
Abstract
Ovarian theca cells play an indispensable role in ovarian follicular development and hormone secretion. miR-26a-5p was reported to be differentially expressed in mature and immature chicken ovaries in our previous study; however, the role of miR-26a-5p in regulating ovarian follicle function is still unclear. In this study, we demonstrated that the expression dynamics of TNRC6A mRNA in either chicken ovaries or follicles showed an opposite trend compared with that of chicken miR-26a-5p expression. miR-26a-5p inhibited TNRC6A mRNA expression by directly targeting its 3′-untranslated region in cultured chicken theca cells. Overexpression of miR-26a-5p promoted chicken follicular theca cell proliferation in vitro. Furthermore, overexpression of miR-26a-5p and knockdown of TNRC6A significantly upregulated the antiapoptotic BCL-2 gene. Taken together, this study revealed the expression dynamics of miR-26a-5p and TNRC6A in chicken ovaries and ovarian follicles and the relationship between the expression of miR-26a-5p and TNRC6A in chicken ovarian theca cells. These results suggest that miR-26a-5p facilitates chicken ovarian theca cell proliferation by targeting the TNRC6A gene.
Introduction
T
miRNAs are a class of endogenous small noncoding RNAs of approximately 22 nucleotides in size (Carrington and Ambros, 2003; Bartel, 2004) that regulate many of crucial biological processes, including organ development, cell proliferation, differentiation, apoptosis, and other pathological and physiological events (Ambros 2004; Kloosterman and Plasterk, 2006). miRNAs can posttranscriptionally regulate gene expression by targeting specific sites in the 3′-untranslated region (3′-UTR) of target genes, resulting in the decay of mRNAs or repression of their translation (Lewis et al., 2003; Ambros and Chen, 2007; Zhang et al., 2007). Many miRNAs were identified in the ovaries of mice (Ro et al., 2007; Mishima et al., 2008), human (Xu et al., 2016), cattle (Hossain et al., 2009; Tripurani et al., 2010; Huang et al., 2011; Zielak-Steciwko et al., 2014), sheep (Shen et al., 2015; Chang et al., 2016; Hu et al., 2016), pigs (Li et al., 2011), domestic fowls (Kang et al., 2013; Xu et al., 2014; Ding et al., 2015; Wu et al., 2017), and other species. Although there is evidence that these numerous ovary-expressed miRNAs may be important for regulating ovarian function, their precise roles and target genes have not been fully elucidated. Our previous study demonstrated that miR-26a-5p and TNRC6A (trinucleotide repeat containing 6A) gene show differential expression in sexually mature and immature chicken ovaries (Kang et al., 2013).
TNRC6A is a 182 kDa protein containing multiple Glycine-Tryptophan (GW) repeats in the N-terminal and also known as GW182 (Lian et al., 2006; Eystathioy et al., 2002). In vertebrate, there are three GW182 paralogs—TNRC6A, TNRC6B, and TNRC6C. GW182 family proteins are necessary components of the miRNA-mediated gene silencing complexes (miRISC) and play a key role in miRNA-mediated gene suppression (Rehwinkel et al., 2005; Ding and Han, 2007; Nishi et al., 2013). They can interact with Argonaute (AGO) proteins via their GW-rich regions in processing bodies known as P-bodies or GW-bodies, which are cytoplasmic foci involved in mRNA degradation and storage as well as RNA-based gene silencing mechanisms (Liu et al., 2005; Lazzaretti et al., 2009; Takimoto et al., 2009), However, the functions and regulatory mechanisms of miR-26a-5p and TNRC6A in chicken ovarian follicular development are largely unknown. In this study, we investigated the expression dynamics of miR-26a-5p and TNRC6A in chicken ovary and ovarian follicles and analyzed the relationship between miR-26a-5p and TNRC6A in chicken ovarian theca cell proliferation; for the first time, we showed that miR-26a-5p facilitates chicken ovarian theca cell proliferation by targeting TNRC6A.
Materials and Methods
Birds and sample collection
Single-comb white Leghorn hens at the ages of 42, 70, 90, 110, and 162 days old (egg-laying) were obtained from the Shanghai Poultry Breeding Co., Ltd. and used in this study. Birds were housed under standard conditions with food and water available ad libitum. The hens were killed by cervical dislocation, and ovarian tissues and follicles of different sizes, F1 (33–34 mm in diameter), F4 (22–24 mm in diameter), F6 (12–15 mm in diameter), Y (yellow follicle, 6–8 mm in diameter), and W (white follicles, 2–4 mm in diameter), were collected. The yolk in the follicles was carefully removed. They were frozen in liquid nitrogen immediately and used for the expression analysis of chicken miR-26a-5p and TNRC6A in the ovary at different developmental stages and different sized follicles. All animal experiments were performed in accordance with the Institutional Animal Care and Use Ethics Committee of Shandong Agricultural University.
