
Abstract
Phosphatase and tensin homolog deleted on chromosome ten (PTEN) is a lipid and protein phosphatase and possesses an antitumor effect in lung cancers. miRNAs are reportedly abnormally expressed in human lung cancers. However, whether miRNA contributes to PTEN expression in non-small cell lung cancers (NSCLCs) has not been clearly clarified. In the present study, we found that miR-1297 probably binds with 3′UTR sequence of PTEN and negatively regulated the levels of PTEN in NSCLC cells. First, the expression levels of PTEN and Skp2 were detected by western blotting in NSCLC specimens and paired normal tissue specimens. The results showed that decreased levels of PTEN were detected in NSCLC tissues, compared with paired control tissues (**p < 0.01). The expression levels of PTEN were conversely correlated with the levels of Skp2 in clinical NSCLC specimens and NSCLC cell line. Transfection with miR-1297 mimic significantly promoted cell viability of A549 cells and NCI-H460 cells by downregulating the level of PTEN and upregulating the expression of Skp2. Interestingly, knockdown of Skp2 did not affect the expression of PTEN in A549 cells. Thus, miR-1297 might work as an oncogene by regulating PTEN/Akt/Skp2 signaling pathway in NSCLC cells. PTEN and Skp2 might be the potential targets in the clinical therapy of lung cancers.
Introduction
L
Tumorigenesis includes a complex series of processes, including inactivation of tumor suppressor genes and activation of oncogenes (Fu et al., 2015; Shao et al., 2016). The phosphatase and tensin homolog deleted on chromosome ten (PTEN) has a lipid and protein phosphatase activity and works as a tumor suppressor in cell proliferation, survival, metastasis, and invasion of tumor cells. In small cell lung carcinoma (SCLC) cells, the inactivation of PTEN accelerated the progression of SCLC in a mouse model (Cui et al., 2014). Till now, PTEN is a key protein and regulated by miRNAs involving in the progression of lung cancer cells. For example, anti-miR-21 increased the expression of PTEN mRNA and protein to inhibit growth, migration, and invasion of NSCLC cells (Liu et al., 2013). Some miRNAs promoted lung cancer progression through PTEN related signaling pathway by target PTEN, such as miR-26a (Liu et al., 2012), miR-92a (Ren et al., 2016), miR-155 (Xue et al., 2016), miR-454 (Zhu et al., 2016), and so on.
It has been reported that PTEN regulated multiple cellular functions, such as cell proliferation, differentiation, survival, motility, invasion, and intracellular trafficking, through PTEN/PI3K/AKT signaling pathway in tumorigenesis of lung cancer cells (Perez-Ramirez et al., 2015). In the present study, we found that miR-1297 might probably bind with 3′UTR sequence of PTEN, however, how miR-1297 regulated PTEN in tumor cell progression of lung cancers was still not clarified till now. We identified the role of miR-1297 in the cell proliferation of NSCLC cells and also explored the role of PTEN in the regulation of Skp2, as well as the involving molecular mechanism in human NSCLC cells, which would give new clues on the clinical therapy for human NSCLCs.
Materials and Methods
Patients
Four groups of NSCLC specimens and paired normal control tissue specimens were collected from the First Affiliated Hospital of Chongqing Medical University from October 2015 to September 2016. The patients were not given chemotherapy or irradiation before. The samples were quickly frozen at liquid N2 and kept at −80°C.
Ethical approval
The patients were well informed and signed the relevant contracts before the experiment. The studies carried out on humans were in compliance with the Helsinki Declaration and approved by the Ethics Committee of the First Affiliated Hospital of Chongqing Medical University.
Cell lines and reagents
Dulbecco's modified Eagle's medium and fetal bovine serum (FBS) were purchased from HyClone Corporation (Logan, UT). Human adenocarcinoma cell line A549 and lung large cell carcinoma H460 were cultured in DMEM containing 10% FBS, at 37°C in a humidified 5% CO2 atmosphere. MTT was obtained from Sigma Aldrich (St. Louis, MO). PTEN ORFs were cloned in frame into pcDNA3.1 (+) plasmid and transfected into lung cancer cells. Skp2 p45 shRNA Plasmid (h) (Cat. No. sc-36499-SH) and control shRNA plasmid (Cat. No. sc-108060) were purchased from Santa Cruz Corporation. hsa-miR-1297 miRNA mimic (Cat. No. MCH01244) and miRNA mimic negative control (Cat. No. MCH000000) and hsa-miR-1297 miRNA inhibitor (Cat. No. MIH01244) and miRNA inhibitor negative control (Cat. No. MIH000000) were purchased from Applied Biological Materials, Inc. (Richmond, Canada).
