
Abstract
Pyruvate kinase M2 (PKM2) has been wildly verified to modulate glycolysis in tumor cells. However, the role of PKM2 on the glycolysis of osteoarthritis (OA) chondrocytes is still unclear. In present study, we investigate the function of PKM2 on OA chondrocyte glycolysis and the collagen matrix generation in vitro. Results showed that PKM2 was upregulated in OA chondrocytes compared with healthy control chondrocytes. In OA chondrocytes, ATP expression was lower compared with healthy control chondrocytes. Loss-of-function experiment showed that PKM2 knockdown mediated by lentivirus transfection could significantly suppress the glucose consumption and lactate secretion levels and decrease glucose transporter-1 (Glut-1), lactate dehydrogenase A (LDHA), and hypoxia inducible factor 1-alpha (HIF-1α), indicating the inhibition of PKM2 knockdown on glycolysis. Moreover, Cell Counting Kit-8 (CCK-8), flow cytometry, and terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) assay showed that PKM2 knockdown inhibited OA chondrocyte proliferation and promoted the apoptosis. Western blot and immunocytochemical staining showed that PKM2 knockdown downregulated the expression levels of COL2A1 and SOX-9. In summary, our results conclude that PKM2 modulates the glycolysis and extracellular matrix generation, providing the vital role of PKM2 on OA pathogenesis and a novel therapeutic target for OA.
Introduction
O
Pyruvate kinase (PK) has been reported as one of the important regulators in reaction of glycolysis (Cruz et al., 2017; Panchabhai et al., 2017). Generally, normal cells generate the pyruvate kinase M1 (PKM1) isoform; however, the pathological cells usually switch to express pyruvate kinase M2 (PKM2) (Israelsen et al., 2013; Dayton et al., 2016). Glycolysis almost occurs in all living organisms and cells, which produce 2 molecules ATP and NADH (Li et al., 2017a). There are 10 parts of enzymic catalytic reaction in the glycolysis, and some of the key enzymes and intermediate products are closely related with cell development (Shang et al., 2017). PKM2 acts as the important rate-limiting enzyme in glycolysis and it usually overexpresses in proliferative cells and tumor cells, regulating the glycolysis progression and Warburg effect (Kremer et al., 2017). Shirai et al. (2016) reported that PKM2 serves a previous role as a molecular integrator of metabolic dysfunction, oxidative stress, and tissue inflammation on coronary artery disease through acting as a protein kinase that phosphorylates the transcription factor STAT3 and boosts IL-6 and IL-1β production (Shirai et al., 2016).
In present study, we assume that PKM participates in the energy metabolism of chondrocytes in OA pathogenesis. Therefore, series of functional experiments are designed to investigate the role of PKM2 on the proliferation, apoptosis, and glycolysis of chondrocytes.
Materials and Methods
Cell isolation and culture
Articular chondrocytes were isolated from the knee joints of six cases of total knee arthroplasty patients (age >60 years old) and six cases of trauma amputation patients (age <35 years old). During the surgery, articular cartilage samples were gained from tibial plateau. The chondrocytes were obtained using phased enzyme digestion method and cultured in vitro according to reported introduction (Chijiiwa et al., 2015). The chondrocytes at second passage were used for experiments, including OA chondrocyte group and healthy control group. All the experiments' design was approved by Ethics Committee of The Second Affiliated Hospital Zhejiang University School of Medicine. Informed consents were taken from every patient.
Lentivirus transfection and stable cell line construction
Lentiviral vector encoded with PKM2 sequence was designed and synthesized by GeneCopoeia (Guangzhou, China). The cDNA oligonucleotides targeting PKM2 (shPKM2) were synthesized by SBO Medical Biotechnology Company (Shanghai, China). The PKM2-shRNA sequence was shown as follows: PKM2-shRNA, Forward: 5′-AACGCTGTGGCTC TAGACACTAAACTCGAGTTTAGTGTCTAGAGCCACAGCTTTTTC-3′, Reverse: 5′-TCGAGAAAAAGCTGTGGCTC TAGACACTAAACTC GAGTTTAGTGTCTAGAGCCACAGCGTT-3′.
ATP, lactate, and glucose consumption detection
Chondrocytes (5 × 104/mL) were seeded into 24-well plates and incubated in incubator. ATP expression levels were measured using ATP Kit (Beyotime Institute of Biotechnology, Shanghai, China) according to manufacturer's instruction. Besides, lactate section levels were measured using Lactate Detection Kit (Jiancheng Biotechnology, Nanjing, China) according to manufacturer's instruction. Glucose consumption was detected using Glucose Detection Kit (Jiancheng Biotechnology, Nanjing, China) according to manufacturer's instruction.
