose for prolonged periods of time. The extracellular vesicles are then endocytosed upon glucose addition to glucose-starved cells. Various yeast gluconeogenic enzymes such as fructose-1,6-bisphosphatase (FBPase), phosphoenolpyruvate carboxykinase, isocitrate lyase, and malate dehydrogenase have been used as markers to study the secretion, endocytosis, and trafficking of vesicles to vacuoles for degradation through the Vid (vacuole import and degradation) pathway (Hoffman and Chiang, 1996; Huang and Chiang, 1997; Brown et al., 2000, 2001, 2002, 2008, 2010b; Shieh et al., 2001; Brown et al., 2003, 2010a; Hung et al., 2004). In addition to these gluconeogenic enzymes, vesicles carrying more than 300 proteins with diverse biological functions are also secreted during glucose starvation (Giardina et al., 2014a). Upon glucose addition, these extracellular vesicles are endocytosed into the cytoplasm. Different mechanisms, including phagocytosis, clathrin/dynamin-1-mediated endocytosis, and micropinocytosis have been shown to mediate the endocytosis of extracellular vesicles (French et al., 2017; Maas et al., 2017; Valdinocci et al., 2017). However, a direct transport of vesicles across the plasma membrane has not been reported.
Yeast eisosomes are immobile organelles on the plasma membrane that may have important roles in the endocytosis of proteins and small molecules (Zhang et al., 2004; Walther et al., 2006, 2007; Deng et al., 2009; Murphy and Kim, 2012; Moreira et al., 2012). Eisosomes are localized to plasma membrane furrows/invaginations (300 nm long and 50 nm deep) and contain a number of proteins, including Pil1p (Stradalova et al., 2009; E and K, 2012; Douglas and Konopka, 2014). Recently, we purified cup-shaped structures (100–300 nm in diameter) required for the endocytosis of extracellular vesicles. These structures contained Pil1p at the rims and multiple small vesicles in the deeper areas/openings. Deletion or mutation of PIL1 impaired the glucose-induced endocytosis of extracellular vesicles (Stein et al., 2017). Based on these results, we hypothesized that extracellular vesicles are internalized from the periplasm to the cytoplasm by vesicle-conducting channels, and that cup-shaped organelles are the extracellular domains of these channels. The aim of this study was to identify VID genes that regulate the function of these organelles in vesicle endocytosis. Our results indicate that VID27 and VID21 play important roles in maintaining the shape of these vesicle-containing organelles. In cells lacking either VID27 or VID21, the distribution of Pil1p on the plasma membrane was altered and cup-shaped organelles were elongated with narrower openings. Furthermore, vesicles carrying other Vid proteins failed to aggregate and were secreted. Increased vesicles near the plasma membrane may alter the microenvironment surrounding Pil1p/eisosomes. This led to increased Pil1p movement and fusion of Pil1p-containing organelles to form elongated structures that inhibited vesicle endocytosis.
Materials and Methods
Reagents
The yeast strains and the primers used in the study are listed in Supplementary Tables S1 and S2 (Supplementary Data are available online at
Cell culture
Cells were cultured in low-glucose YPKG media (1% yeast extracts, 2% peptone, 1% potassium acetate, and 0.5% glucose) for 3 days unless otherwise indicated. For FBPase degradation and vesicle endocytosis experiments, cells were transferred to high-glucose YPD media containing 1% yeast extracts, 2% peptone, and 2% glucose and collected at the indicated time points.
Differential centrifugation
Cytosol- and vesicle-enriched proteins were obtained using previously described techniques (Brown et al., 2000, 2003). Briefly, cells were collected in FBPase buffer (50 mM HEPES, pH 7.2, 5 mM MgSO4, 40 mM (NH4)2SO4, and 0.1 mM EDTA) and total lysates were subjected to centrifugation at 13,000 g for 10 min. Supernatants were centrifuged at 100,000 g for 2 h. The resulting supernatants were then centrifuged for 2 h at 200,000 g. The resultant 200,000 g pellets (P200) contained free vesicles, whereas soluble proteins were enriched in the 200,000 g supernatant (S200) fraction. The S200 and P200 fractions were resolved by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and examined for distribution by western blotting. Vid24p-HA and Vid30-HA were detected using anti-HA antibodies, respectively.
Extraction of periplasmic proteins
Cells were grown in YPKG for 3 days and equal amounts of cells (OD600 = 10/mL) were transferred to medium containing high glucose for 0, 15, and 30 min. At each time point, NaN3 was added to the samples. Cells were then pelleted and extracted with 200 μL of extraction buffer (0.1 M Tris pH 9.4 and 10 mM β-mercaptoethanol) in a 37°C shaker for 15 min (Giardina et al., 2014a). Following incubation, cells were centrifuged at 3000 g for 10 min. Proteins from the supernatant were precipitated using trichloroacetic acid to obtain the extracellular fraction (E) proteins. Cell pellets containing intracellular (I) proteins were lysed in 200 μL of FBPase buffer. Protein from both fractions of lysates were denatured in 2× Laemmli sample buffer and analyzed by western blotting.
Pulldown experiments and immunoblotting
His beads pulldown experiments using Ni-NTA agarose were performed according to the manufacturer's suggestions (Qiagen). For immunoblotting experiments, equal amounts of cells (OD600 = 10/mL) were used to prepare proteins for electrophoresis on polyacrylamide gels and transfer to nitrocellulose membranes. The resulting blots were blocked with 5% nonfat dry milk in Tris-buffered saline (TBS) and incubated with antibodies of interest. Unbound primary antibodies were removed by washing with TBS containing 0.05% Tween-20 (ICI Americas, Inc., Wilmington, DE). Blots were incubated with an enhanced chemiluminescent (ECL) detection system (GE Healthcare, Buckinghamshire, United Kingdom) and exposed to X-ray blue film (Cole Parmer, Vernon Hills, IL) and quantified with ImageJ (NIH, Bethesda, MD).
Fluorescence microscopy for actin staining
Yeast cells expressing Vid27p-GFP or Vid21p-GFP were grown in YPKG for 3 days. Cells were prepared as previously described (Brown et al., 2008; Alibhoy et al., 2012; Giardina et al., 2013), and then incubated for 30 min in the dark at room temperature with 1 μL of rhodamine-conjugated phalloidin (0.2 U/μL in methanol). Fluorescence signals were visualized at 26°C using the FLUAR 100 × objective lens with 1.30 NA as well as the GFP and rhodamine filters. Images were taken using a Zeiss Axiovert S100 inverted microscope with Zeiss Zen Blue Software and an Axiocam 503 Mono camera.
Transmission electron microscopy
Cells were grown in 100 mL of YPKG for 3 days and extracellular organelles were obtained after extraction in extraction buffer containing 0.1 M Tris pH 9.4 and 10 mM β-mercaptoethanol (Giardina et al., 2014a, 2014b). Total extracts were centrifuged sequentially at 3000 g for 10 min, 13,000 g for 20 min, and then additional 2 h at 200,000 g. The resulting 13,000 g pellets contained the cup-shaped organelles, whereas the 200,000 g pellets contained extracellular vesicles. These pellet fractions were resuspended in 20 μL of phosphate-buffered saline (PBS), followed by fixation in 3% paraformaldehyde with 0.2% glutaraldehyde for 2 h or overnight at 4°C. The fixed pellets were incubated for 30 min on nickel grids. The grids were washed once with PBS buffer and twice with distilled water. The grids were dried using Whatman filter paper and stained with 2% uranyl acetate for 5 min. The grids were dried and viewed on a JEOL JEM-1400 electron microscope.
