
Abstract
Ovarian cancer (OC) is the most lethal gynecological cancer. The molecular mechanism of it is complicated, and numerous researches suggest that microRNAs are key regulators for it. This study was to investigate the pivotal role of miR-629 in the progression of OC and to reveal the possible molecular mechanism of its action. Testis-specific Y-like protein 5 (TSPYL5) is a tumor suppressor gene in various cancers, but there is little for its role in OC. OC OVCAR3 cells were transfected with the miR-629 vector, miR-629 inhibitor, and/or small interfering RNA (siRNA) targeting TSPYL5 (si-TSPYL5), respectively. After transfection, cell apoptosis, the ability of migration, and invasion were explored, as well as the level of miR-629 and TSPYL5 protein expression were detected by quantitative polymerase chain reaction and western blot. Compared with the control, there was increasing of miR-629, and decreasing of TSPYL5 and caspase 3 in OC tissue. Overexpression of miR-629 promoted the cell ability of migration and invasion and reduced OC cell apoptosis. In addition, elevated cancer inhibition ability of TSPYL5 induced by the miR-629 inhibitor was significantly blocked by inhibition of TSPYL5 (si-TSPYL5). All the above results suggested that miR-629 could promote OC proliferation, migration, and invasion by directly suppressing TSPYL5 expression, and inhibition of miR-629 might serve as a therapeutic target for OC.
Introduction
O
Testis-specific Y-like protein 5 (TSPYL5), encoding testis-specific Y-like protein, has been postulated to be a tumor suppressor gene in various cancers (Etcheverry et al., 2010; Taghavi et al., 2016; Witek et al., 2016; Lakshmanan et al., 2017). A study demonstrated that significant SNPs in TSPYL5 were associated with elevated plasma E2 in postmenopausal breast cancer patients, which prompted that TSPYL5 appeared to function as a transcription factor (Liu et al., 2013). However, when the function of TSPYL5 is alleviated by various reasons, such as hypermethylation (Zhai et al., 2009; Etcheverry et al., 2010; Qiu et al., 2016) or downregulation of multiple tumor biomarkers (Lakshmanan et al., 2017), in that case, it will promote tumor progression.
miRNAs are a series of small, noncoding RNAs with about 20–24 nucleotides in length that negatively regulate gene expression through binding to the 3′ untranslated region (UTR) of the target mRNA, leading to target mRNA degradation and/or posttranscriptional repression (Ono et al., 2011; Wang and Nanjing, 2011). Mounting evidence has demonstrated that miRNAs are frequently dysregulated in various cancers and act either as oncogenes or tumor suppressors depending on the cellular function of their target (Lu et al., 2005; Ling et al., 2013). In addition, increasing miRNAs has been reported to involve in the development and progression of OC (Zheng et al., 2015). miR-629 has been reported to serve as a tumor promoter and is upregulated in several cancer types, such as gastric cancer, lung cancer, renal cell carcinoma, and glioma (Yang et al., 2012; Jingushi et al., 2014b; Shin et al., 2015). However, there are few reports, which have demonstrated the function of miR-629 in the ovarian development, thus, this study is aimed to investigate the functional roles and the exact mechanisms of miR-629 involved in ovarian carcinogenesis.
In this study, we investigated the potential roles and related target genes of miR-629 in OC through a series of experiments in vitro and in vivo. Furthermore, we demonstrated that miR-629 directly targeted and downregulated TSPYL5 by binding its 3′UTRs. Studies in vitro showed that miR-629 significantly promoted the growth and metastasis of OC. All these data demonstrated that miR-629 plays a key role in the development and progression of OC and may be recognized as a useful therapeutic target for the treatment of OC.
Materials and Methods
Patients and samples
OC tissues and matched adjacent normal tissues were obtained from 20 patients with OC who had received surgery at the First Affiliated Hospital of Soochow University. All tissues were acquired with informed consent from patients, and the following tissue study had been approved by the First Affiliated Hospital of Soochow University. All histological diagnoses for OC and relative normal tissues were reviewed and recognized by three pathologists independently at the First Affiliated Hospital of Soochow University. For the analysis of gene expression, partial fresh samples were timely frozen and stored when obtained in liquid nitrogen until used. Besides, partial fresh samples were directly used for frozen slice and paraffin-embedded tissue sections.
