
Abstract
The Editor-in-Chief of DNA and Cell Biology officially retracts the article entitled, Downregulation of miR-335-5p by Long Noncoding RNA ZEB1-AS1 in Gastric Cancer Promotes Tumor Proliferation and Invasion by Li-li Zhang, Lan-fang Zhang, Xiao-he Guo, De-zhong Zhang, Fang Yang, and Ying-ying Fan (DNA Cell Biol. (2018;37(1):46–52; doi: 10.1089/dna.2017.3926).
A post on the PubPeer platform1 raised concerns about the possibility of duplicated images within Figure 4. An independent analysis found that Figure 4D exhibits a matching section across two other images that are identified in the paper as representing different experimental conditions. Additionally, the analysis identified Figure 6G as a composite image. Other features of the text have raised concerns about the originality and authorship of this work.
The Editor-in-Chief of DNA and Cell Biology contacted all authors listed on the paper, asking them to provide original data and uncropped images for Figure 4, as well as an explanation for the cause for the evident composite image in 6G. No communication or other statements were received from any of the listed authors.
Because of the doubts raised with regards to the originality and reliability of the published work, the Editor-in-Chief fully retracts the published article.
The authors were notified of the decision to retract the paper, but no acknowledgment was received.
DNA and Cell Biology
is committed to upholding the rigors of scientific publishing and the veracity of the literature.
1. Actinopolyspora biskrensis.
Introduction
G
Long noncoding RNAs (lncRNAs) are nonprotein coding transcripts longer than 200 nucleotides (Mercer et al., 2009). Increasing evidence showed that lncRNA is an important class of pervasive genes involved in carcinogenesis and metastasis and plays multiple roles in regulating gene expression at the transcriptional, posttranscriptional, and epigenetic levels (Shi et al., 2013; Fatica and Bozzoni, 2014). For example, reduced LOC285194 in pancreatic ductal adenocarcinoma tissues was correlated with tumor progression and might be a potential molecular biomarker for predicting the prognosis of patients (Ding et al., 2014). lncRNA ANRIL indicated a poor prognosis of cervical cancer and promoted carcinogenesis through the PI3K/Akt pathway (Zhang et al., 2017a). lncRNA UCA1 promoted cell progression by acting as a competing endogenous RNA of ATF2 in prostate cancer (Zhang et al., 2017b).
lncRNA ZEB1 antisense1 (ZEB1-AS1) was an antisense transcript which originated from the promoters of ZEB1. Study showed that ZEB1-AS1 could positively regulate the ZEB1 expression and promote cell growth and metastasis in hepatocellular carcinoma (Li et al., 2016). lncRNA ZEB1-AS1 could act as an oncogene in osteosarcoma by epigenetically activating ZEB1 (Liu and Lin, 2016). lncRNA ZEB1-AS1 could contribute to STAT3 activation by associating with IL-11 in B-lymphoblastic leukemia (Wang et al., 2017). However, whether ZEB1-AS1 is involved in the development of GC and its potential role in the regulation of GC progression remain unclear.
In the present study, we explored the expression and function of ZEB1-AS1 in GC. Results showed that ZEB1-AS1 expression was significantly upregulated and associated with advanced clinical features and poor prognosis in GC patients. ZEB1-AS1 inhibition suppressed GC cell proliferation and invasion both in vitro and in vivo. Furthermore, our data showed that ZEB1-AS1 was involved in the progression of GC by regulating miR-335-5p expression.
Materials and Methods
Sample collection
Seventy-six GC tissues and adjacent nontumor tissues were collected in The First Affiliated Hospital of Xinxiang Medical University. Tissue samples were frozen in liquid nitrogen immediately after dissection and then stored at −80°C. This study was approved by the Ethics Committee of The First Affiliated Hospital of Xinxiang Medical University. Informed consent was obtained from all individual participants included in the study. All the patients did not receive any treatment before the operation.
Cell culture and transfection
Human GC cell lines (SGC7901, HGC27, BGC823, MKN45, MKN28, and AGS) and the normal gastric epithelial cell line GES-1 were obtained from the type Culture Collection of Chinese Academy of Sciences (Shanghai, China). Cells were maintained at 37°C in a humidified incubator containing 5% CO2 in RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS) and 100 U/mL penicillin and 100 μg/mL streptomycin.