Plasmid preparations
To generate plasmids containing Vid27p-tagged V5-His6, the VID27 gene was amplified by polymerase chain reaction (PCR) reactions using MCCA13 and MCCA14 as the forward and reverse primers (Supplementary Table S2). PCR products were purified and cloned into a TOPO plasmid (Invitrogen). Vid27p-V5-His6 was digested with NruI and SwaI to remove the 2 μ sequence and religated. The resulting plasmid was linearized with EcoRI and integrated into the VID27 locus. Following the integration, Vid27p-V5-His6 was expressed under its own promoter. The second copy of untagged Vid27p-V5-His6 was not expressed because of the absence of galactose in the medium. Cells expressing Vid27p-V5-His were examined for the expression of Vid27p-V5-His6 and FBPase degradation. The VID21 gene was amplified by PCR using the F257 forward and R258 reverse primers. PCR products were cloned into a TOPO plasmid. The 2 μ sequence was removed by NruI and SwaI digestion and religated. The plasmid was digested with XhoI and integrated into the VID21 locus. The expression of Vid21p-V5-His was examined using western blotting with anti-V5 antibodies. Vid21p-GFP and Vid27p-GFP was produced by PCR reactions using the primers listed in Supplementary Table S2.
Statistical analysis
An unpaired Student's t-test was performed. Data are presented as mean ± standard error of mean of at least three repeated individual experiments. A value of p < 0.05 was considered to be statistically significant.
Results
VID27 regulates Pil1p distribution and the morphology of cup-shaped organelles
Recent evidence suggests that cup-shaped structures are required for endocytosis of extracellular vesicles targeted for degradation through the Vid trafficking pathway (Stein et al., 2017). In our characterization of the cup-shaped structures, we identified the presence of the major eisosome protein Pil1p on the rims of these organelles. Cup-shaped organelles were absent when PIL1 was mutated or deleted (Stein et al., 2017). Mutations of Pil1p have been shown to alter the distribution of Pil1p on the plasma membrane (Luo et al., 2008; Deng et al., 2009). Disruption of the Vid pathway may affect Pil1p localization on the plasma membrane. To identify VID genes that regulated the distribution of Pil1p on the plasma membrane, vid mutant cells expressing Pil1p-GFP were grown in low glucose for 3 days and screened for changes in Pil1p distribution. As illustrated in Figure 1A, Pil1p was observed in many small puncta on the plasma membrane in wild-type cells examined using fluorescence microscopy. However, in Δvid27 mutant cells, Pil1p-GFP localized to fewer but larger puncta (Fig. 1B). Interestingly, levels of Pil1p in total lysates were similar in wild-type and Δvid27 cells, indicating that changes in Pil1p distribution in the Δvid27 mutant are not due to altered amounts of Pil1p in mutant strains (Fig. 1C). Therefore, VID27 is critical in maintaining the distribution of Pil1p on the plasma membrane.
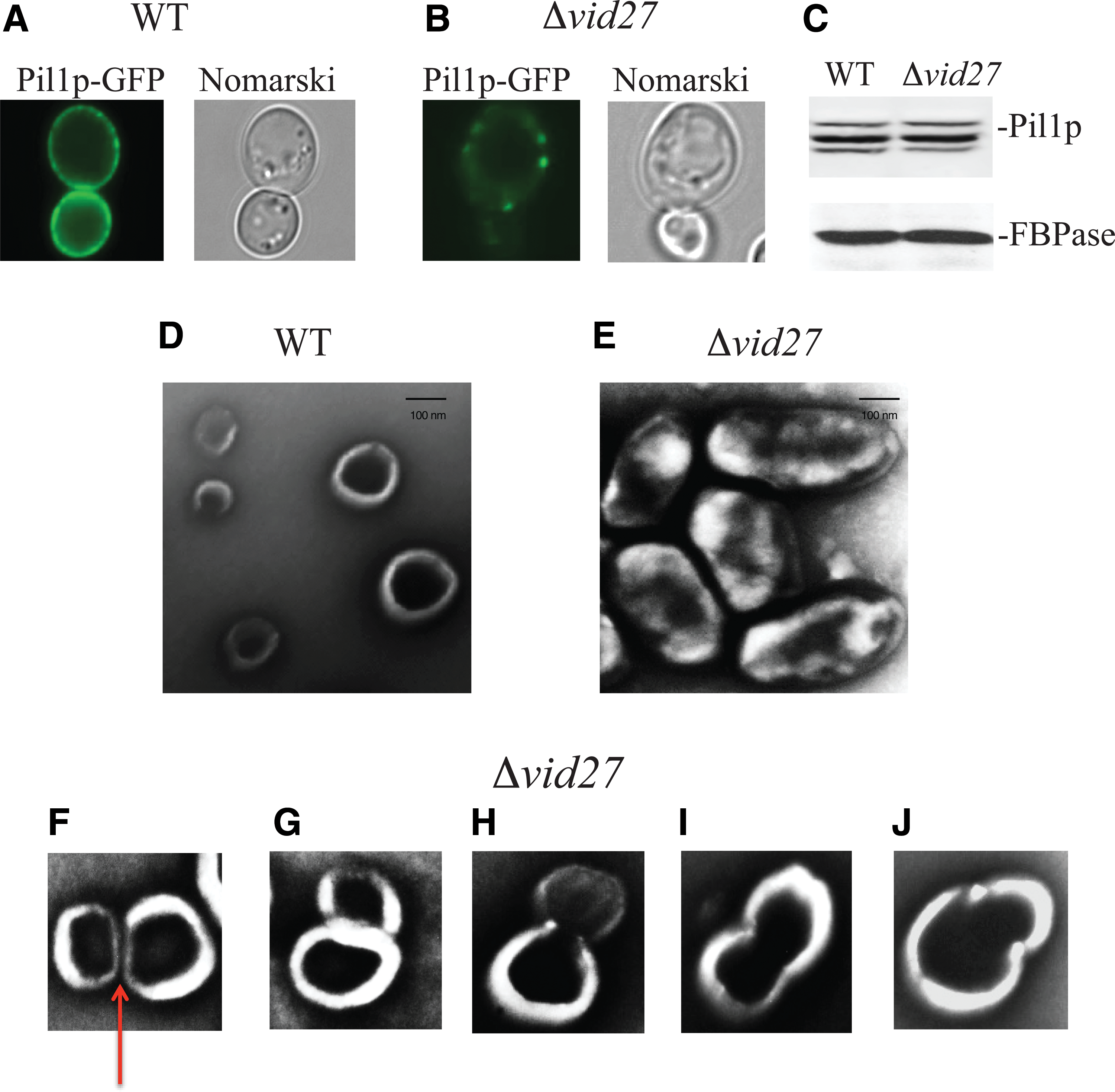
Cup-shaped organelles become elongated in the absence of VID27. Wild-type
We next examined whether changes in overall Pil1p distribution on the plasma membrane correlated with changes in the extracellular domain of Pil1p-containing organelles. Wild-type and Δvid27 mutant cells were glucose starved for 3 days and subjected to an extraction procedure that releases extracellular vesicles and cup-shaped organelles from whole cells. Following extraction, Pil1p-containing organelles were isolated, stained, and visualized using transmission electron microscopy (TEM). In wild-type cells, round-shaped organelles with relatively thin rims were found (Fig. 1D). Note that the darker areas, which may represent openings of the organelles, occupied the majority of these cup-shaped organelles. In contrast, elongated organelles (200–600 nm in length and 100–300 nm in width) were observed in the Δvid27 mutant cells (Fig. 1E). The openings were narrower and the rims were thicker when compared with those isolated from wild-type cells. The lengths of elongated structures were approximately twice those of round-shaped structures, suggesting that one elongated structure is formed from two round organelles. Along these lines, unique structures that may represent intermediate steps in the formation of elongated organelles were detected in the Δvid27 mutant. Figure 1F–J shows two round organelles in close proximity to each other, either with (Fig. 1F, arrow) or without a separating gap (G). In some cases, the darker areas were connected (Fig. 1H–J). These results indicate that VID27 functions to prevent the formation of elongated structures.