Histology of OC and the expression level of proteins
Sample tissues were washed using phosphate-buffered saline (PBS) before fixation. Appropriately sized tissues were fixed in 4% paraformaldehyde overnight. For histological analysis, paraffin-embedded 5 mm tissue sections were cut and stained with Hematoxylin and Eosin. To detect the expression level of caspase 3, immunohistochemistry was carried out in sample tissues. They were fixed for about 1 h at 4°C immediately after dissection (T = 0) or 24 h (T = 24 h) after incubation in complete L-15 medium—in 4% paraformaldehyde, washed in PBS, permeabilized by a 24-h incubation at room temperature in 1% Triton X-100 in PBS, and preincubated for 8 h at room temperature in 1% Triton, 3% Normal Donkey Serum (NDS), and 1% ovalbumin in PBS. Samples were then incubated overnight at 4°C with specific rabbit polyclonal anti-human caspase 3 antibodies (ab203657; Abcam, Cambridge, English) diluted in a PBS solution (1:200) containing 1% BSA, 0.05% Triton, 1% NDS, and 1% ovalbumin. After three washes with PBS, samples were incubated 1 h at room temperature with anti-rabbit secondary antibody (Invitrogen, Carlsbad, CA) conjugated to Alexa Fluor 488 (1:2000) in a PBS solution containing 1% BSA, 0.05% Triton, 1% NDS, and 1% ovalbumin; the cell nuclei were counterstained for 10 min with Hoechst 33342 fluorescent dye (1: 1000; Invitrogen). Samples without incubation with primary antibody were used as negative control (NC). Digital images of stained tissues were obtained at 200 × magnification using Olympus BX51 and the Adobe photoshop 7.0 software.
Cell lines, cell culture, and transfection
OC cell lines OVCAR3, A2780, 3AO, SKOV3, and HRA were purchased from CCTCC (China Center for Type Culture Collection). All cell lines were cultured at 37°C under a 5% CO2 humidified atmosphere. Transfection was performed when cells were grown to 70% confluence, using the Lipofectamine 2000 (Invitrogen) according to the manufacturer's instructions.
RNA extraction and quantitative real-time PCR analyses
Total RNA was isolated from cell lines and tissue slides using TRIzol reagent (Invitrogen), respectively. For the detection of miR-629, RNA was reverse transcribed into cDNA using the All-in-One™ miRNA First-Strand cDNA Synthesis Kit (GeneCopoeia, Guangzhou, China), and then was used to perform quantitative real-time PCR (qRT-PCR) using the All-in-One miRNA qRT-PCR Detection Kit (GeneCopoeia). For the detection of mRNAs, RNA was reverse transcribed into cDNA using the GoScript Reverse Transcription (RT-PCR) System (Promega, Madison, WI), and the qRT-PCR for mRNAs was performed using GoTaq® qPCR Master Mix (Promega). All qRT-PCR was performed using ABI Prism 7500 system. U6 or GAPDH was used as an internal control for miRNA and mRNA quantification, respectively. The primer sequences for the qRT-PCR are listed in Table 1.
F, forward primer; R, reverse primer; RT, reverse transcription primer; TSPYL5, Testis-specific Y-like protein 5.
Cell proliferation, migration, invasion assay, and cell cycle assay
The cell apoptosis rate was detected by flow cytometry. Cells were seeded at 3 × 105 cells per well in six-well plates without FBS for 48 h. Then, cells were harvested and labeled with FITC-conjugated Annexin V and Propidium Iodide (BD Pharmingen) following the manufacturer's instructions. The apoptotic cells were measured by fluorescence-activated cell sorting (FACS). The migration and invasion ability of cells were detected by scratch test and transwell assay, respectively, as described in previous articles (Jingushi et al., 2014b). For the cell cycle analysis, cells were harvested by trypsinization, washed twice using cold PBS, and fixed in 70% ethanol overnight at 4°C. Then, cells were incubated with 20 μg/mL Propidium Iodide (LiankeBio, Zhejiang, China) for 20 min at room temperature, and cell cycle was analyzed with FACS flow cytometry (BD Biosciences, Franklin Lakes, NJ).