For analysis of ZEB1-AS1, specific siRNAs against ZEB1-AS1 (si-ZEB1-AS1) and a scrambled negative control (si-NC) were synthesized by RiboBio Co., Ltd. (Guangzhou, China). For analysis of ZEB1-AS1 related miRNA, miR-335-5p mimics, miR-335-5p inhibitors, and si-NC were synthesized by RiboBio Co., Ltd. For transfection, Lipofectamine™2000 (Invitrogen) was used as the transfection reagent according to the manufacturer's instructions.
Quantitative real-time PCR
Total RNA was isolated from tumor tissues and cell lines using TRIzol reagent (Invitrogen). The purity of RNA was examined by spectrophotometry, and the first strand cDNA was synthesized using reverse transcription reagents (ABI) or the TaqMan MicroRNA Reverse Transcription Kit (ABI) following the manufacturer's instructions. Quantitative real-time PCR (qRT-PCR) was performed using SYBR Select Master Mix for CFX (Invitrogen) and using the CFX Connect™ Real-time PCR system (Bio-Rad). All results were normalized to the expression of GAPDH or snRNA U6. The quantitative analysis was calculated using 2−ΔΔCt method.
Cell viability assay
Cell viability was determined using a Cell Counting Kit (CCK-8; Beyotime) according to the manufacturer's protocol. Briefly, the transfected cells (2 × 105 cells/mL) were seeded into 96-well plate and then 10 μL CCK-8 solution was added to each well of the plate, and the plates were cultured for 2 h. The absorbance of each well at a wavelength of 450 nm was measured using a multi-detection microplate reader.
Cell invasion assay
Cell invasion was detected using transwell chambers (8 μm pore size; Millipore) coated with Matrigel (BD). The transfection cells (5 × 104) suspending in 150 μL serum-free RPMI-1640 medium were added into the upper chamber, and the bottom compartment of the chamber was filled up with 500 μL RPMI-1640 medium with 20% FBS as a chemoattractant. After incubation for 48 h, the noninvading cells inside the upper chamber were scraped off by a cotton swab. The invaded cells on the lower chamber were then fixed with 4% formaldehyde and stained with 0.1% crystal violet. The cells were counted under a light microscope.
Tumor formation assay in nude mice
Four-week-old male nude mice were purchased from the National Laboratory Animal Center (Shanghai, China) and divided into two groups for subcutaneous injection using cells transfected with sh-NC or sh-ZEB1-AS1. Animal tumors were collected for volume measuring every week and sacrificed 7 weeks after injection. The tumor volume was calculated by the formula: volume (mm3) = length × width2/2. All experiments were performed strictly in accordance with a protocol approved by the Administrative Panel on Laboratory Animal Care.
Luciferase reporter assays
A web-based program known as DIANA Tools program (
RNA-binding protein immunoprecipitation
RIP experiments were performed using the Magna RIP RNA-Binding Protein Immunoprecipitation Kit (Millipore) and the Ago2 antibody (Abcam) following the manufacturer's protocol. Coprecipitated RNAs were subjected to qRT-PCR analysis.
Statistical analysis
Data were presented as mean ± SD. Data were analyzed by Student's t-test or one-way ANOVA using SPSS 18.0 software. A p-value less than 0.05 was considered to be statistically significant.