Elongated structures contain fewer vesicles and impair vesicle endocytosis
Cup-shaped organelles have been postulated to serve as extracellular domains of vesicle-conducting channels that transport vesicles from the periplasm to the cytoplasm during vesicle endocytosis (Stein et al., 2017). As such, each of these cup-shaped structures should have an opening to allow vesicles to enter from the periplasm and a second opening for vesicles to pass through. In wild-type cells, round structures with large openings were observed. These structures were either empty (Fig. 2A, left) or contained five to seven small vesicles (Fig. 2A, middle and right). In general, the vesicles appeared to localize in the deeper portions of the organelles. In certain cases, two thin lines (100–130 nm in length) were observed in the center of the organelles. These lines were in parallel (Fig. 2B, left) or formed smaller openings that were 80–100 nm in diameter (Fig. 2B, middle and right).
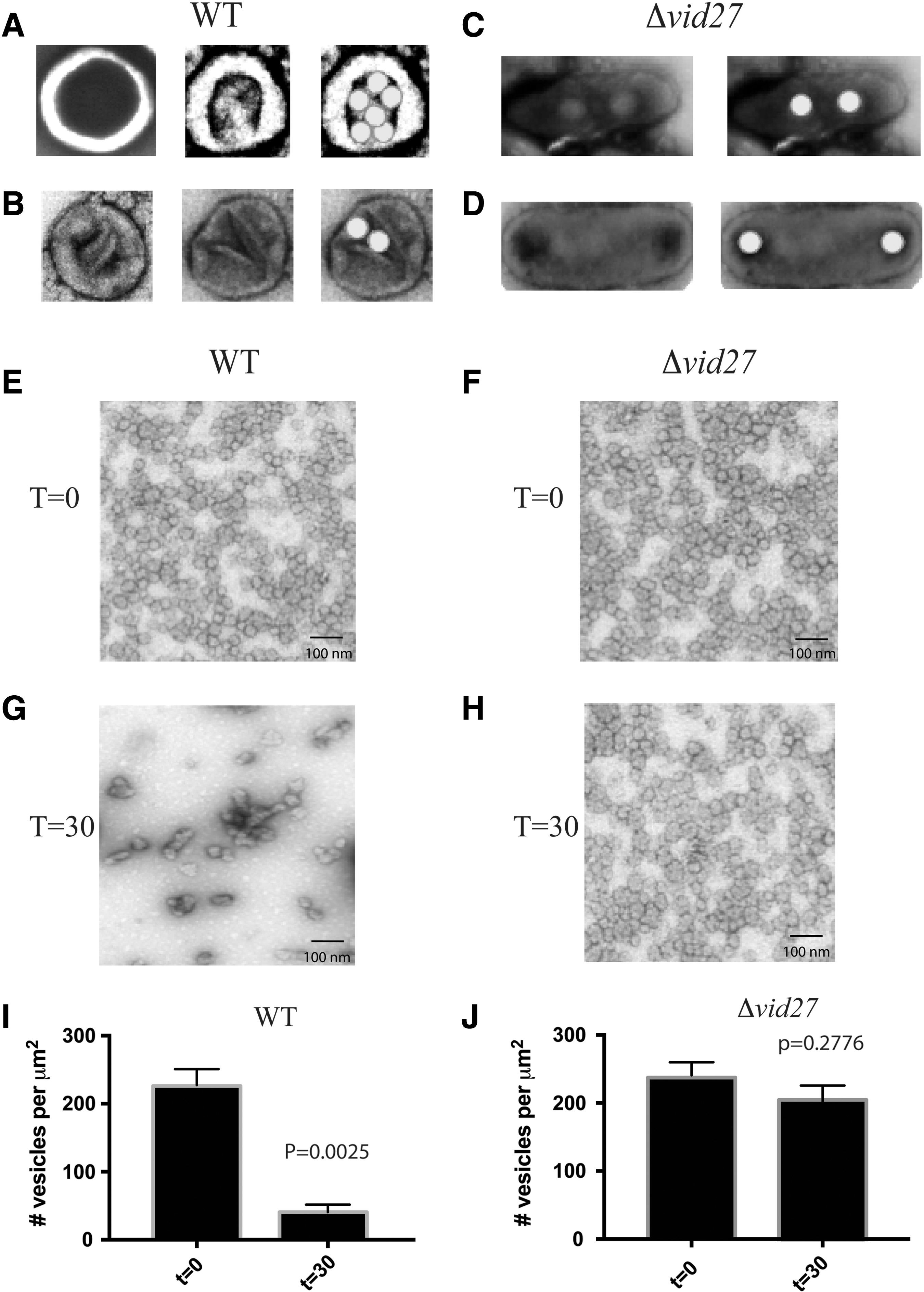
Elongated structures contain fewer vesicles and inhibit vesicle endocytosis. Wild-type
Based on our TEM results, changes in the morphology of the cup-shaped organelles may affect the ability of these structures to accommodate vesicles. Although cup-shaped organelles became elongated in the Δvid27 mutant, these structures still appeared to contain openings that allowed for the entry of extracellular vesicles. However, there was a reduction in the number of vesicles that could fit into deeper areas of the organelles. In these structures, deeper areas were continuous and contained two to four vesicles (Fig. 2C). Another unusual feature of these elongated structures was the presence of two to three openings (each opening was 40–60 nm in diameter) separated by a diaphragm (Fig. 2D).
Given that fewer vesicles were found in the deeper regions of elongated structures, we determined whether endocytosis of vesicles in response to glucose was affected. It has been reported that glucose addition to glucose-starved wild-type cells leads to a decline in the number of extracellular vesicles in the periplasm (Giardina et al., 2014a). Wild-type and Δvid27 mutant cells were glucose starved for 3 days and then transferred to high glucose for 0 and 30 min. Extracellular vesicles were obtained following the extraction procedure and examined through TEM. At t = 0, small vesicles were detected in the periplasm of wild-type (Fig. 2E) and mutant cells (Fig. 2F). However, 30 min after glucose replenishment in wild-type cells, a decrease in the number of extracellular vesicles was observed (Fig. 2G, I). In contrast, there were only slight reductions in extracellular vesicle levels in mutant cells at t = 30 min (Figs. 2H and 3J). This indicates that endocytosis of vesicles is compromised in the absence of VID27, and this is associated with changes in the shape of vesicle-containing organelles.