Real-time PCR
Total RNA was isolated from cells using the RNeasy Mini Kit and DNase I according to the manufacturer's illustration (Qiagen, Hilden, Germany). cDNA was reverse transcribed from 1 μg using iScript cDNA Synthesis Kit (Bio-Rad, Hercules, CA). RT-PCR was performed using Real MasterMix SYBR ROX (Eppendorf, Hamburg, Deutschland). Primers in this study were shown as follows: PKM1, forward, 5′-AGTCATACGCGGAGAACCTC-3′, reverse, 5′-GCTATGTCCAGCGATTTGTT-3′; PKM2, forward, 5′-AGTACCATGCGGAGACCATC-3′, reverse, 5′-GCGTTATCCAGCGTGATTTT-3′; β-actin, 5′-CTCTTCCAGCCTTCCTTCCT-3′, reverse, 5′-AGCACTGTGTTGGCGTACAG-3′. β-actin acted as the control. The relative expression value was calculated with the 2−ΔΔCt method.
Proliferation ability
Proliferation ability of chondrocytes was measured using Cell Counting Kit-8 (CCK-8; Dojindo, Japan) according to manufacturer's instruction. Briefly, 1 × 104 cells were seeded into 96-well plates. For each well, 10 μL CCK-8 solution was added. Then, the cells were incubated at 37°C for 2 h. The absorbance at 450 nm was measured using a spectrophotometer every day.
Western blotting
Total protein and cytoplasmic and nuclear proteins were isolated from chondrocyte and tissue according to the manufacturer's instructions. Protein samples were subjected to SDS-polyacrylamide gel electrophoresis and transferred to PVDF membranes. The membranes were washed with TBST and blocked with 5% nonfat milk powder and incubated with primary antibodies at room temperature for 2 h and overnight at 4°C. Primary antibodies included anti-PKM1 (1:1000 dilution; Abcam), anti-PKM2 (1:1000 dilution; Abcam), anti-Glut1 (1:1000 dilution; Abcam), anti-lactate dehydrogenase (anti-lactate dehydrogenase A (LDHA), 1:1000 dilution; Abcam), anti-hypoxia inducible factor 1-alpha (anti-HIF-1α, 1:1000 dilution; Abcam), anti-COL2A1 (1:1000 dilution; Abcam), anti-SOX-9 (1:1000 dilution; Abcam), and anti-ADAMTS-5 (1:1000 dilution; Santa). Then, member was incubated with horseradish peroxidase-conjugated secondary antibody at room temperature for 2 h. The PVDF membrane was visualized using ImmunoStar LD (Wako Pure Chemical, Osaka, Japan) and measured using LAS-4000 Mini (FujiFilm, Tokyo, Japan). Relative protein expression was normalized according to β-actin.
TUNEL staining
Terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) was performed using Apoptosis Detection Kit (KeyGEN Bio, China) according to the manufacturer's instructions. Briefly, chondrocytes were washed using normal saline and then fixed with 4% neutral formaldehyde for 20 min at 4°C. Chondrocytes were treated with 0.2% Triton X-100 and immersed into 100 mL equilibration buffer. Cells were incubated with 50 μL TdT and stained by propidium iodide. Samples were examined under a laser-scanning confocal microscope (Zeiss, Germany).
Flow cytometry apoptosis
Chondrocytes were digested with trypsin and washed with phosphate-buffered saline. Briefly, cells (2 × 105) were suspended with 50 μL binding buffer, stained with Annexin V-PE/7-AAD Apoptosis Detection Kit (KeyGen, Nanjing, China). Cells were incubated with 5 μL 7-AAD for 15 min and 1 μL Annexin V-PE for 15 min. Then, the stained cells were measured by BD FACSCanto II Flow Cytometer (Becton Dickinson, Franklin Lakes, NJ).
Immunocytochemical staining
Chondrocytes were fixed in 10% formalin and embedded in paraffin. Briefly, after deparaffinization and rehydration, sections were incubated with anti-COL2A1 antibody overnight. Goat anti-mouse IgG antibody (1:1000; Invitrogen) was used as a secondary antibody. Finally, the sections were lightly counterstained with hematoxylin. The sections were observed under a confocal microscope (BX51WI; Olympus, Tokyo, Japan).
Statistical analysis
The data of experiments are presented as mean ± standard deviation and calculated using SPSS software (ver. 19.0; SPSS, Inc., Chicago, IL). Between-group variance was compared using one-way analysis of variance and Student's t-test. p < 0.05 was considered as statistical significance.
Results
PKM2 expression level was upregulated in OA chondrocytes
PKM1 and PKM2 expression chondrocytes were measured by western blot (Fig. 1A). Quantitative results showed that PKM2 protein expression level was significantly higher in OA chondrocytes than that in control healthy chondrocytes, instead of PKM1 (Fig. 1B). RT-PCR showed that PKM2 mRNA expression was significantly upregulated in OA chondrocytes than that in control chondrocytes (Fig. 1C). Western blot showed that the PKM2 protein expression was enriched in cytoplasm than that in nucleus (Fig. 1D, E). ATP level measurement showed that the ATP level in OA chondrocyte was lower compared with control chondrocyte. Besides, ATP level was significantly decreased in OA and control chondrocyte treated with 2-DG (glycolytic inhibitor, 20 mM), instead of OLIG (oxidative phosphorylation inhibitors, 20 μM) (Fig. 1F). In summary, results verified that PKM2 expression was upregulated in OA chondrocytes, and the major manner of energy source was glycolysis, instead of oxidative phosphorylation.