Results
ZEB1-AS1 was upregulated in GC
In the present study, we determined ZEB1-AS1 expression in 76 GC patients and analyzed the relationship between ZEB1-AS1 expression and clinicopathological features. Results showed that ZEB1-AS1 was significantly upregulated in GC tissues compared with adjacent nontumor tissues (Fig. 1A; p < 0.05). Then, the expression of ZEB1-AS1 in GC tissues was classified into low ZEB1-AS1 expression and high ZEB1-AS1 expression group according to the median value of ZEB1-AS1 expression. Results showed that high ZEB1-AS1 expression was associated with advanced TNM stage and lymph node metastasis (Table 1; p < 0.05), but was not significantly correlated with gender, age, tumor size, or differentiation (Table 1; p > 0.05). ZEB1-AS1 expression was clearly increased in patients with III+IV TNM stage compared with that in the patients with I+II TNM stage (Fig. 1B; p < 0.05), in patients showing lymph node metastasis compared with that in the patients not showing lymph node metastasis (Fig. 1C; p < 0.05). In addition, Kaplan–Meier analysis showed that GC patients with high ZEB1-AS1 expression have a poor overall survival (Fig. 1D; p < 0.05). To compare our data to other available expression data for ZEB1-AS1, the Kaplan–Meier plotter database (
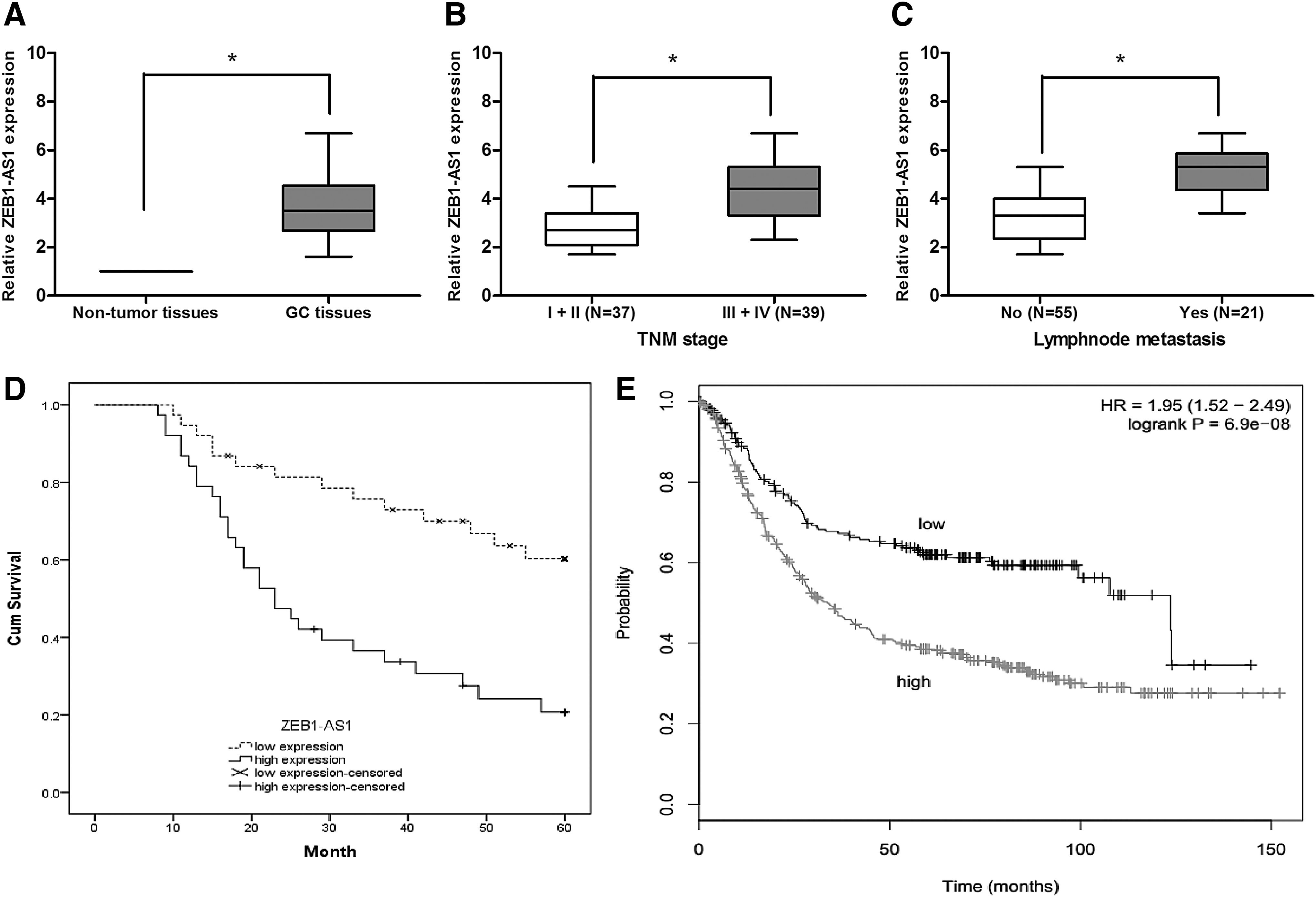
lncRNA ZEB1-AS1 was upregulated in GC.