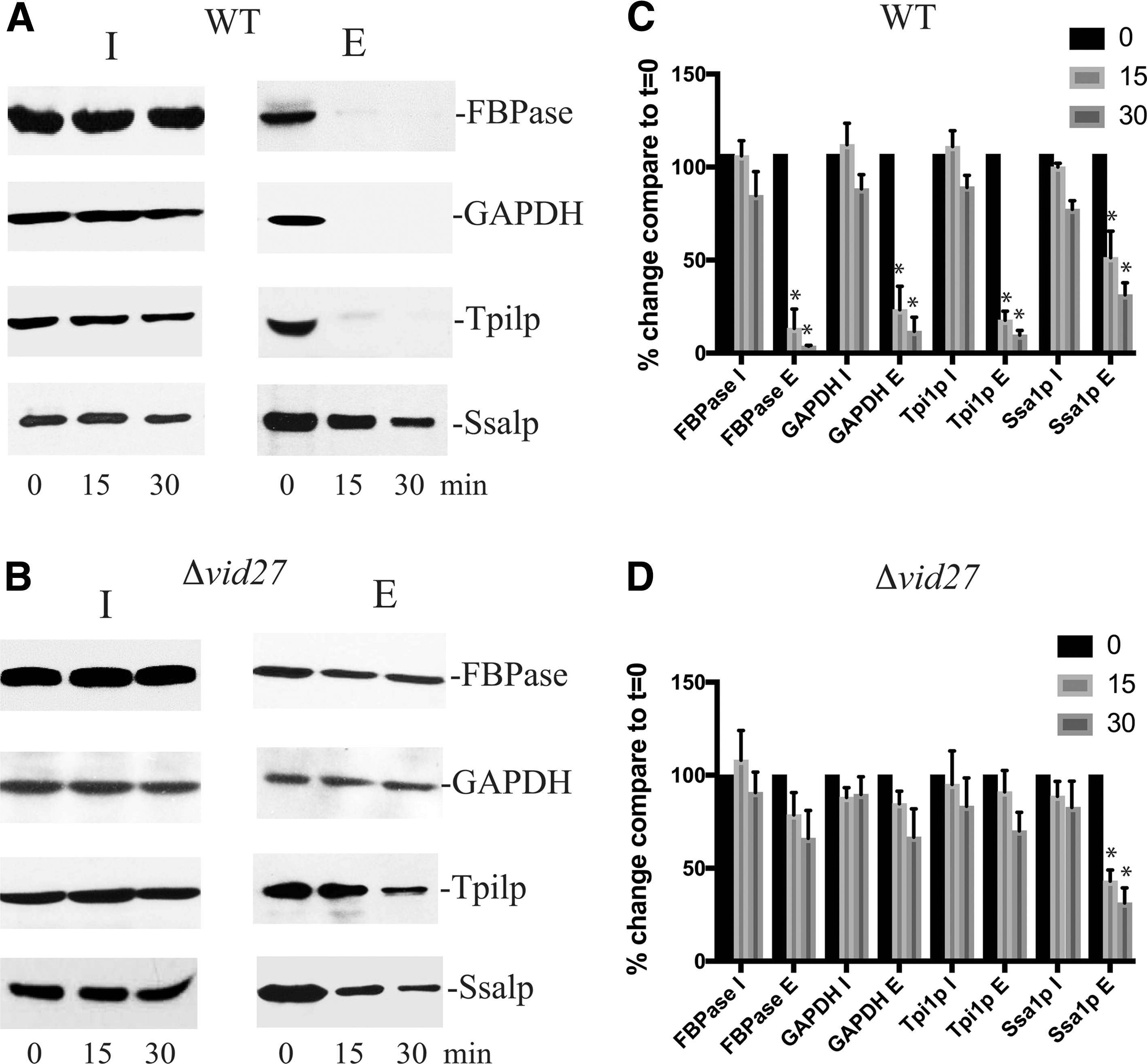
Elongated organelles impair the endocytosis of vesicles containing metabolic enzymes. Wild-type
Elongated organelles inhibit endocytosis of vesicles carrying metabolic enzymes
We next investigated whether elongated organelles affect endocytosis of vesicles containing different cargo proteins. The glucose-induced endocytosis of extracellular vesicles is divided into a fast and a slow pathway. The fast pathway is the predominant pathway used by vesicles carrying metabolic enzymes, whereas the slow pathway is the minor pathway used by vesicles carrying the 70 kDa heat shock protein Ssa1p (Stein et al., 2017). Endocytosis leads to a decrease in protein levels of vesicle-associated proteins in the extracellular fraction (Giardina et al., 2014a, 2014b). Wild-type and Δvid27 mutant cells were glucose deprived for 3 days and then refed with high glucose for 0, 15, and 30 min. Following extraction, vesicle-associated proteins released into the supernatant are referred to as proteins in the extracellular (E) fraction, whereas proteins that remain associated with cells are referred to as proteins in the intracellular (I) fraction. It has been reported that vesicles containing the metabolic enzymes FBPase, GAPDH, and Tpi1p are secreted during glucose starvation (Giardina et al., 2014a). Following glucose addition, these extracellular vesicles are internalized by the fast endocytic pathway (Stein et al., 2017). In wild-type cells, a rapid decrease of metabolic enzymes in the E fraction was observed following glucose replenishment (Fig. 3A, C). In contrast, the decline in the E fraction was impaired in cells lacking VID27 (Fig. 3B, D). Amounts of Ssa1p in the E fraction were reduced at a slower rate in both wild-type (Fig. 3A) and the Δvid27 strains (Fig. 3B). Thus, the presence of elongated structures is associated with impairment of the fast endocytic pathway used by vesicles carrying metabolic enzymes.
VID21 is critical in maintaining the shape of vesicle-containing organelles
To identify additional VID genes that have similar functions in maintaining the morphology of cup-shaped organelles, we screened for vid mutants that showed changes in Pil1p distribution on the plasma membrane. In cells lacking VID21, the distribution of Pil1p was altered (Fig. 4A), although levels of Pil1p were similar in wild-type and the Δvid21 strains (Fig. 4B). VID21 also plays an important role in maintaining the morphology of cup-shaped organelles. In cells lacking VID21, elongated organelles were found (Fig. 4C). Unique structures that contained two linked round-shaped organelles were observed (Fig. 4D–F) with dark areas that were either separated (Fig. 4D, E) or connected (Fig. 4F). In rare cases, three linked round structures were detected (Fig. 4G). Thus, VID21 also functions to prevent the formation of elongated organelles.
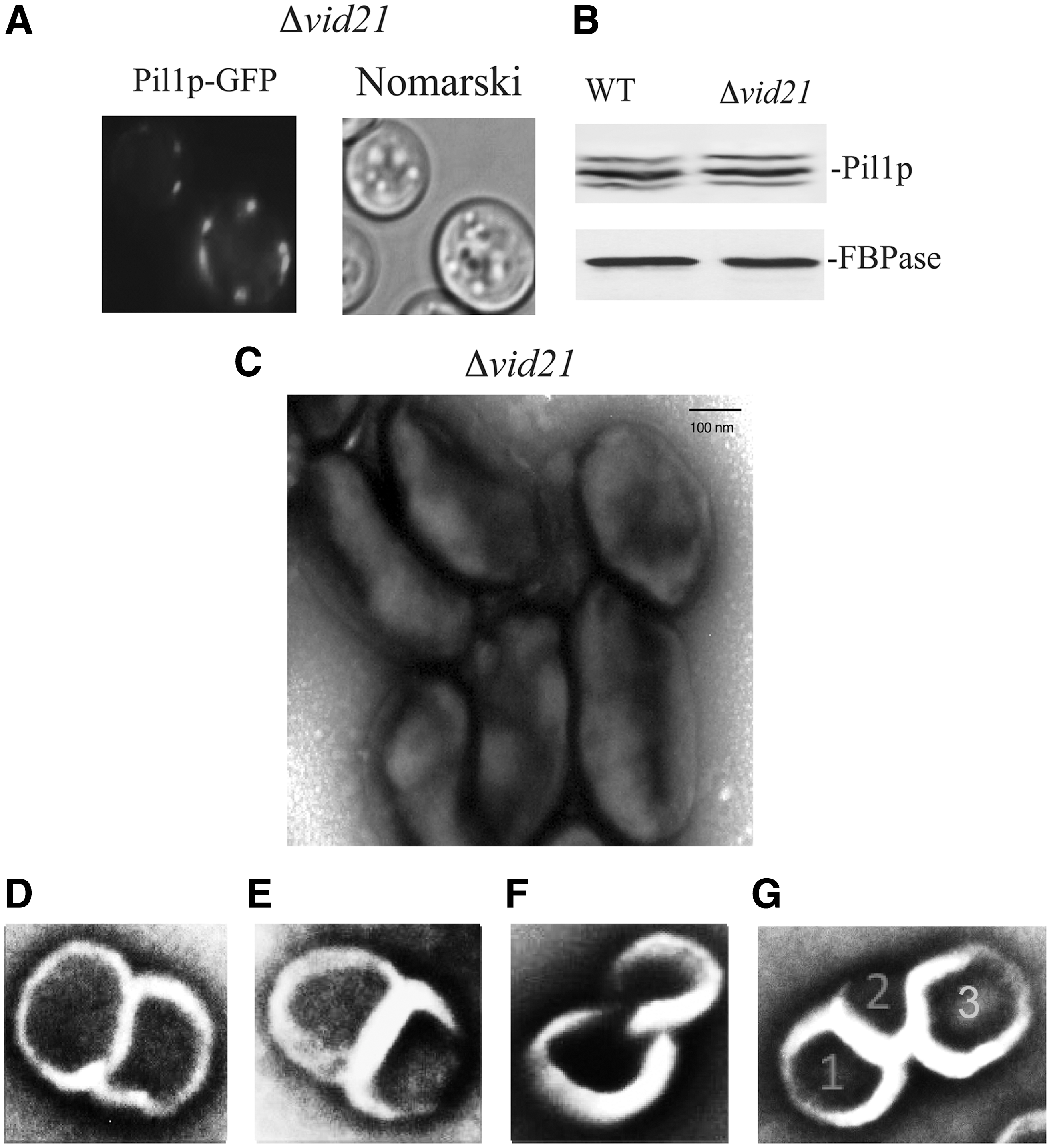
VID21 prevents the formation of elongated organelles.