Western blot
A quantity of 30 μg of lysates per sample was separated by SDS-PAGE (sodium dodecyl sulfate–polyacrylamide gel electrophoresis) by 10% polyacrylamide gels and transferred onto polyvinylidene fluoride membrane. After blocking with 5% nonfat milk, the blots were incubated with the following primary antibodies: rabbit monoclonal TSPYL5 antibody (ab203657, 1:800 dilution), the secondary antibody was anti-rabbit (1:10,000). Then corresponding were immunodetected by incubation with horseradish peroxidase-linked related GAPDH antibody (KC-5G5, 1:1000 dilution; Kangchen Bio-Tech, Shanghai, China) as reference. Protein expression levels were detected using Image Lab software (Bio-Rad, CA).
Statistical analyses
Statistical analyses were carried out using SPSS, version 18.0 (SPSS, Inc., Chicago, IL). The difference among groups was determined by the two-tailed Student's t-test. p-Values <0.05 was considered as statistically significant.
Results
The pathological characteristics and expression of miR-629 and TSPYL5 in OC tissues
The pathological result showed that, compared with the adjacent normal tissues, there were lots of large cell nuclei in OC tissues (Fig. 1A). Moreover, the expression of caspase 3 was significantly repressed in OC tissues (Fig. 1B). Then our study detected miR-629 level in 20 clinical OC tissues (Table 2). To verify the high expression of miR-629 in OC patients, miR-629 expression of each OC tissue paired with adjacent normal tissues were detected by qRT-PCR. The result showed that miR-629 was significantly overexpressed in OC tissues (Fig. 1D). At the same time, downregulation of TSPYL5 was also confirmed in these clinical OC tissues (Fig. 1C, E). Taken together, these results strongly prompt us that miR-629 and TSPYL5 may be correlated with the progress of OC.