Knockdown of ZEB1-AS1 inhibited GC cell proliferation and invasion
To explore the oncogenic roles of ZEB1-AS1 on GC, ZEB1-AS1 expression was detected by qRT-PCR in GC cell lines (SGC7901, HGC27, BGC823, MKN45, MKN28, and AGS) and the normal gastric epithelial cell line GES-1. Results showed that ZEB1-AS1 expression was upregulated in GC cell lines compared with GES-1 cell line (Fig. 2A; p < 0.05). According to ZEB1-AS1 expression level in GC cells, we transfected with si-ZEB1-AS1 or si-NC into BGC823 and MKN28 cells, and the knockdown efficiency was confirmed by qRT-PCR (Fig. 2B; p < 0.05). Cell proliferation was subsequently measured by CCK-8 assay. Results showed that ZEB1-AS1 inhibition significantly suppressed BGC823 and MKN28 cell viability compared to si-NC group (Fig. 2C, D; p < 0.05). In addition, transwell invasion assay showed that knockdown of ZEB1-AS1 significantly decreased the invasion abilities of BGC823 and MKN28 cells (Fig. 2E; p < 0.05).
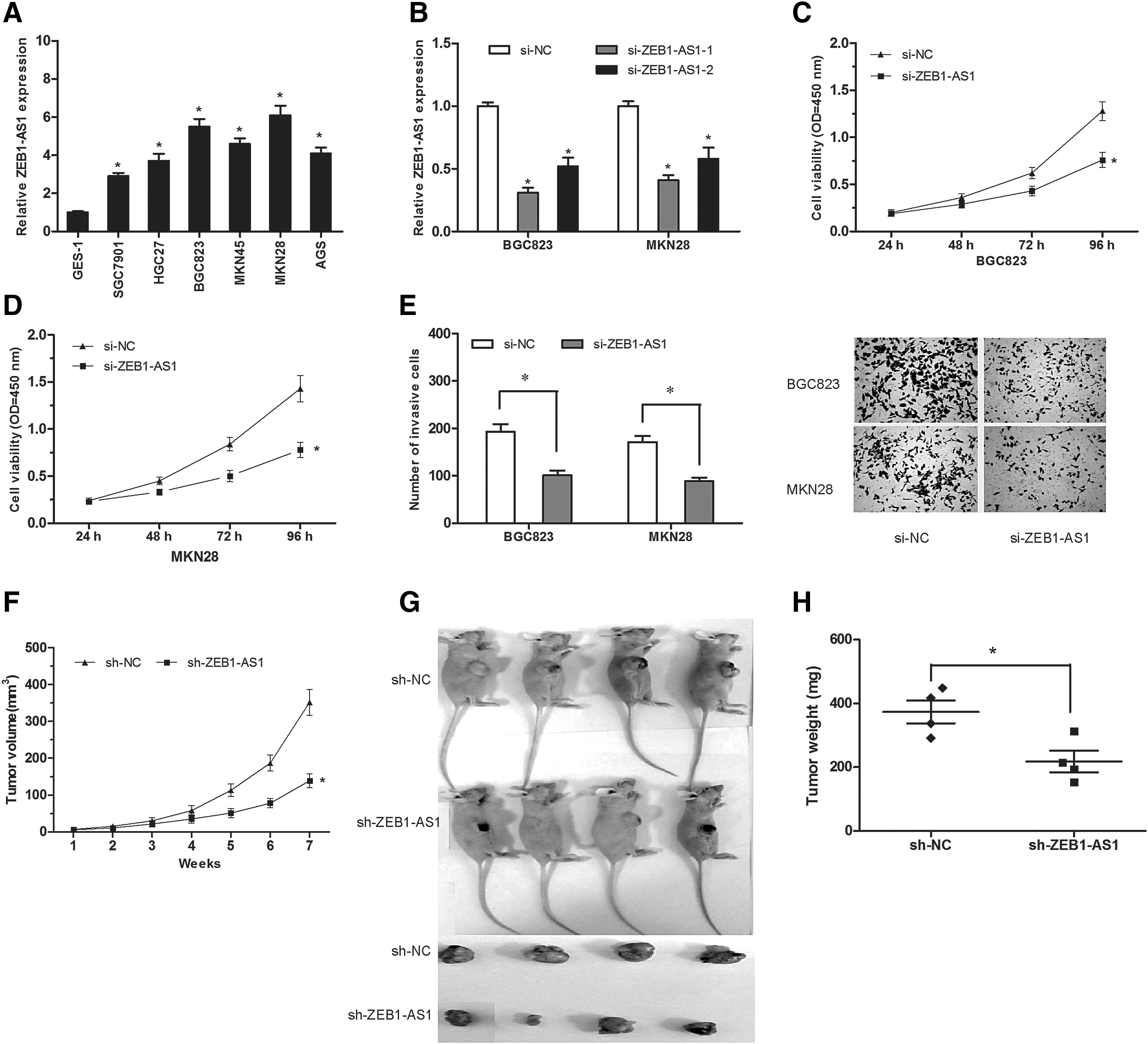
lncRNA ZEB1-AS1 suppression inhibited GC cell proliferation and invasion.
To confirm the biological function of ZEB1-AS1 in GC cells in vivo, BGC823 cells stably transfected with sh-ZEB1-AS1 or sh-NC were subcutaneously injected into the nude mice. Tumor volume was determined every week. Results showed that the tumor growth was significantly reduced after ZEB1-AS1 was knocked down (Fig. 2F; p < 0.05). Subcutaneous tumors were removed subsequently and weighed 7 weeks after implantation. Results showed that tumor size and weight in sh-ZEB1-AS1 transfected group were less than that in sh-NC group (Fig. 2G, H; p < 0.05).