Elongated organelles isolated from the Δvid21 mutant contained two to four small vesicles in the deeper areas (Fig. 5A), suggesting that the elongated morphology reduces the capacity of the organelle to take up vesicles. Furthermore, two to four openings separated by a diaphragm were also observed in elongated organelles (Fig. 5B).
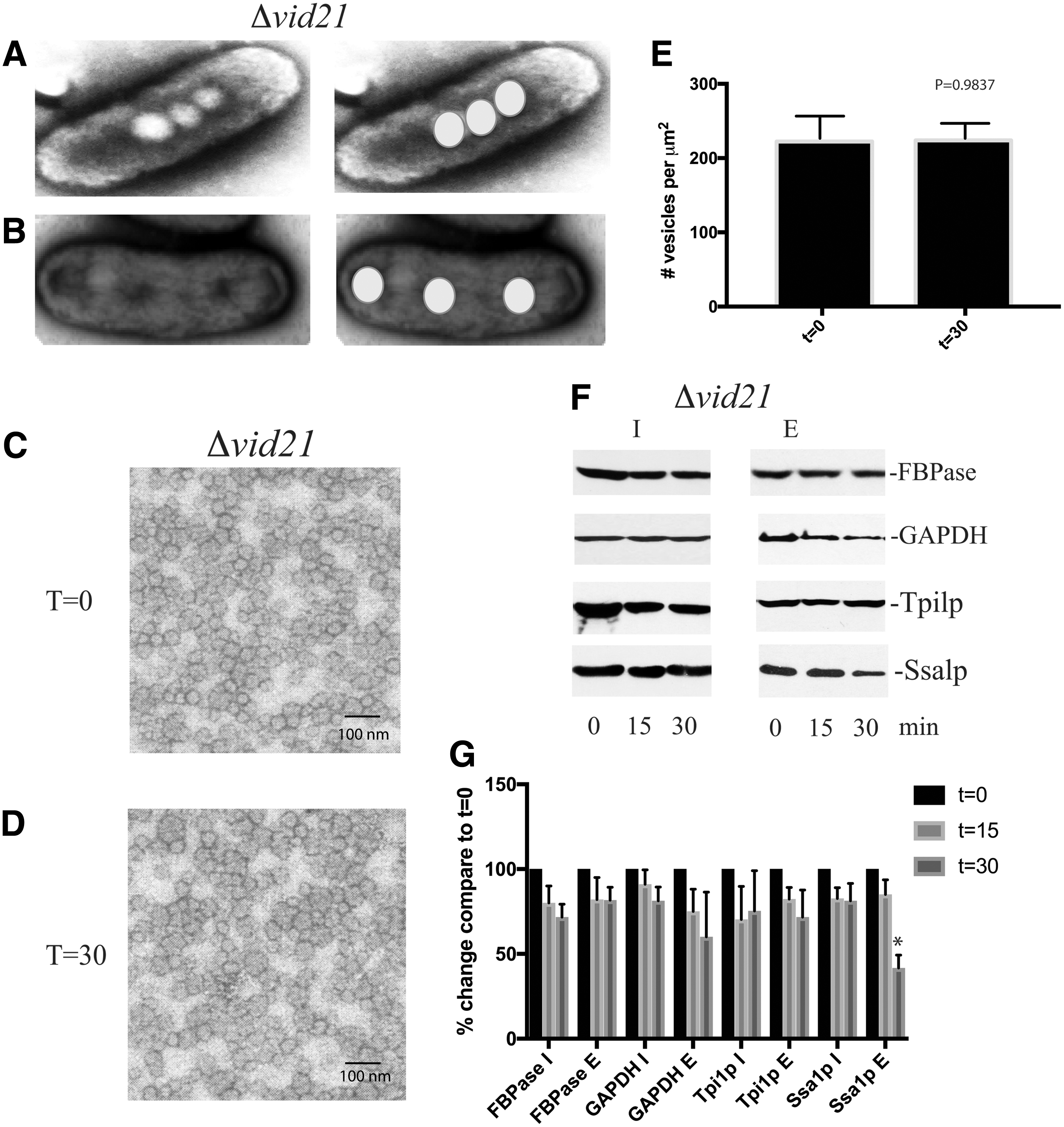
Elongated organelles from the Δvid21 mutant contain fewer vesicles and impair vesicle endocytosis.
We next determined whether endocytosis of extracellular vesicles was impaired in the Δvid21 mutant. When the Δvid21 mutant cells were starved of glucose for 3 days, small vesicles were found in the extracellular fraction (Fig. 5C). Following glucose addition for 30 min to this mutant strain, there was no change in the number of extracellular vesicles (Fig. 5D, E). Similar to our observations in Δvid27 mutant cells, the glucose-induced decline of metabolic enzymes in the E fraction was impaired in the Δvid21 mutant, whereas the slow decline of extracellular Ssa1p still occurred in this mutant strain (Fig. 5F, G). Taken together, these data indicate that VID21 plays a critical role in maintaining the morphology of cup-shaped organelles required for endocytosis of vesicles carrying metabolic enzymes.
Vid27p and Vid21p are localized to vesicle clusters and interact with other Vid proteins
To gain a better understanding of how VID27 and VID21 function to maintain normal cup-shaped morphology, we characterized Vid27p and Vid21p. Previously, we reported that some Vid proteins are constitutively expressed, whereas others are induced upon glucose replenishment to glucose-starved cells (Chiang and Chiang, 1998; Alibhoy et al., 2012; Giardina et al., 2013). Wild-type cells expressing Vid27p tagged with either V5-His6 or GFP were grown in low glucose for 3 days, transferred to glucose for the indicated time points, and examined for the expression of Vid27p. The amounts of Vid27p-V5-His6 (Fig. 6A) and Vid27p-GFP (Fig. 6B) did not increase when cells were refed with fresh glucose. However, FBPase was degraded in these strains. Thus, Vid27p is expressed constitutively and the presence of the tags does not interfere with FBPase degradation.