Immunofluorescence observation
A549 cells were transfected with hsa-miR-1297 inhibitor and inhibitor negative control for 24 h. The cells were washed with PBS buffer, fixed with 100% methanol for 5 min, and permeabilized with 0.1% Triton X-100 for 5 min. The cells were then blocked in 2% BSA in 0.1% PBS-Tween for 1 h to block nonspecific protein–protein interactions. Then, primary PTEN antibody was added and incubated overnight at 4°C. Anti-PTEN[A2b1] antibody was purchased from Abcam Corporation. The catalog number is ab79156. The secondary antibody (green) was ab150113 Alexa Fluor® 488 goat anti-mouse IgG (H+L) used at 2 μg/mL for 1 h. DAPI was used to stain the cell nuclei (blue) at a concentration of 1.43 μM. The image was observed by fluorescence confocal microscopy at 200 × .
MTT assay
The lung cancer cells were plated into 48-well plate and transfected with PTEN shRNA and control shRNA, or Skp2 shRNA and control shRNA for 24, 48, and 72 h, respectively. MTT assay was performed to test the cell viability of cancer cells as described (Wang et al., 2015; Iskandarani et al., 2016). All the data were read on a microplate reader at a test wavelength of 490 nm using the iMark microplate reader (Bio-Rad).
Western blot analysis
Cells were lysed by RIPA lysis buffer in PTEN or Skp2 shRNA transfected cells and control shRNA transfected cells. PTEN or Skp2 expression was detected by western blotting as described (Gao et al., 2015; Liang et al., 2015). All the antibodies used were listed as follows: anti-PTEN antibody [Y184] (ab32199) was a rabbit monoclonal [Y184] to PTEN, which was purchased from Abcam Corporation. Anti-SKP2 antibody (ab19877) was a rabbit polyclonal to SKP2 and purchased from Abcam Corporation. Anti-p-Akt1/2/3 antibody (A-12) (Cat. No. sc-271964) was a rabbit polyclonal IgG provided at 100 μg/mL and purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Anti-pan-AKT antibody (ab8805) was a rabbit polyclonal to pan-AKT and obtained from Abcam Corporation. β-Actin Antibody (C4): sc-47778 was a mouse monoclonal IgG1 provided at 200 μg/mL and purchased from Santa Cruz Biotechnology. The brightness of each band was scanned by gel image analysis system, and the gray value of the bands was detected and calculated automatically by the software.
Statistical analysis
The data were analyzed with t-test analysis. All the results were shown as mean ± standard deviation. A value of p ≤ 0.05 was considered statistically different.
Results
PTEN is negatively correlated with the expression of Skp2 in NSCLC tissues and normal control tissues
It has been reported that PTEN exerts tumor suppressor functions, and Skp2 is a well-known oncogene in various cancers. To investigate the association between PTEN and Skp2 in human NSCLC tissues, the expression levels of PTEN and Skp2 were determined by western blot analysis. As shown in Figure 1, the levels of PTEN in NSCLC specimens were obviously decreased compared with normal control specimens. However, the expression of Skp2 in NSCLC specimens was highly increased compared with normal control specimens. The data demonstrated that the expression level of PTEN negatively correlated with Skp2, which probably had a converse function in the development and progression of NSCLC.