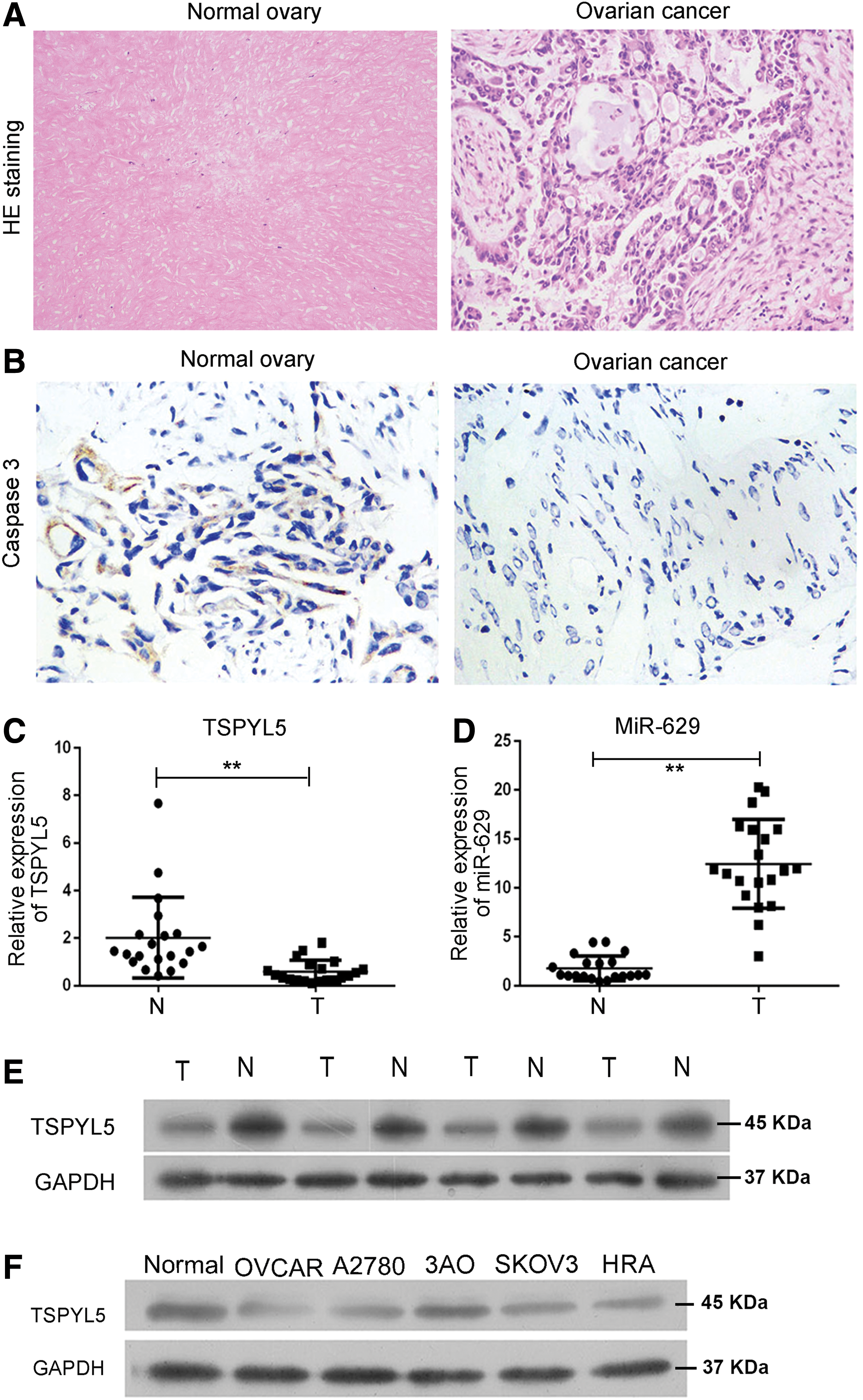
The pathological characteristics and expression of miR-629 and TSPYL5 in OC tissues and adjacent normal tissues.
TSPYL5 was downregulated in OC cell
To further explore the expression level of miR-629 in vitro, we next carried out studies in five OC cell lines (SKOV3, A2780, 3AO, OVCAR3, and HRA) and finally chose one of the cell lines (OVCAR3), in which miR-629 was effectively upregulated for the use in the continuing in vitro experiment. In agreement with the observations in vivo, we found that TSPYL5 was downregulated compared with the normal ovarian cells 0088 (Fig. 1F). This result consists of the caspase 3 expression in OC tissues for the function of caspase 3 (Fig. 1B).
The protective function of TSPYL5 in OC cell lines
TSPYL5 was repeatedly reported as tumor inhibitor in various cancers, and to confirm its role in OC, we examined the function of TSPYL5 after overexpression of it in OVCAR3 cells. Following this, the cell function experiments were carried out (Fig. 2). The results showed that TSPYL5 played a key role in inhibiting the progress of OC, and overexpression of TSPYL5 could improve OVCAR3 cell apoptosis (p = 0.022; Fig. 2A); the result of flow cytometry showed that TSPYL5 inhibited the cell proliferation actively and shortened the S phase of cell cycle, and increased the ratio of G0/G1 obviously (p = 0.022; Fig. 2B). TSPYL5 also repressed their migration and invasion (Fig. 2C, D). Compared with the NC, the rate of cell apoptosis was significantly increased (p < 0.01, Fig. 2A), whereas the ability of migration and invasion were effectively reduced (p < 0.01, Fig. 2C–E). All these results showed that TSPYL5, as a tumor inhibitor, would also exert the same function in OC.