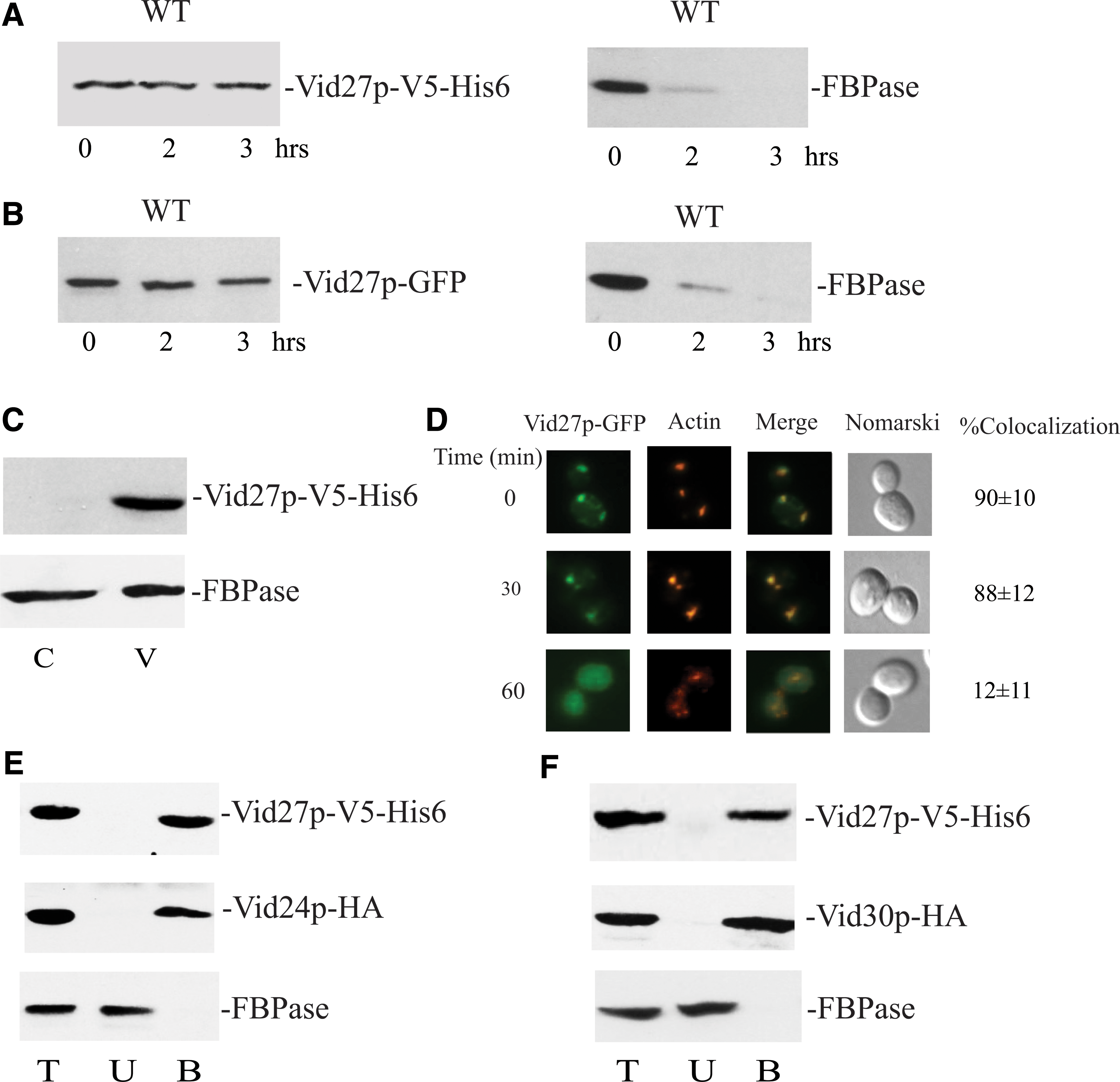
Vid27p is localized to vesicles and interacts with other Vid proteins. Wild-type cells expressing Vid27p-V5-His6
Vesicles carrying Vid proteins are distributed in free vesicles and vesicle clusters in the cytoplasm (Alibhoy et al., 2012; Giardina et al., 2013). To determine whether Vid27p is associated with free vesicles, wild-type cells expressing Vid27p-V5-His6 were starved of glucose and examined for Vid27p distribution in the vesicle-enriched pellet fraction (Fig. 6C). This protein was found in the pellet fraction enriched in free vesicles, whereas FBPase was in both the vesicle- and cytosol-enriched fractions,
Vid vesicles also aggregate to form large clusters that associate with actin patches during glucose starvation and following glucose addition for 30 min, whereas less association is found following glucose addition for 60 min (Brown et al., 2008, 2010a). To investigate the distribution of Vid27p-GFP with actin patches, a wild-type strain expressing Vid27p-GFP was glucose starved for 3 days and refed with glucose for 0, 30, and 60 min. As illustrated in Figure 6D, colocalization of Vid27p-GFP with actin patches was observed in cells maintained under the starvation condition. This association remained when cells were transferred to high glucose for 30 min. At the 60-min time point, however, Vid27-GFP exhibited less colocalization with actin patches. Thus, Vid27p is also localized to vesicle clusters.
Given that Vid27p showed distribution patterns similar to other Vid proteins such as Vid24p and Vid30p, we next determined whether Vid27p interacts with these proteins. Wild-type cells coexpressing Vid27p-V5-His6 and Vid24p-HA were grown in low glucose for 3 days and refed with glucose for 20 min. Vid27p-V5-His6 was pulled down and proteins were separated into the total (T), unbound (U), and bound (B) fractions. The distribution of proteins in these fractions was determined. Vid24p-HA was coprecipitated with Vid27p-V5-His6 in the bound fraction (Fig. 6E). In contrast, FBPase was in the unbound fraction. In wild-type cells coexpressing Vid27p-V5-His6 and Vid30p-HA, Vid30p-HA was in the bound fraction that was precipitated by Vid27p-V5-His6, whereas FBPase was in the unbound fraction (Fig. 6F). These results indicate that Vid27p interacts with Vid24p and Vid30p.
To characterize Vid21p, this protein was first tagged with V5-His6. Levels of Vid21p were similar before and after glucose addition (Fig. 7A). FBPase was degraded in this strain, indicating that this tag does not impair FBPase degradation. Similar to Vid27p, Vid21p was found in the pellet fraction enriched for free vesicles (Fig. 7B). Vid21p was also present in vesicle clusters and associated with actin patches during glucose starvation and after glucose refeeding for 30 min, whereas less association with actin patches was observed at the t = 60 min time point (Fig. 7C). In cells that expressed Vid21p-V5-His6 and Vid24p-HA (Fig. 7D), Vid24p-HA was in the bound fraction that was precipitated by Vid21p-V5-His6. In cells expressing both Vid21p-V5-His6 and Vid30p-HA, Vid30p-HA was coprecipitated with Vid21p-V5-His6 in the bound fraction. In contrast, FBPase was in the unbound fraction in both strains (Fig. 7D, E). Hence, Vid21p is localized to free vesicles and vesicle clusters. This protein is constitutively expressed and interacts with Vid24p and Vid30p.