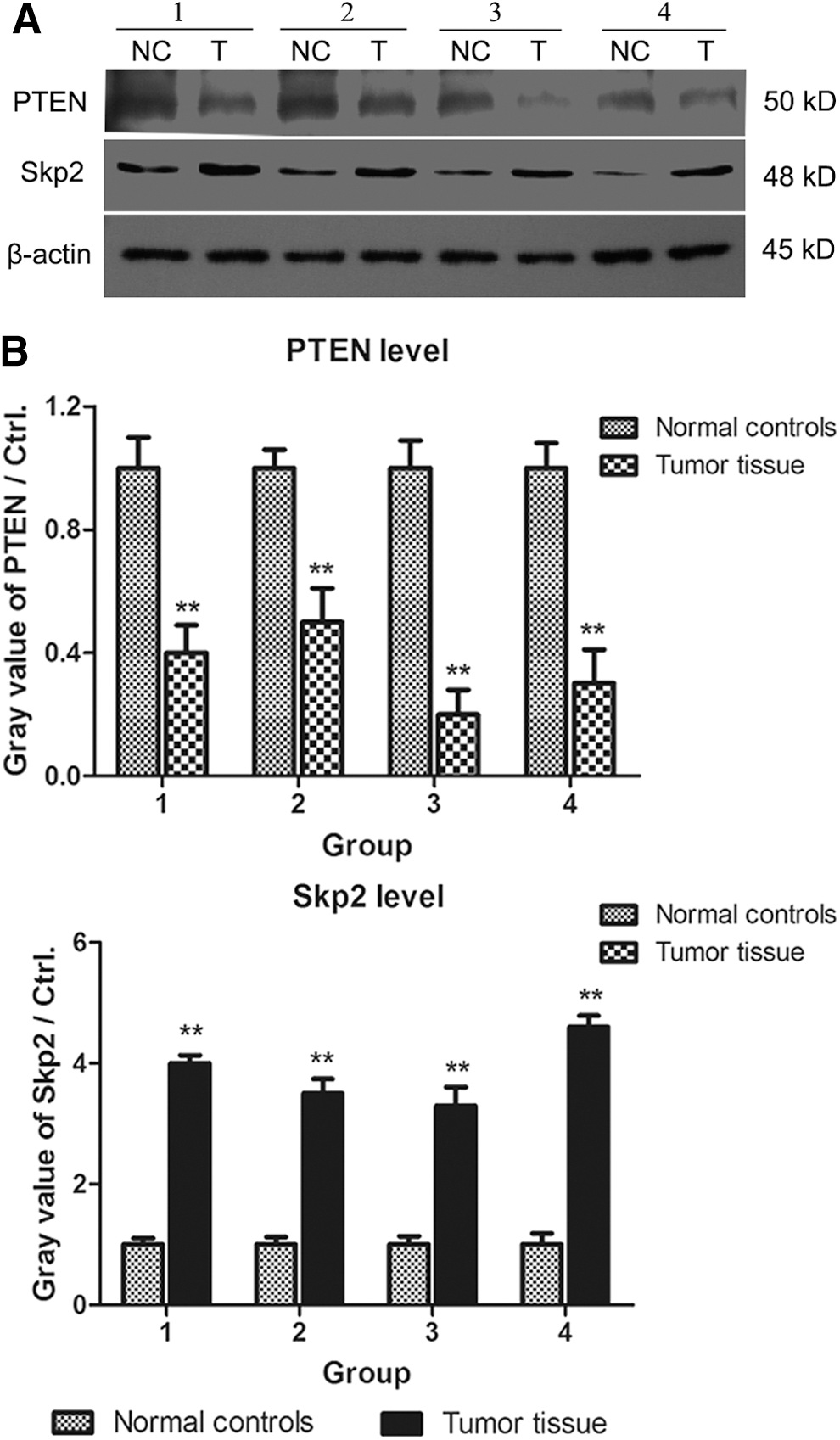
PTEN is negatively correlated with the expression of Skp2 in NSCLC tissues and normal control tissues.
miR-1297 regulates the expression of PTEN in A549 cells
To test whether the expression of PTEN was regulated by miRNAs, we used TargetScan to predict the related miRNAs, as shown in Figure 2A; miR-1297 could probably bind with the 3′UTR sequence of PTEN gene suggesting that hsa-miR-1297 might regulate the role of PTEN in the progression of lung cancer cells. Next, A549 cells were transfected with miR-1297 inhibitor and inhibitor negative control for 24 h and the western blot analysis result demonstrated that inhibition of miR-1297 increased the expression of PTEN in A549 cells. Consistently, transfection with miR-1297 mimic decreased the levels of PTEN in A549 cells. This was consistent with the results detected by immunofluorescence observation for PTEN (Fig. 2C).