Theca cell dispersion and culture
The five largest follicles (F1–F5) from individual hens were carefully removed after cervical dislocation and placed in cold phosphate-buffered saline (PBS). The granulosa and theca layers were isolated according to the protocol described (Gilbert et al., 1977; Tilly et al., 1991). The isolated theca layers were then minced with microdissection scissors and digested with 0.2% (w/v) collagenase II (Gibco) at 37°C for 40 min with gentle agitation in a flask. The dispersed theca cells were filtrated via a 200 mesh filter and centrifuged at 2000 × rpm for 10 min. Cell suspensions were seeded in 24-well plates at a density of 3 × 105 cells per well in cell culture medium containing M199 (HyClone) supplemented with 10% (v/v) fetal bovine serum (FBS; HyClone) and 1% (v/v) penicillin/streptomycin (Gibco). For methyl thiazolyl tetrazolium (MTT) assays, the cells were cultured in 96-well plates at a density of 5 × 104 per well. Cells were cultured at 38.5°C in an atmosphere of 95% air: 5% CO2.
Plasmid construction
For overexpression of miR-26a-5p, a pcDNA3.1-pre-miR-26a plasmid was constructed. The 565-base-pair (bp) genome segment encompassing the mature miR-26a-5p sequence (miRBase: MIMAT0001118) and its 235 bp 5′ and 309 bp 3′ flanking regions were amplified using the primers miR-26a-F/R. Then, the amplification products were cloned into KpnI and XhoI sites of the pcDNA3.1(+) vector (Invitrogen). The TNRC6A 3′-UTR luciferase reporter plasmid was generated by amplifying a 722 bp DNA fragment of the chicken TNRC6A (XM_414871) mRNA 3′-UTR harboring the putative miR-26a-5p binding sites using the primer pair TNRXbaI-F/R. The purified polymerase chain reaction (PCR) product was then inserted into the XbaI restriction sites of pGL3-promotor vector (Promega) and named TNRC6A-3′UTR. The miR-26a-5p binding site in the TNRC6A-3′UTR was mutated from “TACTTGAA” to “TACAA,” a 3 bp deletion in the target sites, using primer pairs muTNRXbaI-F/R and a QuikChange Site-Directed Mutagenesis Kit (Stratagene) and named muTNRC6A-3′UTR. The sequences of these primers are shown in Table 1. All insert fragments were confirmed by DNA sequencing.
siRNA, small interfering RNA.
Transfection and dual-luciferase reporter assay
For confirmation that TNRC6A is a target of miR-26a-5p in vitro, freshly isolated chicken ovarian theca cells from the five largest follicles (F5–F1) were seeded into 24-well plates. When the cells reached 60–70% confluence, cotransfection with 400 ng pcDNA3.1-pre-miR-26a and 200 ng TNRC6A-3′UTR or muTNRC6A-3′UTR vector, with 400 ng pcDNA3.1(+) empty vector as a control, was performed with Lipofectamine™ LTX and PLUS™ Reagent (Invitrogen) according to the manufacturer's instructions. Cells were incubated in serum-free medium for 6 h, and then, the culture medium was replaced with M199 containing 10% FBS.
After transfection for 24 h, the cells were washed using PBS twice and then harvested by adding 100 μL passive lysis buffer. Renilla and firefly luciferase activities were measured with the Dual-Luciferase Assay System (Promega) in a GloMax 20/20 Luminometer (Promega), and the firefly luciferase signal was normalized to the renilla luciferase signal for each sample. Each transfection was performed at least in triplicate. The transfection efficiency as well as miR-26a-5p and TNRC6A expression level were tested by real-time quantitative reverse transcription PCR (qRT-PCR).
RNA extraction and qRT-PCR
Total RNA from ovarian and follicle tissues or chicken ovarian theca cells that were treated with pcDNA3.1-pre-miR-26a plasmid or pcDNA3.1(+) empty vector, were isolated with TRIzol reagent (Invitrogen). For quantitative analysis of TNRC6A mRNA, the cDNA was synthesized by Transcriptor Reverse Transcriptase (Roche, Indianapolis, IN) according to the manufacturer's instructions. The qRT-PCR analysis was performed using SYBR Premix Ex Taq (TaKaRa) on a MX3000p (Stratagene) sequence detection system with primers TNRC6A-F/R. For miR-26a-5p, we used a miRcute miRNA First-Strand cDNA Synthesis Kit (Tiangen) to synthesize the cDNA of the miRNA. qRT-PCR was performed by using a miRcute miRNA qPCR Detection Kit (SYBR; Tiangen) as previously described (Kang et al., 2013). The chicken β-actin and 5s rRNA were used as internal control products, and all primer sequences are listed in Table 1. The relative expression levels were evaluated using the 2−▵▵Ct method (Livak and Schmittgen, 2001).