TSPYL5 inhibits the characteristics of OC cell lines in vitro. When compared with the NC
miR-629 promotes OC cell proliferation, migration, and invasion in vitro
Having confirmed the association of miR-629 expression with OC patients, we carried out functional experiment of miR-629 on OVCAR3 cells. We enhanced or repressed the function of miR-629 in OVCAR3 cells through infecting with miR-629 mimic or miR-629 inhibitor, using the empty vector (Mock) as control. We then used the flow cytometry assay to assess the effects of miR-629 on cell apoptosis, and the result showed that miR-629 overexpression significantly decreased the apoptosis rates of OVCAR3 cells (Fig. 3A, E), and there was a significant difference between the mimic group and the mock group. The inhibiting effect of miR-629 on cell apoptosis and proliferation was further confirmed by cell cycle assay (Fig. 3B). Next, we explored the possible effect of miR-629 on cell migration and invasion. The scratch test demonstrated that overexpressing miR-629 promoted cell migration effectively compared with mock group cells, whereas the cells transfected with miR-629 inhibition had the widest cell scratch (Fig. 3C). At the same time, the transwell assay indicated that overexpression of miR-629 significantly promoted the invasion of OVCAR3 cells (Fig. 3D), and there was a significant difference between the mimic group and the mock group (Fig. 3F). All these results clearly demonstrated that miR-629 promoted OC cell proliferation, migration, and invasion.

miR-629 promoted OC cell proliferation, migration, and invasion in OVCAR3 cells.
TSPYL5 is a direct target of miR-629 in OC
Having demonstrated the biological function of miR-629 on TSPYL5 in OC cells, we therefore hypothesized that TSPYL5 may be a target gene of miR-629. Thus, we used the miRWALK (
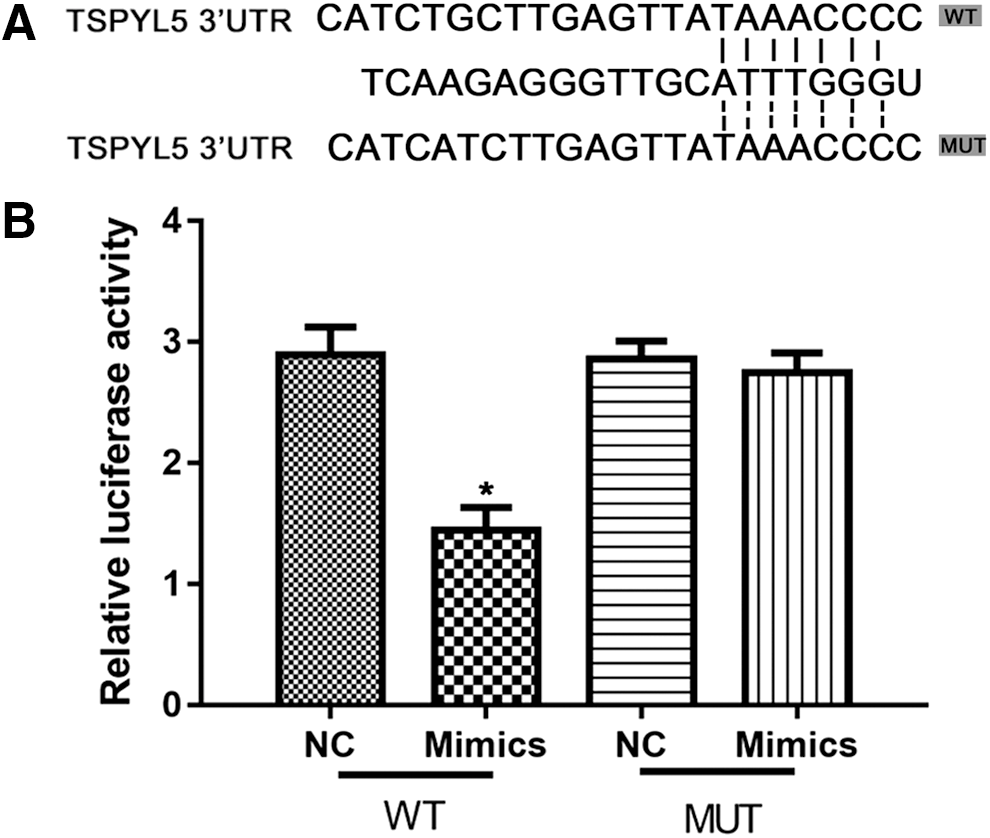
TSPYL5 is a direct target of miR-629 in OVCAR3 cells.