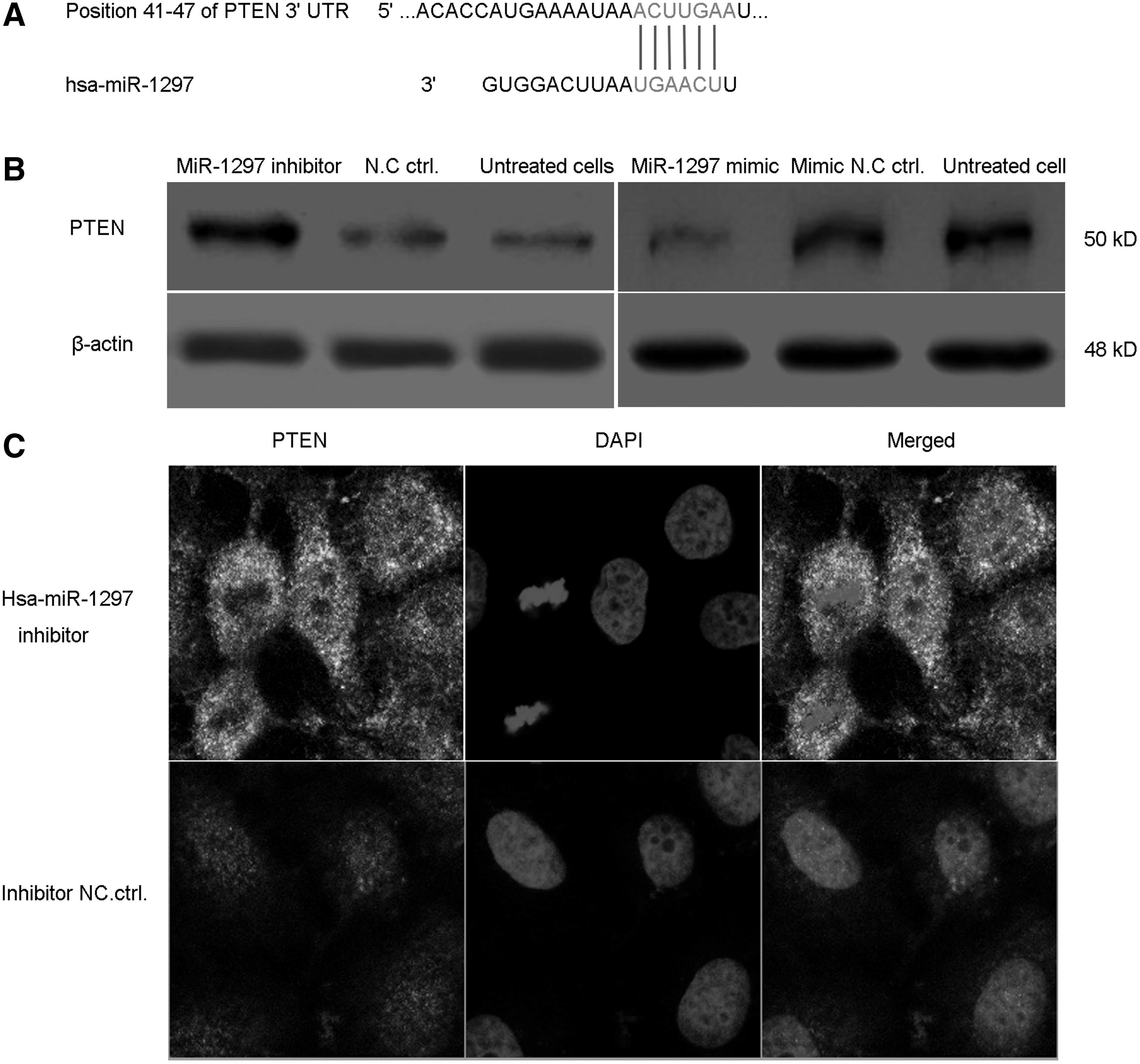
miR-1297 regulates the expression of PTEN in A549 cells.
Transfection with miR-1297 mimic decreases the expression of PTEN and increases the level of endogenous Skp2
To determine how miR-1297 regulated the expression of PTEN in lung cancer cell lines, A549 cells and NCI-H460 cells were transfected with miR-1297 mimic and mimic negative control for 48 h; PTEN and Skp2 levels were detected by western blot analysis. As shown in Figure 3A and B, the level of PTEN in A549 cells and NCI-H460 cells was significantly decreased in miR-1297 mimic-transfected cells compared with mimic negative control-transfected cells. Moreover, the decreased PTEN level was accompanied by increasing level of Skp2 in miR-1297 mimic-transfected cells, suggesting that lower expression of PTEN was associated with the higher level of Skp2 in lung cancer cell lines.