Cell proliferation assay
For analysis of the role of miR-26a-5p in the proliferation of chicken ovarian theca cells, MTT assays were used. Cells were seeded at 5 × 104 cells/well in a 96-well plates and transfected with pcDNA3.1-pre-miR-26a or pcDNA3.1(+) empty vector as a control. Cell proliferation was assessed at 0 (no treatment), 48, and 72 h transfection. Then, 20 μL MTT (5 mg/mL) was added to each corresponding well and incubated for an additional 4 h at 38.5°C. The media were removed, and formazan was dissolved by adding 200 μL dimethyl sulphoxide (DMSO) to each well. Optical density was measured at 490 nm. All measurements were performed in sextuplicate, and each experiment was repeated at least three times. To further verify the effects of miR-26a-5p on the chicken ovarian theca cell proliferation, we measured the expression of the antiapoptotic BCL-2 (NM_205339) gene using qRT-PCR with the primers BCL2-F/R (Table 1).
RNA interference
Three specific interference sequences targeting the TNRC6A gene were designed and synthesized by RiboBio (Guangzhou, China). The three small interfering RNAs (siRNAs) are listed in Table 1 (TNRC6A-siRNA1-3). Chicken ovarian theca cells were seeded in 24-well plates and transfected with TNRC6A-siRNA and the NC-siRNA. After 48 h, the cells were collected, and total RNA was isolated. The samples were used for selection of the most effective TNRC6A-specific siRNA and detection the TNRC6A and BCL-2 mRNA expression.
Statistical analysis
The statistical analysis was performed using SPSS v 16.0 software, and one-way analysis of variance was used to assess the significance of the results of our experiments. All data are presented as the mean ± standard error of the mean. Differences were considered significant at p < 0.05.