The function of TSPYL5 on OVCAR3 cells was regulated by miR-629 in the cell itself
To further explore whether the function of TSPYL5 on OVCAR3 cells was regulated by miR-629 in the cell itself, we did experiment to compare the different functions of TSPYL5 on OVCAR3 cells with or without miR-629 in the cell itself through transfecting the cell with NC or inhibitor; the result showed that compared with the NC group, the progress of the OC was inhibited by transfection of miR-629. As shown in Figure 5, cell apoptosis decreased posttreatment with miR-629 inhibitor (Fig. 5A), G0/G1 phase was enhanced (Fig. 5B), and cell mobility (Fig. 5C, D) was elevated after cells were transfected with miR-629 inhibitor. However, apoptosis, cell cycle, migration, and invasion could be blocked significantly by cotransfection with TSPYL5 siRNA (small interfering RNA) (Fig. 5), which reflected that the tumor inhibition function was largely exerted by TSPYL5. In addition, the OVCAR3 cells in the blank group was the most active due to the ability of migration and invasion, which demonstrated that in this OVCAR3 cells, the tumor inhibition function is usually inhibited by miR-629 (Fig. 5).
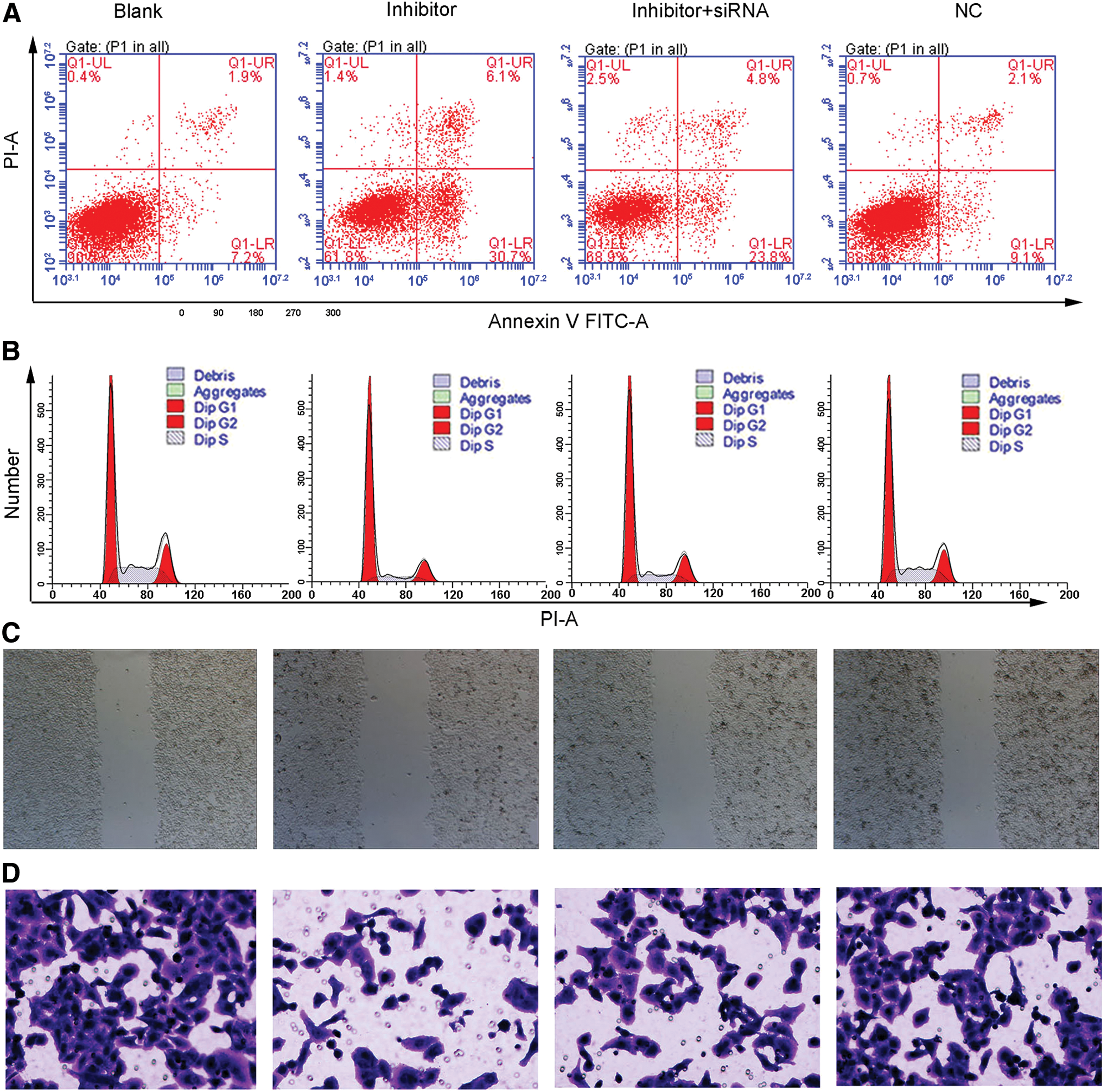
miR-629 suppresses OC oncogenesis through TSPYL5 in vitro. The OVCAR3 cells were transfected with miR-629 inhibitor and/or siRNA-TSPYL5. Then the progress of the OC (including apoptosis, cell cycle, migration, and invasion) was determined. The
Discussion
OC is the most common gynecological cancer with a high mortality rate and low 5-year survival rate, which threatens the health of many women. Pathological mechanism and pathogen have been reported to be complex for it (Gilks and Prat, 2009; Karst and Drapkin, 2010). Series of miRNAs have been verified to act as oncogenes or tumor suppressor genes depending on the regulatory function of their targets (Yuan et al., 2015). Emerging data have indicated that miR-629 involved in the development of different cancers (Jingushi et al., 2014a, b). However, there were few reports focusing on the function of miR-383 on ovarian carcinogenesis.