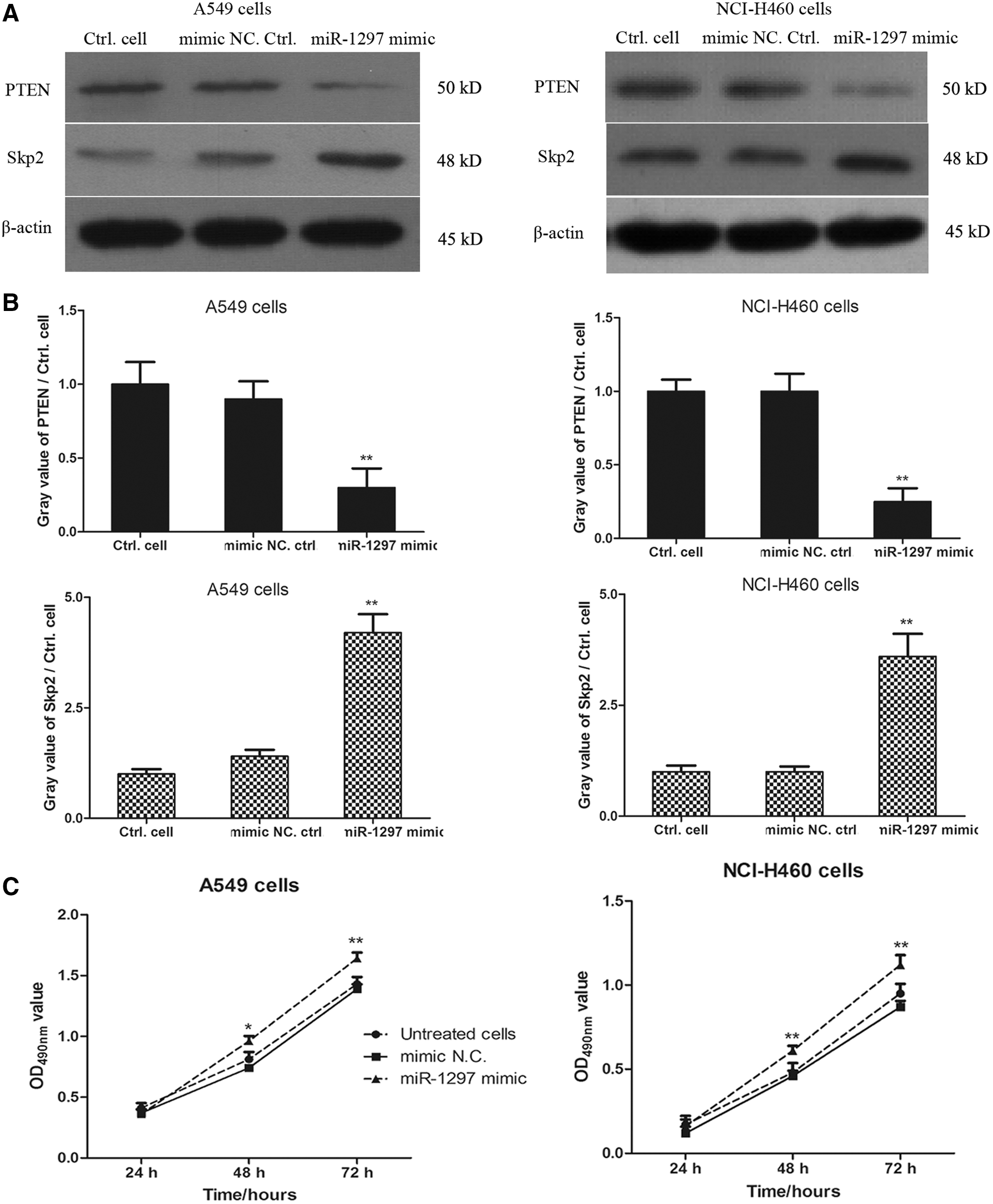
Transfection with miR-1297 mimic decreases the expression of PTEN and increases the level of endogenous Skp2.