Results
Expression dynamics of TNRC6A in chicken ovary and follicles
The expression dynamics of chicken miR-26a-5p in different stages of the ovary and in different-sized follicles were reported in our previous study (Kang et al., 2013). TNRC6A in the chicken genome has been predicted to be a target of miR-26a-5p. Therefore, in this study, we further investigated the expression dynamics of TNRC6A mRNA in the same tissues used for miR-26a-5p expression analysis. We found that the ovary of 42-day-old chicken exhibited the lowest level of TNRC6A, and the levels increased from 70 to 110-day-old. However, its expression level was decreased in 162-day-old chicken, although expression was still higher than that in the 42-day-old chicken (p < 0.05) (Fig. 1A). Compared with our previous study on miR-26a-5p expression (Kang et al., 2013), the expression pattern of TNRC6A was completely different, even opposite. In follicles, the expression of TNRC6A mRNA was significantly higher in W than that in other follicles (p < 0.01) and then dramatically decreased in Y. Then, it increased with follicle growth and began to decrease in F1 (Fig. 1B). From Y to F1 follicles, the expression of TNRC6A also exhibited the opposite trend from that of miR-26a-5p (Kang et al., 2013).
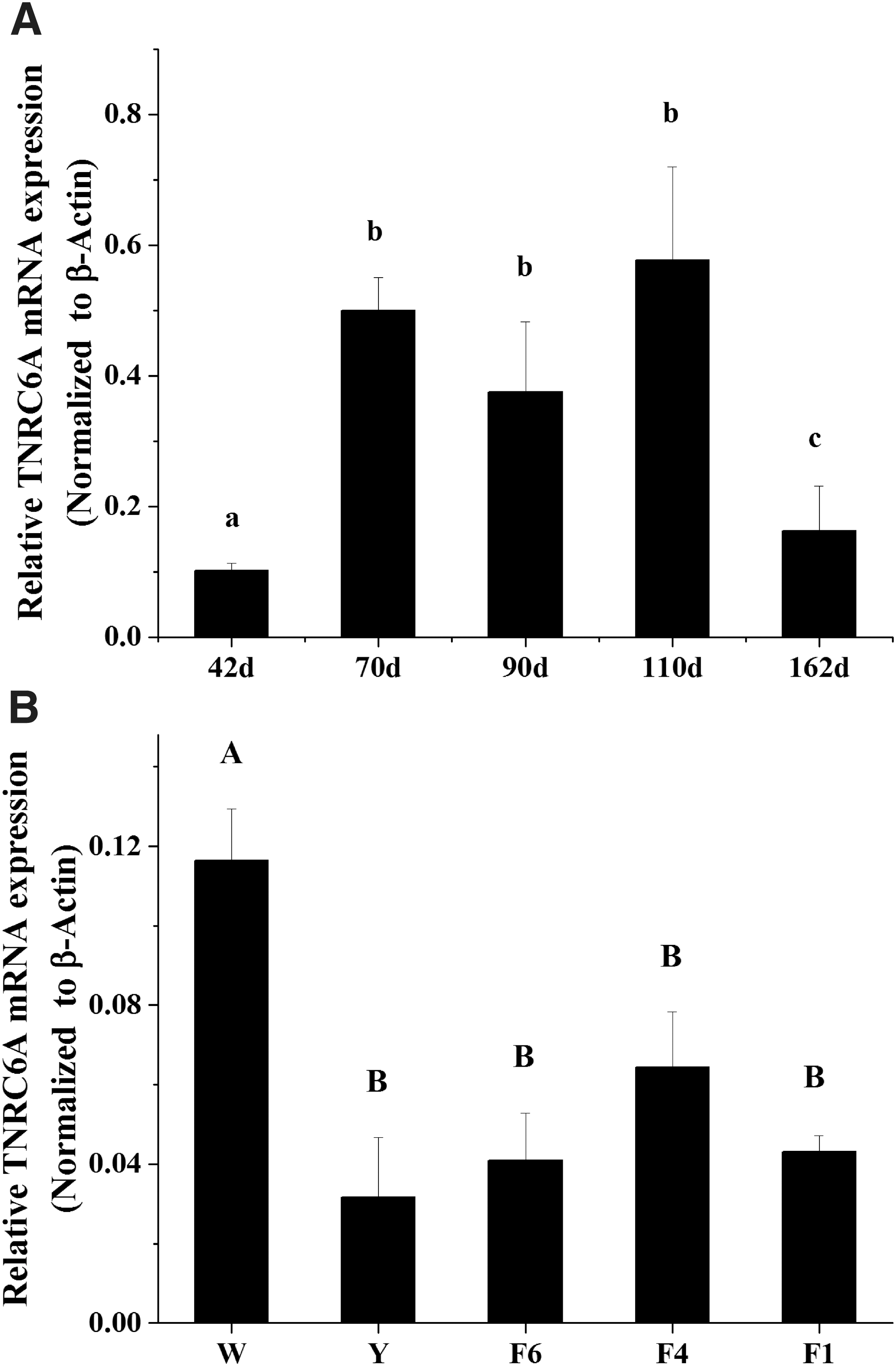
The expression pattern of chicken TNRC6A in different developmental stages of the ovary and ovarian follicles.
TNRC6A is directly targeted and regulated by miR-26a-5p in chicken ovarian theca cells
Bioinformatics prediction analysis of chicken miR-26a-5p target sequences on the TNRC6A 3′-UTR was performed by using TargetScan (
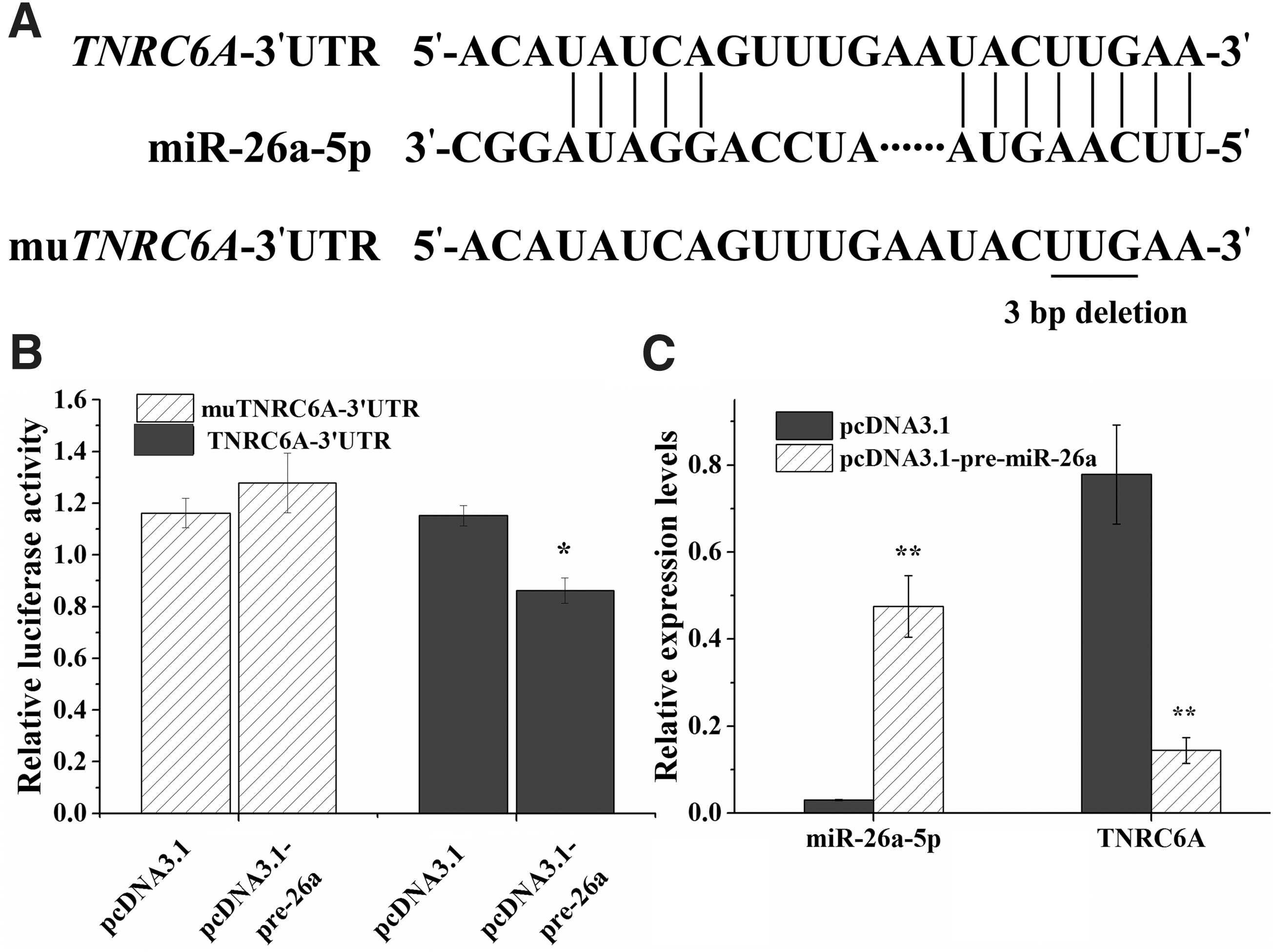
miR-26a-5p binds to the 3′-UTR of TNRC6A and regulates gene expression in chicken theca cells.