miR-335-5p expression was directly regulated by ZEB1-AS1
Increasing evidence has shown that lncRNA might serve as a ceRNA or molecular sponge to regulate the expression and function of miRNA (Salmena et al., 2011; Tay et al., 2014). In the present study, the bioinformatics tool DIANA was used to predict target miRNA regulated by ZEB1-AS1 and miR-335-5p selected as the potential target (Fig. 3A). To support this hypothesis, we detected miR-335-5p expression in GC tissues. We found that miR-335-5p was significantly decreased in GC tissues compared with adjacent nontumor tissues (Fig. 3B; p < 0.05). Pearson's correlation analysis showed an inverse correlation between miR-335-5p and ZEB1-AS1 in GC tissues (Fig. 3C; p < 0.05). We further determined miR-335-5p expression in ZEB1-AS1 inhibited GC cells and found that miR-335-5p was boosted after ZEB1-AS1 downregulation (Fig. 3D; p < 0.05), while ectopic overexpression of miR-335-5p did not affect ZEB1-AS1 expression (Fig. 3E; p > 0.05). Furthermore, luciferase reporter assay indicated that miR-335-5p overexpression reduced the luciferase activity of ZEB1-AS1-Wt, but had no inhibitory effect on ZEB1-AS1-Mut (Fig. 3F; p < 0.05). To more extensively analyze the mechanism that might be involved in the reciprocal relationship, RNA immunoprecipitation experiments were taken using antibody against Ago2, a key component of RISC complex. We found that both miR-335-5p and ZEB1-AS1 were in the Ago2-pulled down pellet (Fig. 3G; p < 0.05). Collectively, these findings indicated that ZEB1-AS1 directly interacted with miR-335-5p and negatively regulated miR-335-5p expression in GC cells.