Transfection with miR-1297 mimic increases cell viability of A549 cells and NCI-H460 cells
To test whether transfection with miR-1297 mimic affected the cell proliferation of lung cancer cells, MTT assay was performed to detect the cell viability in miR-1297 mimic-transfected cells and mimic negative control-transfected cells. As shown in Figure 3C, the data showed that miR-1297 mimic-transfected cells had higher proliferative ability compared with mimic negative control-transfected cells (*p < 0.05, **p < 0.01). There was no statistical difference between the untreated cells and mimic negative control-transfected cells.
Knockdown of Skp2 inhibits the proliferation of A549 cells but has no effects on the expression of PTEN
To identify the regulatory role between PTEN and Skp2, we further interfere with the expression of Skp2 in A549 cells and detected the levels of PTEN in Skp2 shRNA-transfected lung cancer cells. As shown in Figure 4A, interference with the expression of Skp2 in A549 cells did not affect the expression of PTEN. However, knockdown of Skp2 in lung cancer cells significantly inhibited cell proliferation of A549 after transfection of Skp2 shRNA for 48 and 72 h (Fig. 4D). All the results revealed that Skp2 might be regulated by PTEN in lung cancer cells, but the exact molecular mechanism need to be explored.
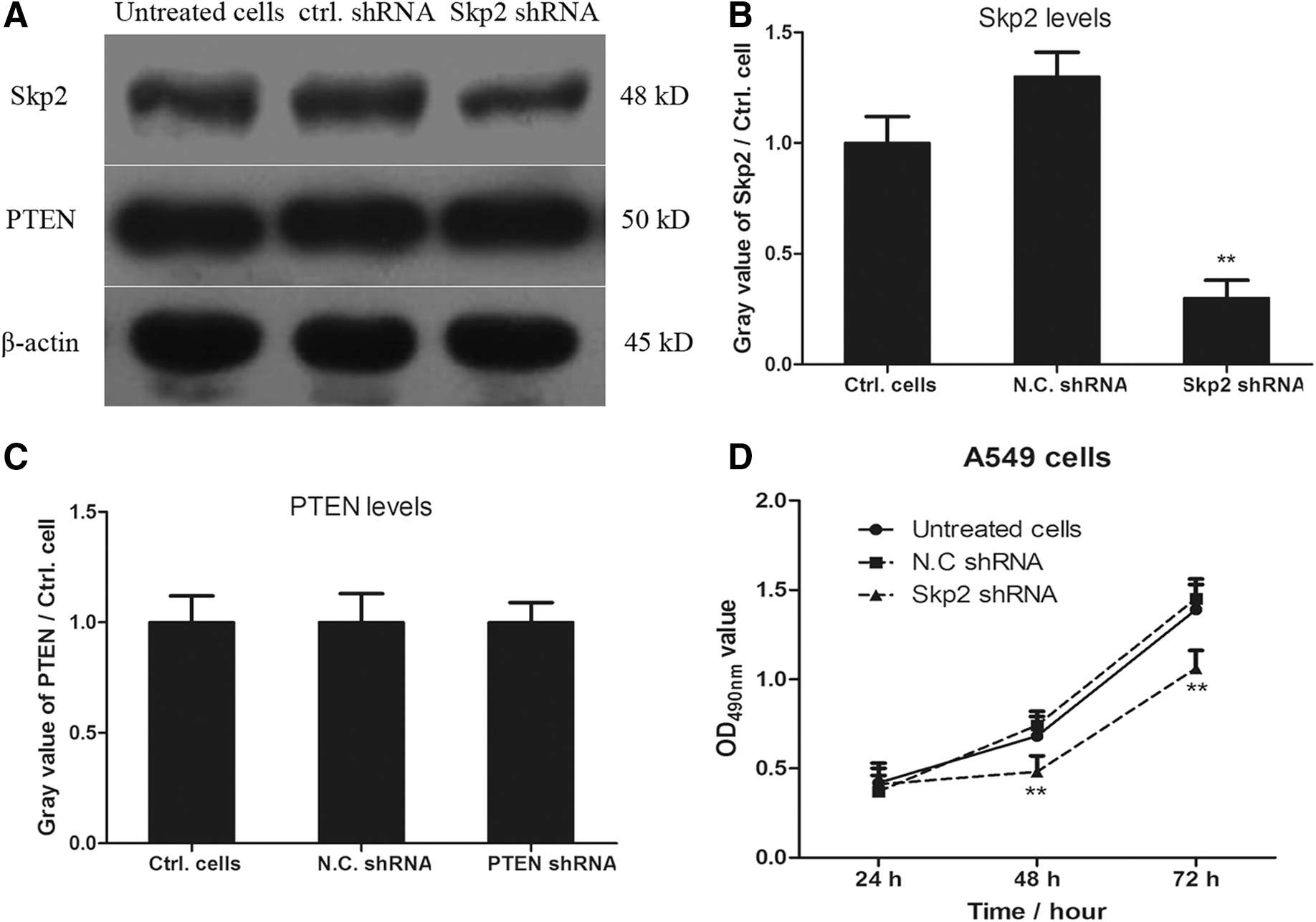