We further examined the effect of miR-26a-5p on endogenous TNRC6A expression in chicken ovarian theca cells. Overexpression of miR-26a-5p in chicken theca cells significantly decreased chicken TNRC6A expression compared with that of the pcDNA3.1 empty vector control (p < 0.01) (Fig. 2C).
Effect of miR-26a-5p overexpression on chicken ovarian theca cell proliferation
We further examined the role of miR-26a-5p on chicken theca cell proliferation by overexpressing miR-26a-5p. MTT assays of chicken ovarian theca cells transfected with the pcDNA3.1-pre-miR-26a vector and pcDNA3.1 empty vector revealed that overexpression of chicken miR-26a-5p significantly increased cell proliferation at 72 h (p < 0.05) (Fig. 3).
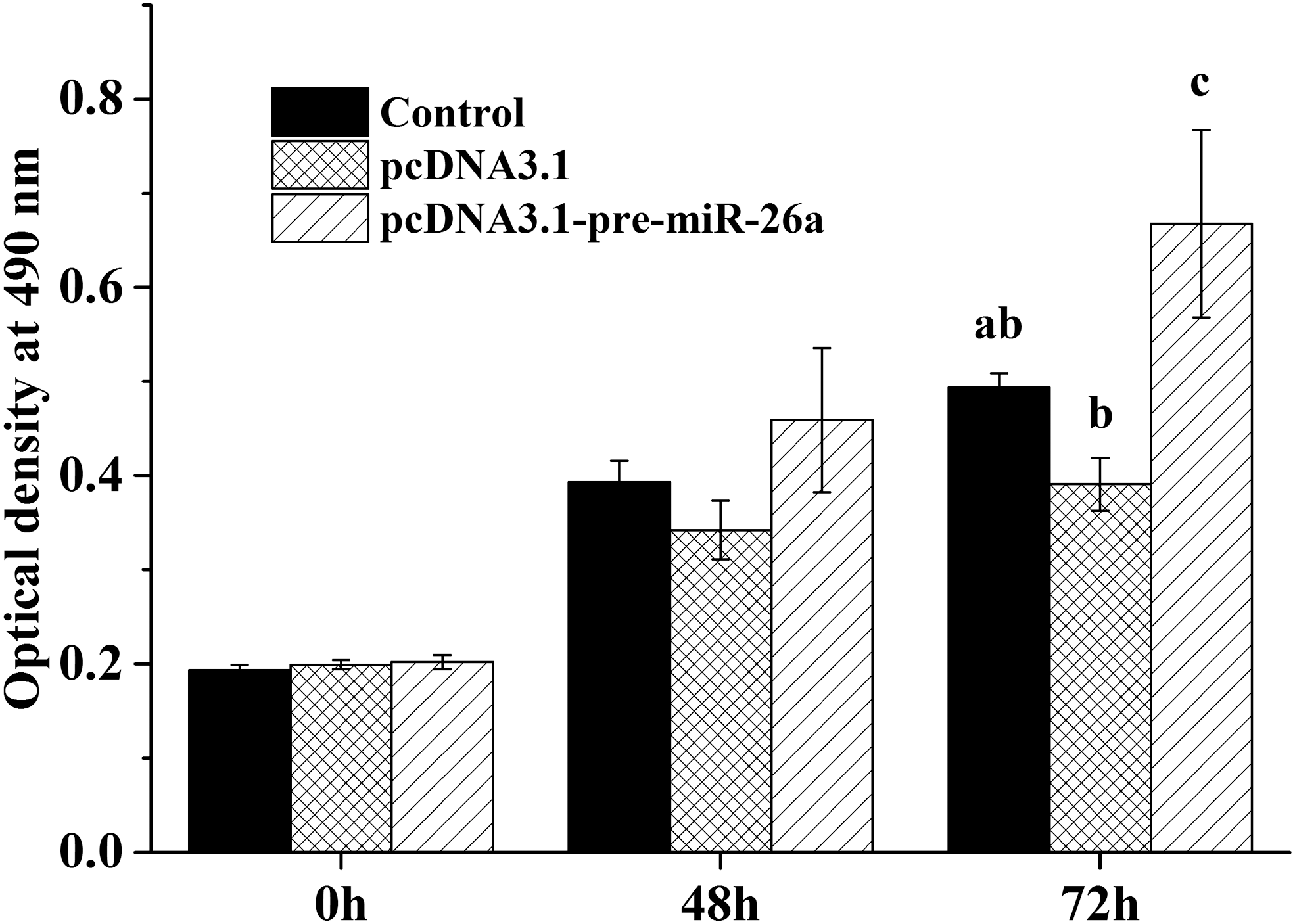
miR-26a-5p promotes the proliferation of chicken theca cells. Theca cells were seeded at 5 × 104 cells/well in a 96-well plate and transfected with