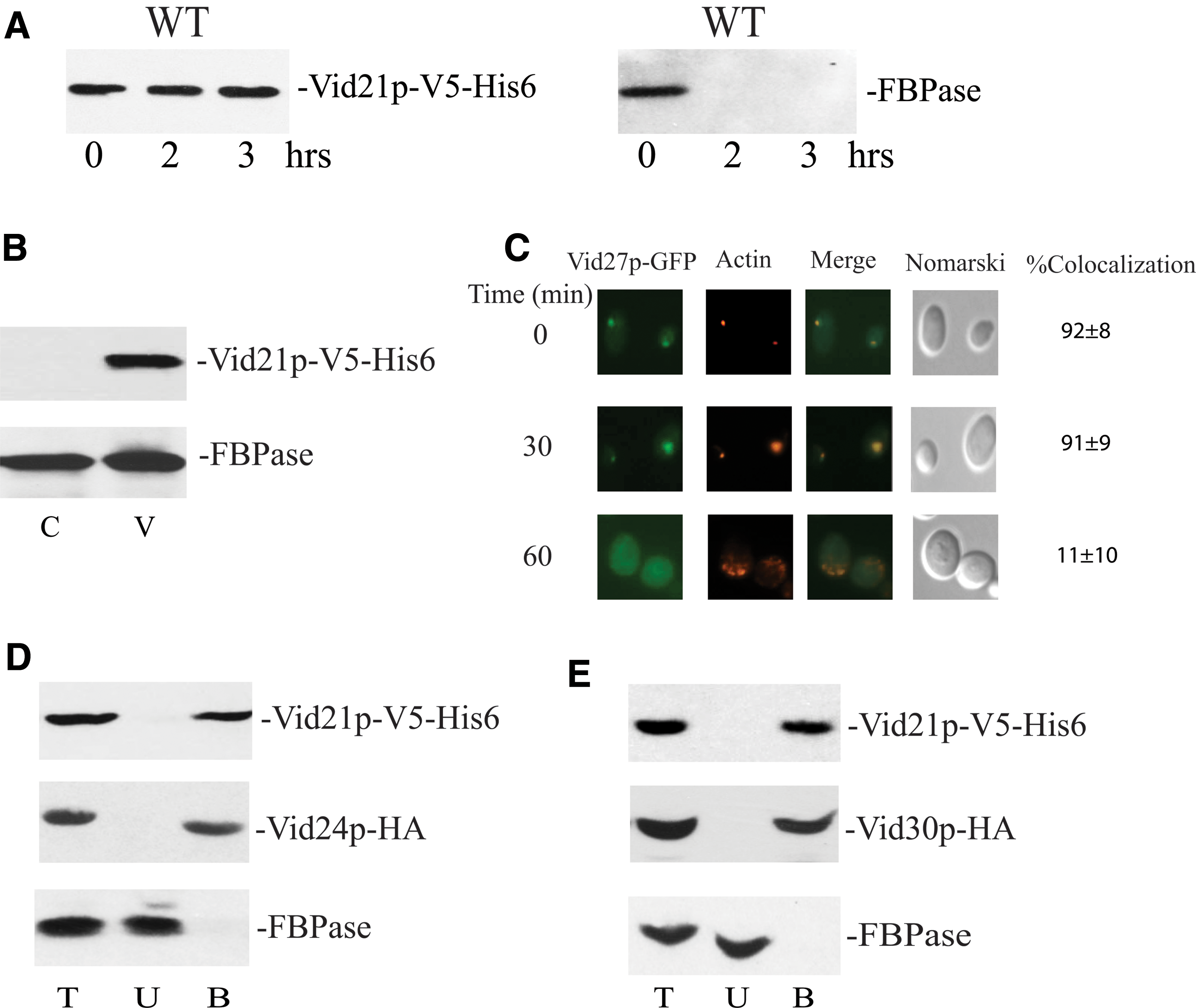
Vid21p interact with other Vid proteins and is localized to vesicle clusters.
Vid27p and Vid21p are critical in forming vesicle clusters containing other Vid proteins and retaining these vesicles in the cytoplasm
Interaction of vesicles containing Vid proteins is important for vesicles to aggregate (Giardina et al., 2013). Given that Vid27p and Vid21p interacted with other Vid proteins, we determined whether these genes have roles in forming vesicle clusters containing other Vid proteins. GFP-Vid24p was expressed in wild-type, Δvid27, and Δvid21 cells and examined for the presence of clusters containing GFP-Vid24p using fluorescence microscopy (Fig. 8A). In wild-type cells, GFP-Vid24p was found in puncta that colocalized with actin patches. In cells lacking either VID27 or VID21, GFP-Vid24p did not form puncta, although actin patches were present. Hence, VID27 and VID21 are important in forming vesicle clusters containing Vid24p.

Vid27p and Vid21p have important roles in forming vesicle clusters containing Vid proteins and retaining these vesicles in the cytoplasm.
Subtypes of vesicles are present in glucose-starved wild-type cells. For example, vesicles carrying cargo proteins such as metabolic enzymes and Ssa1p are secreted into the periplasm (Giardina et al., 2014b). In contrast, vesicles carrying Vid24p and Vid30p are retained in the cytoplasm (Giardina et al., 2013). Given that vesicles carrying Vid proteins failed to aggregate in mutant strains, these vesicles may be secreted. Wild-type, Δvid27, and Δvid21 mutant cells were glucose starved for 1–3 days and examined for the distribution of Vid24p and Vid30p in the I and E fractions. In wild-type cells, Vid24p and Vid30p were not found in the E fraction (Fig. 8B). In contrast, both proteins were detected in the E fraction of mutant strains that were starved of glucose for 3 days (Fig. 8C, D). Levels of metabolic enzymes and Ssa1p in the E fractions showed similar distribution patterns in wild-type and mutant strains (Fig. 8E–G). Hence, when vesicles containing Vid proteins failed to aggregate, they were secreted.
Discussion
Extracellular vesicles have been shown to be internalized by mechanisms, including clathrin/dynamin-1-mediated endocytosis, micropinocytosis, and phagocytosis (French et al., 2017; Maas et al., 2017; Valdinocci et al., 2017). However, a direct transport of vesicles across the plasma membrane has not been described. A recent report in yeast showed that cup-shaped structures are important in internalizing vesicles for degradation through the Vid pathway (Stein et al., 2017). In the current study, we found that the function of cup-shaped organelles in vesicle endocytosis is dependent on the shape of the structures regulated by VID21 and VID27. In wild-type cells, these cup-shaped structures were round with centers deeper than peripheries. The deletion of either VID27 or VID21 resulted in the formation of elongated organelles with longer but narrower openings that contained fewer vesicles in the deeper areas. Elongated structures were approximately twice as long as cup-shaped structures, suggesting that one elongated organelle is formed from two round-shaped structures. Along these lines, we have detected several novel morphologies that may represent various intermediate steps in forming elongated structures. For instance, we have observed two round organelles in close proximity separated by a thin gap. In other samples, there was no distinct division between the two circular structures, suggesting that the dark areas are connected.
The change in shape resulted in fewer vesicles that could fit into the deeper areas of elongated organelles. For example, in wild-type cells, five to seven small vesicles were observed in the wide openings of cup-shaped organelles. In theory, two round-shaped organelles should accommodate 10–14 vesicles. However, one elongated organelle contained only two to four vesicles located in the deeper area. Thus, if the elongated organelle was formed from two round organelles, this would result in a 60–80% reduction in the number of vesicles that each structure could take up during endocytosis. This reduced capacity could explain the defective endocytosis of extracellular vesicles seen in the Δvid21 and Δvid27 mutant strains.
Based on previous and current studies, we propose that cup-shaped structures are extracellular domains of vesicle-conducting channels required for vesicle endocytosis (labeled in green in Fig. 9A). In this model, extracellular vesicles enter each cup-shaped organelle through an opening on the periplasmic side and exit through another opening on the opposite side. A wide opening on the periplasmic side would allow more vesicles to enter the organelle. Therefore, we suggest the side that contains a wide opening with multiple vesicles in the deeper area is the entry side, and this opening accounts for 70–80% of the organelle surface. For example, if a round-shaped organelle has a diameter of 200 nm, the opening would be 130–150 nm. On the other hand, the smaller opening surrounded by two thin lines (see Fig. 2B, left) was 80–100 nm in diameter and may allow two to three vesicles to pass through. We speculate that the smaller opening is on the exit side that is connected to the transmembrane domain. When these two thin lines were aligned in parallel, they were 110–130 nm long, which may function to prevent molecules from moving in and out of the cell freely. However, these thin lines may open when cup-shaped structures are filled with vesicles. Because vesicle endocytosis is nearly completed within 15 min, these thin lines may only open for up to 15 min following glucose replenishment. Using TEM, Pil1p has been observed in furrow-like invaginations that are 300 nm long and 50 nm deep (Stradalova et al., 2009; E and K, 2012; Douglas and Konopka, 2014). The two thin lines of cup-shaped organelles appeared to protrude from the round organelles and were shorter than furrow-like invaginations. Therefore, if the two thin lines are inserted into furrows (labeled in pink in Fig. 9A) and then open upon the arrival of vesicles, complete channels may be formed. As such, vesicles may enter from a wide opening on the periplasmic side, transit to a smaller opening on the exit side, and continue to the furrow with an opening in the cytoplasm. Future experiments will be needed to test this model.