Knockdown of Skp2 inhibits the proliferation of A549 cells, but has no effects on the expression of PTEN. A549 cells were transfected with Skp2 shRNA and scrambled shRNA for 48 h. The expression of Skp2 and PTEN was detected by western blotting
miR-1297 inhibitor promotes the expression of PTEN and negatively regulates Skp2 involving in Akt-related signaling pathway
It has been reported that PTEN regulated a number of cellular processes, such as cell death and proliferation, through the phosphoinositide 3-kinase/protein kinase B (PI3K/AKT) signaling pathway (Wang et al., 2016; Xie et al., 2016). We wanted to detect whether PTEN regulated Skp2 through Akt-related signaling pathway. As shown in Figure 5A, overexpression of PTEN in A549 cells obviously decreased the levels of phosphorylated Akt in 48 h, as well as the levels of Skp2, while the total Akt level was not obviously changed. Moreover, A549 cells were transfected with miR-1297 inhibitor and negative control for 48 h; PTEN levels were obviously increased, while the levels of p-Akt and Skp2 were significantly decreased in miR-1297 inhibitor-transfected A549 cells (Fig. 5B). All the results demonstrated that PTEN negatively regulated the levels of Skp2 by decreasing the phosphorylation of Akt.
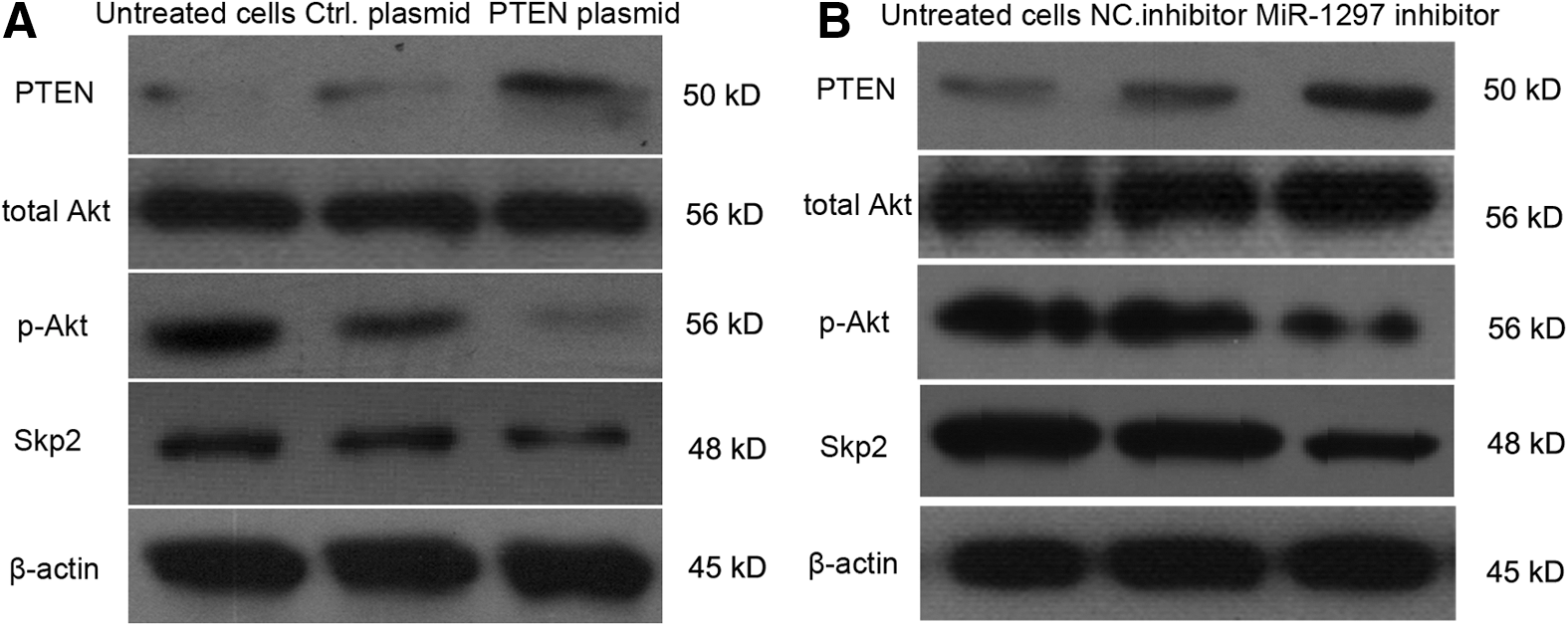
miR-1297 inhibitor promotes the expression of PTEN and negatively regulates Skp2 involving in Akt-related signaling pathway.