pcDNA3.1-pre-miR-26a vector and pcDNA3.1 empty vector and cultured for 0 (no treatment), 48, and 72 h. Then, the optical density at 490 nm was detected. The control group was untreated. The data represent the mean ± SEM from at least five repeat experiments, and different lowercase letters represent significant differences at p < 0.05.
Overexpression of miR-26a-5p upregulates BCL-2 mRNA in chicken ovarian theca cells
We also examined the effect of miR-26a-5p on BCL-2 mRNA expression in chicken ovarian theca cells. The results indicated that overexpression of miR-26a-5p significantly increased the expression level of the antiapoptotic BCL-2 gene (p < 0.01) (Fig. 4), suggesting that miR-26a-5p likely promotes chicken ovarian follicle theca cell proliferation by inhibiting apoptosis.
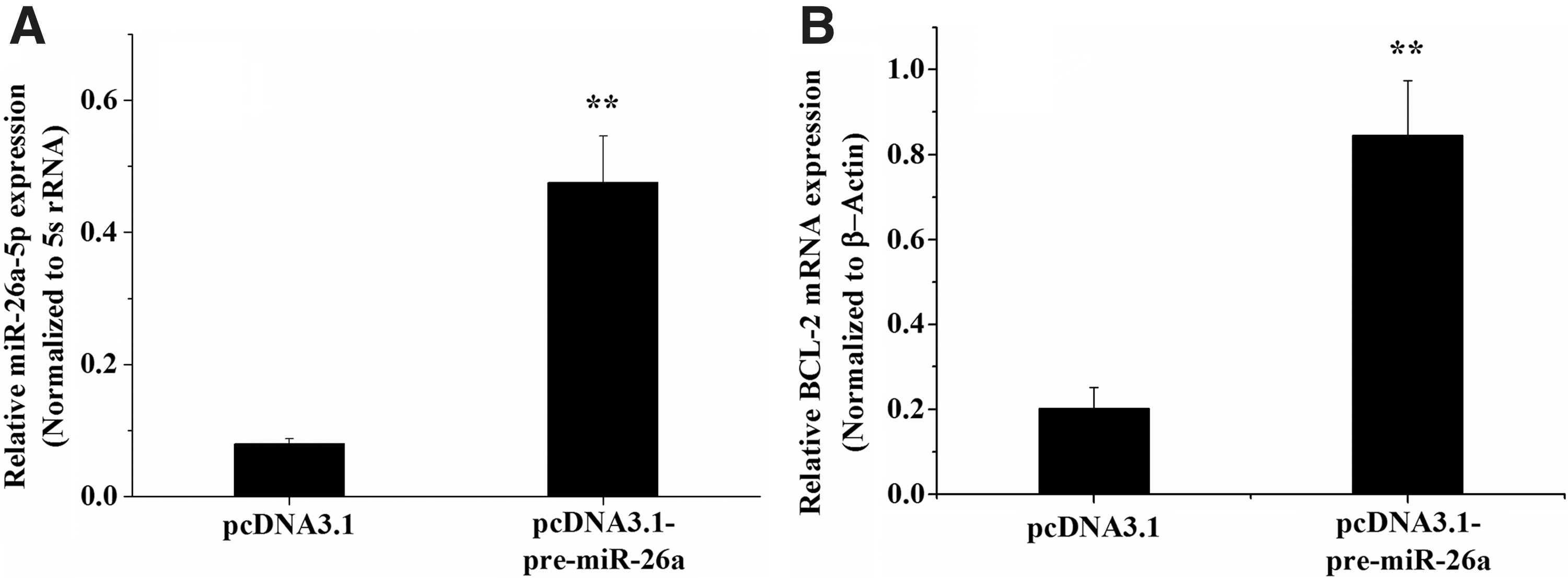
Overexpression of miR-26a-5p upregulates BCL-2 mRNA in chicken ovarian theca cells.