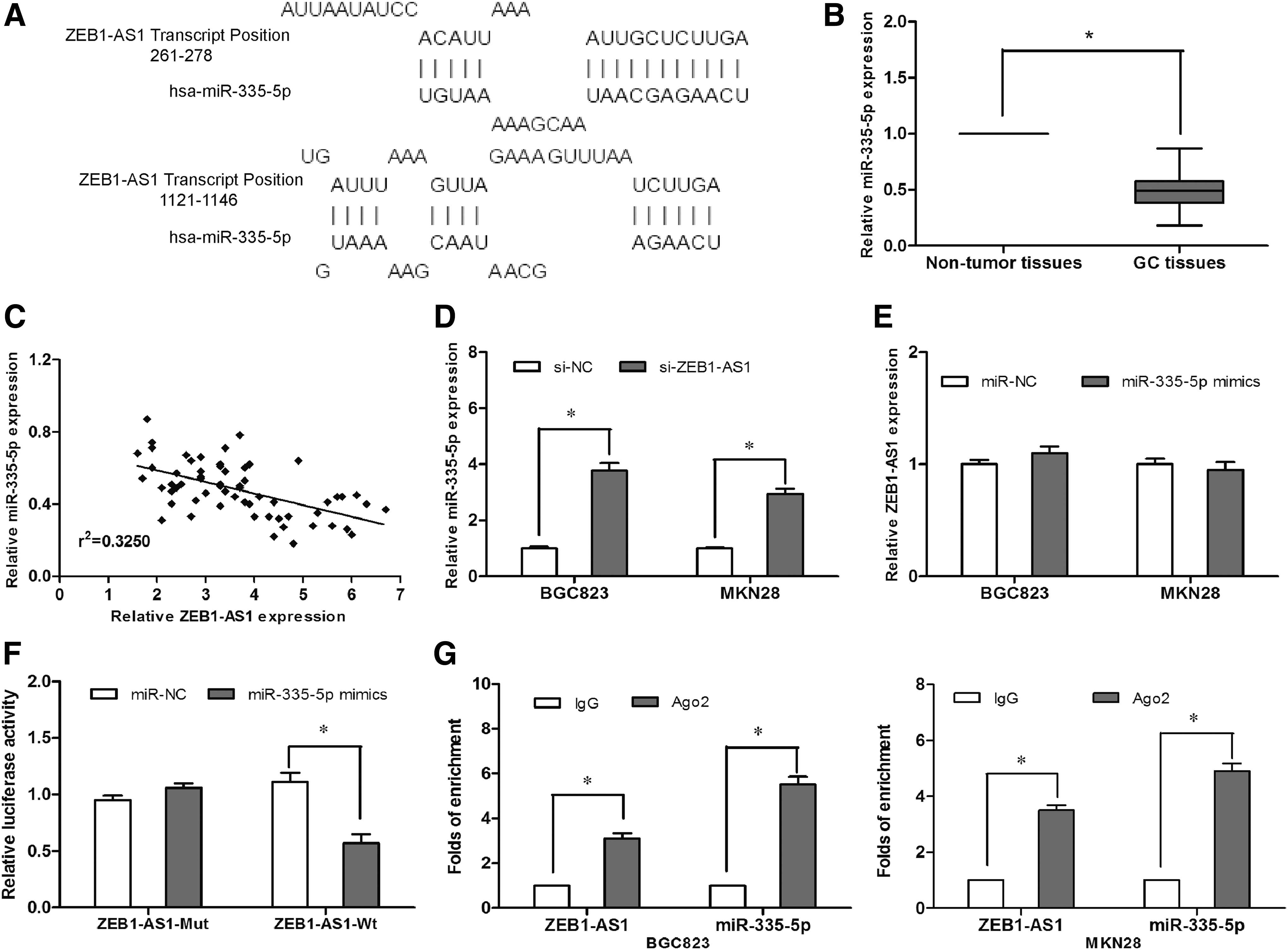
miR-335-5p expression was directly regulated by ZEB1-AS1.