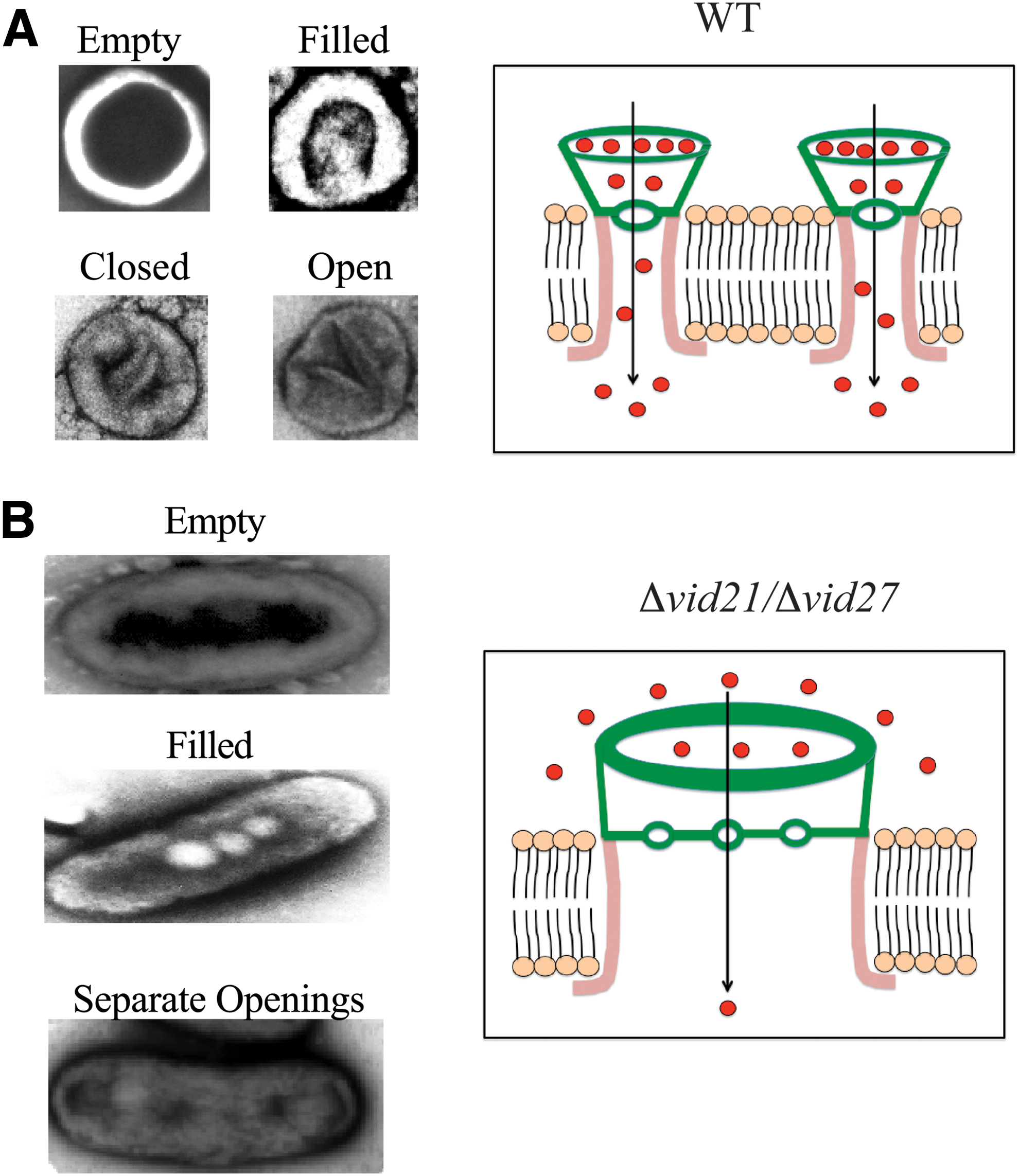
Schematic model showing how cup-shaped organelles
In cells lacking VID21 or VID27, cup-shaped organelles became elongated (Fig. 9B). The dimensions of elongated structures vary from 200 to 600 nm in length and 100–300 nm in width. For instance, an elongated structure of 400 nm long and 200 nm wide will have an opening of 280–300 nm long and 40–60 nm wide (Figs. 2C and 5A). Elongated structures with two to four openings separated by a diaphragm were also found (Figs. 2D and 5B). Each of these openings was 40–60 nm in diameter and one small vesicle (30–50 nm) could pass through each opening during the transport process. The diaphragm may serve to prevent vesicles from moving back and forth freely. Interestingly, the two thin lines that were observed in round-shaped organelles of wild-type cells (Fig. 2B, left) were not found in elongated structures of mutant cells, suggesting that these lines are excluded from these organelles.
We have shown previously that Pil1p is present in both the intracellular and extracellular fractions (Stein et al., 2017). In this study, we found a correlation between changes in the morphology of extracellular Pil1p-containing organelles at the ultrastructural level and the overall localization of Pil1p on the plasma membrane. In Δvid27 and Δvid21 mutant cells, Pil1p exhibited fewer, but larger puncta. This is in contrast to the pattern of many small punctate structures observed in wild-type cells. Pil1p/eisosomes are immobile structures on the plasma membrane (Zhang et al., 2004; Walther et al., 2006, 2007; Deng et al., 2009; E and K, 2012; Moreira et al., 2012). However, the presence of fewer, but larger puncta of Pil1p in Δvid27 and Δvid21 mutant strains may indicate an increase in Pil1p movement on the plasma membrane. When one Pil1p-containing structure encounters another one, fusion may occur leading to the formation of elongated organelles in mutant strains. This change in shape reduces their capacity to take up vesicles and impairs the endocytosis of vesicles.
Vid21p and Vid27p are essential for the formation of vesicle clusters containing Vid proteins and the retention of these vesicles in the cytoplasm. In cells lacking VID27 or VID21, vesicles containing Vid24p failed to form clusters and were secreted. We suggest that accumulation of vesicles containing Vid24p/Vid30p near the plasma membrane in mutant strains alters the microenvironment surrounding Pil1p/eisosomes. As such, increased vesicles may remove factors near the intracellular or the extracellular domain that normally restrict Pil1p movement. These changes lead to an increase in Pil1p movement and the fusion of two or more of cup-shaped organelles to form elongated organelles.
Conclusions
Vid21p and Vid27p play important roles in maintaining the shape of Pil1p-containing organelles that function as the extracellular domain of vesicle-conducting channels. In the absence of these genes, elongated structures were formed resulting in impaired vesicle endocytosis. Vid21p and Vid27p are critical for the aggregation of vesicles containing other Vid proteins. When these vesicles failed to aggregate, they were secreted. Increased secretion of vesicles containing Vid proteins correlates with an enhanced Pil1p movement on the plasma membrane. This leads to the fusion of Pil1p-containing organelles and the formation of elongated organelles with reduced capacity to take up vesicles during endocytosis.
Footnotes
Acknowledgments
The authors thank Dr. C. Randell Brown for review of the article. They also thank Dr. Dickson (University of Kentucky) for the generous gift of Pil1p antibody. Primers were synthesized at the Core Facility of the Penn State University College of Medicine. This study was supported by a Penn State Gift Fund to H.-L.C.
Authors' Contributions
C.M.W. performed and analyzed the experiments shown in this article. L.Q.H.B. analyzed data and wrote the article. H.-L.C. contributed to the writing and preparation of figures for the article. All authors approved the final version of the article.
Disclosure Statement
These authors declare that they have no conflicts of interest with the contents of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.