Inhibition of TNRC6A promotes BCL-2 expression in chicken ovarian theca cells
To further confirm the effect of TNRC6A on chicken ovarian theca cell function, we used RNA interference (RNAi) to knockdown its expression in chicken ovarian theca cells. The TNRC6A mRNA levels decreased significantly in transfected TNRC6A-siRNA2 cells compared with those of NC-siRNA-transfected cells (p < 0.05) (Fig. 5A). The expression levels of BCL-2 mRNA were substantially increased in the cells after transfection by TNRC6A-siRNA2 (p < 0.05) (Fig. 5B).
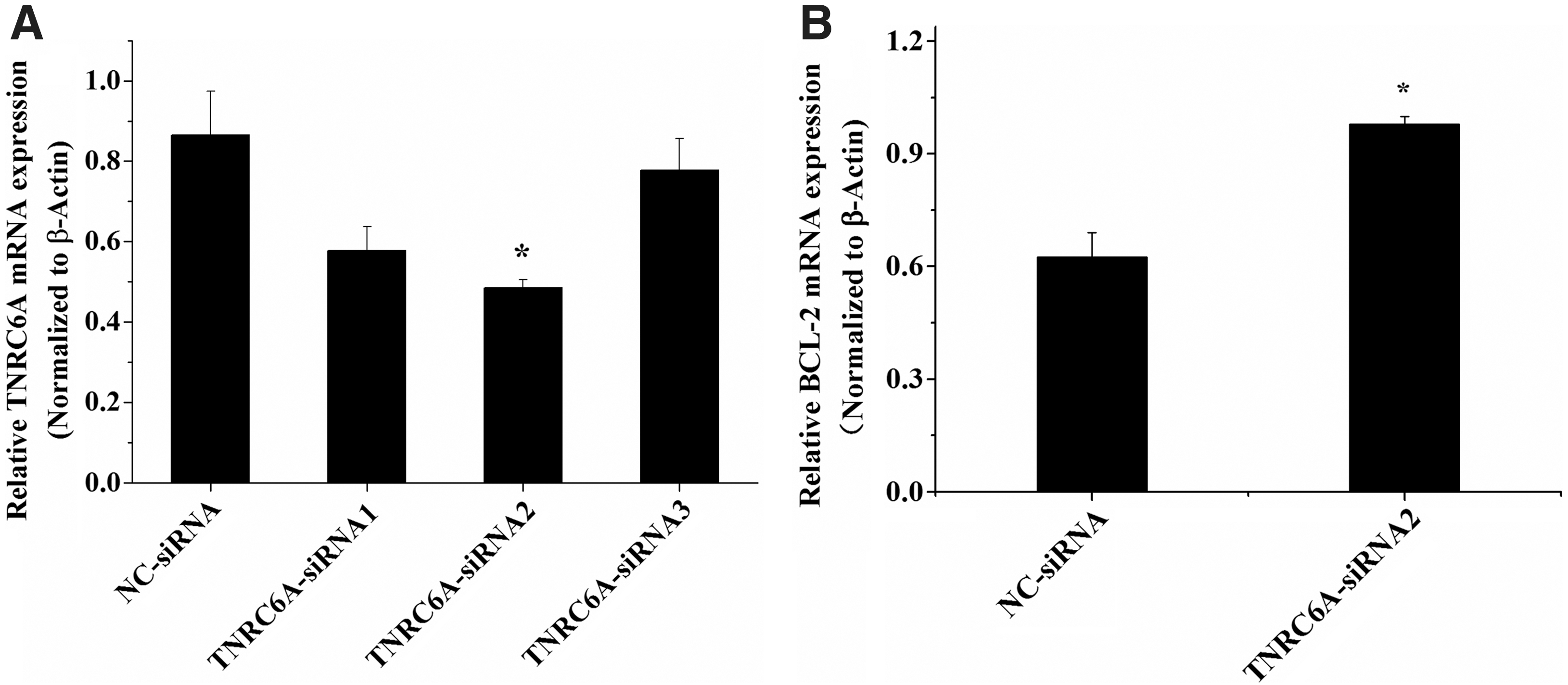
Inhibition of TNRC6A expression upregulates BCL-2 mRNA in chicken ovarian theca cells.