ZEB1-AS1 promoted GC cell proliferation and invasion through miR-335-5p
To determine whether ZEB1-AS1 exerted biological functions through miR-335-5p, we performed rescue experiments by suppressing miR-335-5p expression in ZEB1-AS1 knockdown cells (Fig. 4A; p < 0.05). CCK-8 assay revealed that knockdown of ZEB1-AS1 decreased cell proliferation and then miR-335-5p inhibitors blocked this decrease mediated by ZEB1-AS1 (Fig. 4B; p < 0.05). Meanwhile, we also found that knockdown of ZEB1-AS1 decreased cell invasion, and miR-335-5p inhibitors blocked this decrease mediated by ZEB1-AS1 (Fig. 4C; p < 0.05). Taken together, these results demonstrated that ZEB1-AS1 regulated GC cell proliferation and invasion at least partially through miR-335-5p.
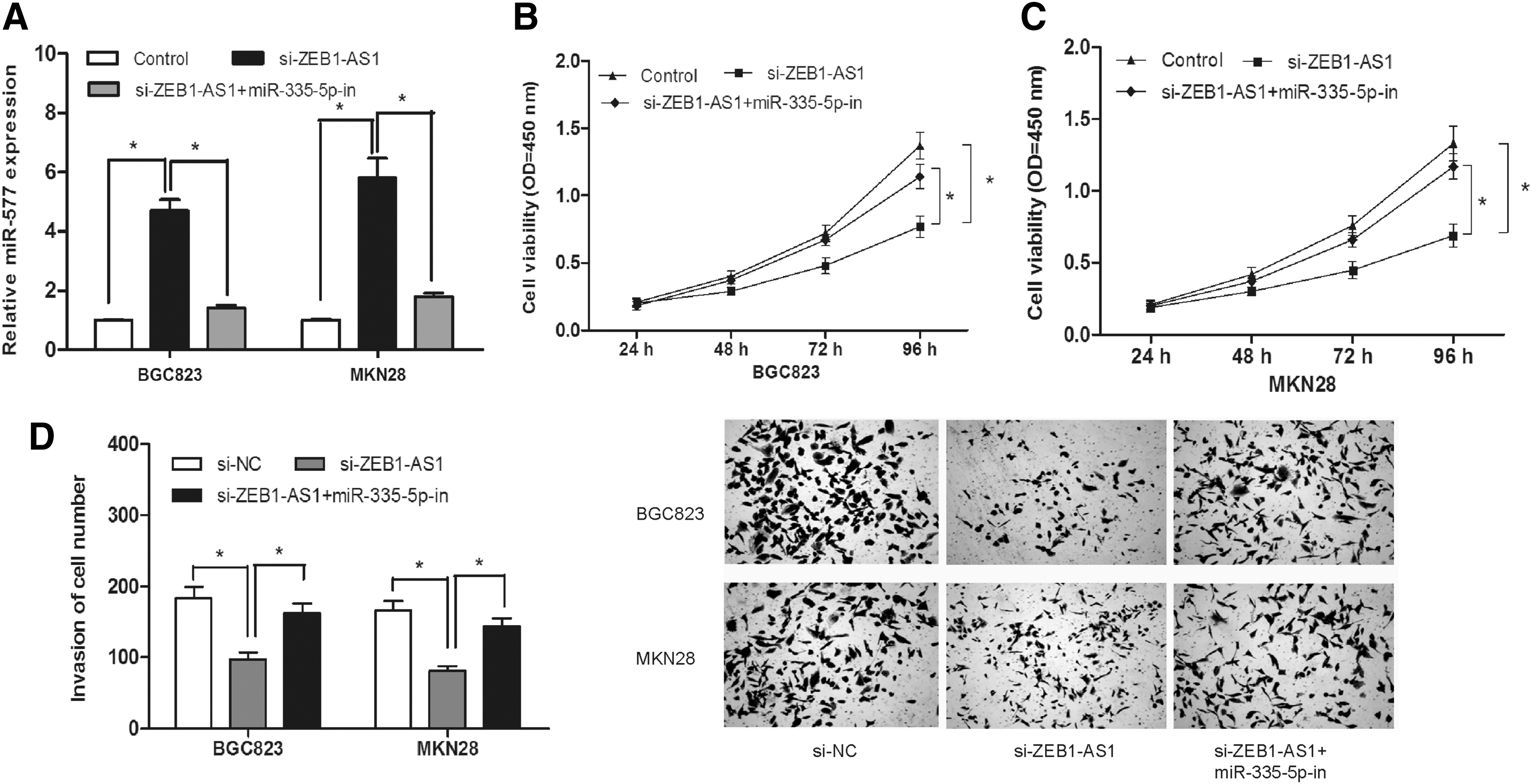
ZEB1-AS1 promoted GC cell proliferation and invasion through miR-335-5p.