Discussion
In the present study, we found that the levels of PTEN were obviously decreased in NSCLC tissues compared with paired normal control tissues, as well as the NSCLC lines. Moreover, it also demonstrated that miR-1297 negatively regulated the expression of PTEN and positively regulated the expression levels of Skp2 by regulating the phosphorylation of Akt.
First, the endogenous PTEN was detected in human NSCLC specimens and paired normal tissue specimens by western blotting, and the results revealed that the levels of PTEN were significantly decreased in NSCLC specimens than that in controls of peritumoral tissues. Consistently, it has found that loss of PTEN is observed in clinical specimens of NSCLC, which is an independent poor prognostic factor for patients with NSCLC. Moreover, both p-Akt and PTEN are concerned with invasion and metastasis of NSCLC (Tang et al., 2006; Lim et al., 2007). Thus, it has been identified as loss of PTEN, and overexpression of Skp2 was observed in NSCLC patients and cell lines.
It is reported that PTEN exerts tumor suppressor functions in various human cancers, while Skp2 plays a role as the oncogene to promote tumorigenesis and progression (Deben et al., 2015; Van Nostrand et al., 2015). S-phase kinase-associated protein 2 is an enzyme involving in the ubiquitination and subsequent proteasomal degradation of target proteins in the process of cell cycle progression, signal transduction, and transcription (Hung et al., 2010). It is reported that Skp2 is overexpressed in various human cancers and induces cancer cell development and progression (Wei et al., 2013; Li et al., 2016; Zhang et al., 2016), which is associated with chemotherapeutic drug resistance in prostate cancer cells (Yang et al., 2016). Thus, Skp2 plays a role as an oncogene in the development and progression of human cancers, but little is known if or how the tumor suppressor PTEN regulates the oncogene Skp2 in the progression of human NSCLCs.
A549 and NCI-H460 cells were transfected with miR-1297 mimic for indicated time, and the data showed that transfection with miR-1297 mimic led to decrease of PTEN expression and upregulation of Skp2 in lung cancer cells, which promoted cell viability and cell survival. However, knockdown of Skp2 in A549 obviously inhibited cell proliferation of A549 cells suggesting that PTEN worked as a tumor suppressor gene, while Skp2 was an oncogene in the proliferation of lung cancer cells and there was a negative correlation on the expression of PTEN and Skp2 in lung cancer cells.
Next, the downstream target of PTEN was also to be investigated in the study. We have found that the expression levels of PTEN were conversely correlated with the levels of Skp2 in clinical NSCLC specimens and NSCLC cell line. Then, we tested whether the expression of Skp2 was regulated by PTEN. Luckily, miR-1297 mimic decreased the expression of PTEN and upregulation of Skp2 expression by decreasing levels of phosphorylated Akt. Thus, we speculated that miR-1297 mimic negatively regulated PTEN and positively regulated Skp2 through PIP3/Akt signaling pathway. Thus, PTEN and Skp2 might be the potential targets for the gene therapy for NSCLC patients.
Footnotes
Disclosure Statement
No competing financial interests exist.