Discussion
In poultry, the ovary is an important organ affecting egg production. Thus, it is necessary to understand the molecular regulatory mechanisms involved in ovarian function. miRNAs have emerged as major regulators of a variety of biological processes. Genome-wide analyses of miRNA in ovarian tissues and follicle cells have been reported in several species, and multiple miRNAs were found to play critical roles in ovarian follicle development and steroid hormone secretion. In mouse ovary, miR-224 directly targets SMAD4 to promote GC proliferation (Yao et al., 2010), and miR-383 functions to increase estradiol release in GCs by targeting RBMS1 (Yin et al., 2012). In porcine GCs, miR-378 can regulate estradiol production by targeting aromatase (Xu et al., 2011). A recent study showed that miR-183/-96/-182 cluster miRNAs target suppression of FOXO1 expression and promote bovine GC proliferation and cell cycle transition (Gebremedhn et al., 2016). Another study demonstrated that miR-26b regulates GC apoptosis by directly targeting the SMAD4 gene in the porcine ovary (Liu et al., 2014).
miR-26a and miR-26b belong to the miR-26 family. Using high-throughput sequencing, our previous study found that miR-26a was a highly abundant miRNA (Kang et al., 2013) and was also found in human, mouse, and cow ovary (Ro et al., 2007; Zielak-Steciwko et al., 2014; Xu et al., 2016). These studies suggest that miR-26a may play an important role in ovarian function. However, its function in ovarian cells was unclear. In this study, expression analysis showed that miR-26a-5p expression was inversely related to the expression of TNRC6A mRNA not only in different stage ovaries but also in different-sized follicles. TNRC6A is an essential protein involved in miRNA-mediated gene suppression. The expression dynamics of the chicken TNRC6A gene in the ovary during sexual maturation and chicken ovarian follicle growth (Fig. 1) suggest an important role in female chicken reproduction.
By using dual-luciferase reporter gene assay, we showed that miR-26a-5p could inhibit the luciferase activity, while mutation of the predicted miR-26a-5p binding site in the 3′-UTR of TNRC6A abolished the inhibitory effect on luciferase activities. Moreover, overexpression of miR-26a-5p in chicken ovarian theca cells decreased TNRC6A mRNA expression. These results suggest that TNRC6A is a target gene of miR-26a-5p in chicken ovarian theca cells.
Subsequently, we used an in vitro theca cell culture model to investigate its role. miR-26a-5p was overexpressed by transfecting pre-miR-26a into chicken theca cells, and the results showed that miR-26a can promote chicken ovarian theca cell proliferation. These results were further validated by detecting the expression of the antiapoptotic gene BCL-2. BCL-2 plays a supportive role in regulating ovarian follicle cell proliferation and/or differentiation (Johnson et al., 1997; Flaws et al., 2001). Our finding that the BCL-2 mRNA level was upregulated in pre-miR-26a plasmid-transfected cells compared with that of control-transfected cells and was consistent with its role in promoting theca cell proliferation. Theca cells play a significant role in follicular growth, steroidogenesis, and follicular formation. It is thought to be recruited from surrounding stromal tissue by factors secreted from an activated primary follicle (Young and McNeilly, 2010), but the control of recruitment and function of theca cells are not yet fully understood.
miR-26a has been identified as an important tumor regulator and is involved in diverse human cancers (Chen et al., 2016). For example, in hepatocellular carcinoma, miR-26a directly inhibited the mRNA level of the ERα gene through its 3′-UTR and prevented hepatoma cell growth (Chen et al., 2011). In addition, recent studies have shown that miR-26a regulates osteogenesis of human adipose-derived mesenchymal stem cells by directly targeting the 3′-UTR of the GSK-3β mRNA (Wang et al., 2015). Another study found that miR-26a regulates the vascular smooth muscle cell function by targeting SMAD-1 (Leeper et al., 2011). However, none of these studies examined ovarian follicle function. Ovarian follicular development requires oocyte–granulosa–theca cell interactions. Functional studies of miRNA in chicken follicular theca cells are lacking compared with those of GCs. To confirm that miR-26a-5p targets TNRC6A mRNA, we transiently transfected the TNRC6A-siRNA into chicken ovarian theca cells, and the mRNA levels of BCL-2 were increased significantly compared with those of NC-siRNA. This result is consistent with miR-26a-5p overexpression in chicken ovarian theca cells.
In conclusion, miR-26a-5p can inhibit TNRC6A expression in chicken ovarian theca cells by directly targeting the TNRC6A mRNA seed sequence in the 3′-UTR and promote chicken ovarian theca cell proliferation in vitro. Overexpression of miR-26a-5p or knockdown of TNRC6A upregulated the expression of BCL-2. Our findings provide novel mechanistic insights into the role of miR-26a-5p in follicular theca cell proliferation in the chicken ovary. These results suggest that both miR-26a-5p and TNRC6A play important roles in chicken ovarian follicular development and function.
Footnotes
Acknowledgments
We thank Dr. Huaxiang Yan and Dr. Changsuo Yang at the Shanghai Poultry Breeding Co., Ltd. for providing the White Leghorn chickens. This study was funded by the National Natural Science Foundation of China (31301962, 30871777), the Project of Shandong Province Higher Educational Science and Technology Program of China (J12LF02), the Shandong Province Natural Science Foundation of China (ZR2013CM014, ZR2014CZ003), and Funds of Shandong “Double Tops” Program (SYL2017YSTD12).
Disclosure Statement
No competing financial interests exist.