In previous three studies, miR-629 was found to be involved in the development of cancers through a miRNA-feedback loop way, for example, miR-629-HNF4α-miR-124 inflammatory feedback loop circuit in liver cancer (Hatziapostolou et al., 2011; Cao et al., 2013; Gougelet and Colnot, 2013). In the present study, we further validated the expression and function in OC, and the results showed that miR-629 was significantly upregulated in most of OC specimens compared with the adjacent nontumor tissue. Moreover, we found that overexpression of miR-629 significantly promoted proliferation and invasion in OVCAR3 cells, meanwhile repressed the expression of TSPYL5, whereas inhibition of miR-629 decreased proliferation and invasion of OVCAR3 cells and increased the expression and function of TSPYL5. These findings together suggested that miR-629 may play an important role in promoting proliferation and invasion of OC through targeting TSPYL5.
However, the specific regulation mechanism underlying how miR-629 is involved in OC was unknown, therefore, we predicted potential targets of miR-629, and TSPYL5 was selected for further study. TSPYL5 is a key mediator for multiple cancers, such as TSPYL5 gene expressions, and are potentially correlated with patient survival in all endometrial cancer grades (Witek et al., 2016); MUC16 regulates TSPYL5 for lung cancer cell growth and chemoresistance by suppressing p53 (Lakshmanan et al., 2017). TSPYL5 plays a key role in live cancer through inflammatory feedback loop circuit (Qiu et al., 2016). This study found that overexpression of miR-629 decreased TSPYL5 in both mRNA and protein levels in OC cells. Furthermore, luciferase reporter assay demonstrated that miR-629 reduced the luciferase reporter activity of wild-type 3′UTR, but not mutant 3′UTR of TSPYL5.
Conclusion
All the above results prompted us that miR-629 may play an important role in OC progression as a novel oncogenic miRNA in both in vitro and in vivo studies. miR-629 was upregulated in OC and overexpression of miR-629 promoted OC cell proliferation and metastasis in both in vitro and in vivo. The function of miR-629 in OC may be exerted through downregulation of the target gene TSPYL5 directly.
There is some deficiency in this study, to our knowledge, to verify the function of miR-629 on OC tumorigenicity, and it is better to carry out an in vivo experiment in an animal model. Therefore, in future studies, the tumor-promotive role of miR-629 in vivo needs to be performed.
Footnotes
Disclosure Statement
No competing financial interests exist.