Discussion
GC is one of the most deadly malignancies; the prognosis is poor because lots of GC patients are diagnosed at a later stage when treatment is not effective (Japanese Gastric Cancer Association, 2011). Therefore, the identification of novel prognostic biomarkers and therapeutic targets may have an enormous potential to improve the outcomes of GC. Multiple evidences illustrate that aberrant lncRNA expression is involved in GC progression, and applying lncRNA as a novel diagnostic and therapeutic tool has obtained more and more attention (Sánchez et al., 2013). For example, decreased expression of lncRNA GAS5 indicated a poor prognosis and enhanced cell proliferation in GC (Sun et al., 2014). lncRNA PVT1 indicated a poor prognosis of GC and promoted cell proliferation through epigenetically regulating p15 and p16 (Kong et al., 2015). lncRNA 00152 promoted tumor cell cycle progression by binding to EZH2 and repressing p15 and p21 in GC (Chen et al., 2016).
In our study, we focus on lncRNA ZEB1-AS1. To our knowledge, the current research is the first time to offer evidence for the function and mechanisms of ZEB1-AS1 in GC. In the present study, we found that ZEB1-AS1 expression was upregulated and positively correlated in advanced TNM stage, lymph node metastasis, and poor overall survival in GC patients. ZEB1-AS1 suppression reduced the proliferation and invasion of GC cells both in vitro and in vivo. Furthermore, our data revealed that ZEB1-AS1 functioned as a sponge of miR-335-5p in GC cells. Collectively, ZEB1-AS1 is critical for the proliferation and invasion of GC cells by regulating miR-335-5p. Our findings indicated that ZEB1-AS1 might offer a potential novel therapeutic target for the treatment of GC.
Recently, the ceRNA hypothesis proposed that lncRNAs communicate with other protein-coding RNA transcripts through shared common miRNA binding sites (Tay et al., 2014). This hypothesis was supported by a series of studies. For example, lncRNA SPRY4-IT1 promoted proliferation and invasion by acting as a ceRNA of miR-101-3p in colorectal cancer cells (Jin et al., 2017). lncRNA PVT1 promoted pancreatic cancer cell proliferation and migration through acting as a molecular sponge to regulate miR-448 (Zhao et al., 2017). lncRNA TUG1 promoted cervical cancer progression by regulating the miR-138-5p-SIRT1 axis (Zhu et al., 2017).
In our study, based on bioinformatics prediction and sequence analysis, we found that there was an interaction between ZEB1-AS1 and miR-335-5p. qRT-PCR showed that the expression of miR-335-5p was decreased and inversely correlated with ZEB1-AS1 expression in GC tissues. We further showed that miR-335-5p was increased after ZEB1-AS1 downregulation in GC cells, while overexpression of miR-335-5p did not affect ZEB1-AS1 expression. Luciferase reporter assay indicated that miR-335-5p overexpression reduced the luciferase activity of ZEB1-AS1-Wt. RNA immunoprecipitation experiments showed that both miR-335-5p and ZEB1-AS1 were in the Ago2-pulled down pellet. Moreover, results showed that miR-335-5p inhibition abrogated the suppression of proliferation and invasion of GC cells induced by ZEB1-AS1 inhibition.
Taken together, our results indicated that lncRNA ZEB1-AS1 could function as an oncogenic lncRNA in GC progression. It promoted cell proliferation and invasion through suppressing the tumor suppressor miR-335-5p. Consequently, these findings suggested that ZEB1-AS1 acted as a key role in GC pathogenesis and served as a potential therapeutic target for GC patients.
Footnotes
Acknowledgment
Thanks to the Center Lab of The First Affiliated Hospital of Xinxiang Medical University for their selfless help and support on our experimental technology and experiments.
Disclosure Statement
No competing financial interests exist.