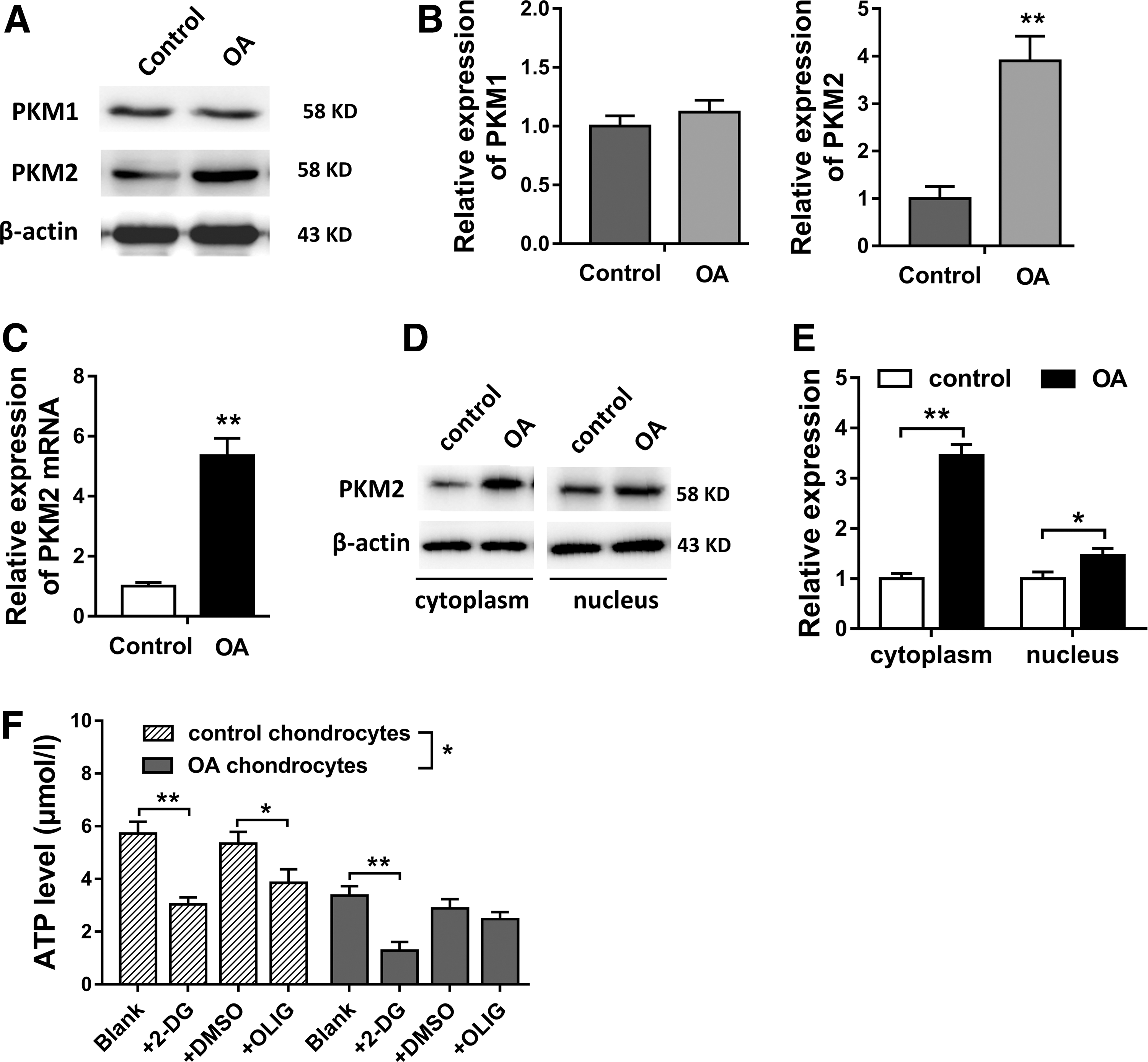
PKM2 expression was upregulated in OA chondrocytes, and ATP levels were decreased.
PKM2 knockdown suppressed the glycolysis in OA chondrocyte
It had been tested that PKM2 was upregulated in OA chondrocyte, and ATP level was decreased. Thus, in further research, we constructed PKM2 knockdown (shPKM2) using lentivirus transfection to test the role of PKM2 on glycolysis of OA chondrocyte. Results showed that shPKM2 transfection significantly decreased the PKM2 expression in OA chondrocyte, instead of PKM1 (Fig. 2A). ATP level measurement showed that the ATP level in shPKM2 group was lower compared with sh-NC and blank group (Fig. 2B). Moreover, glucose consumption and lactate secretion levels were both decreased in shPKM2 group compared with other group (Fig. 2C, D). Western blot showed that glucose transporter-1 (Glut-1), LDHA, and HIF-1α were decreased in shPKM2 group compared with sh-NC and control group (Fig. 2E, F). In summary, results revealed that PKM2 knockdown suppressed the glycolysis in OA chondrocyte.
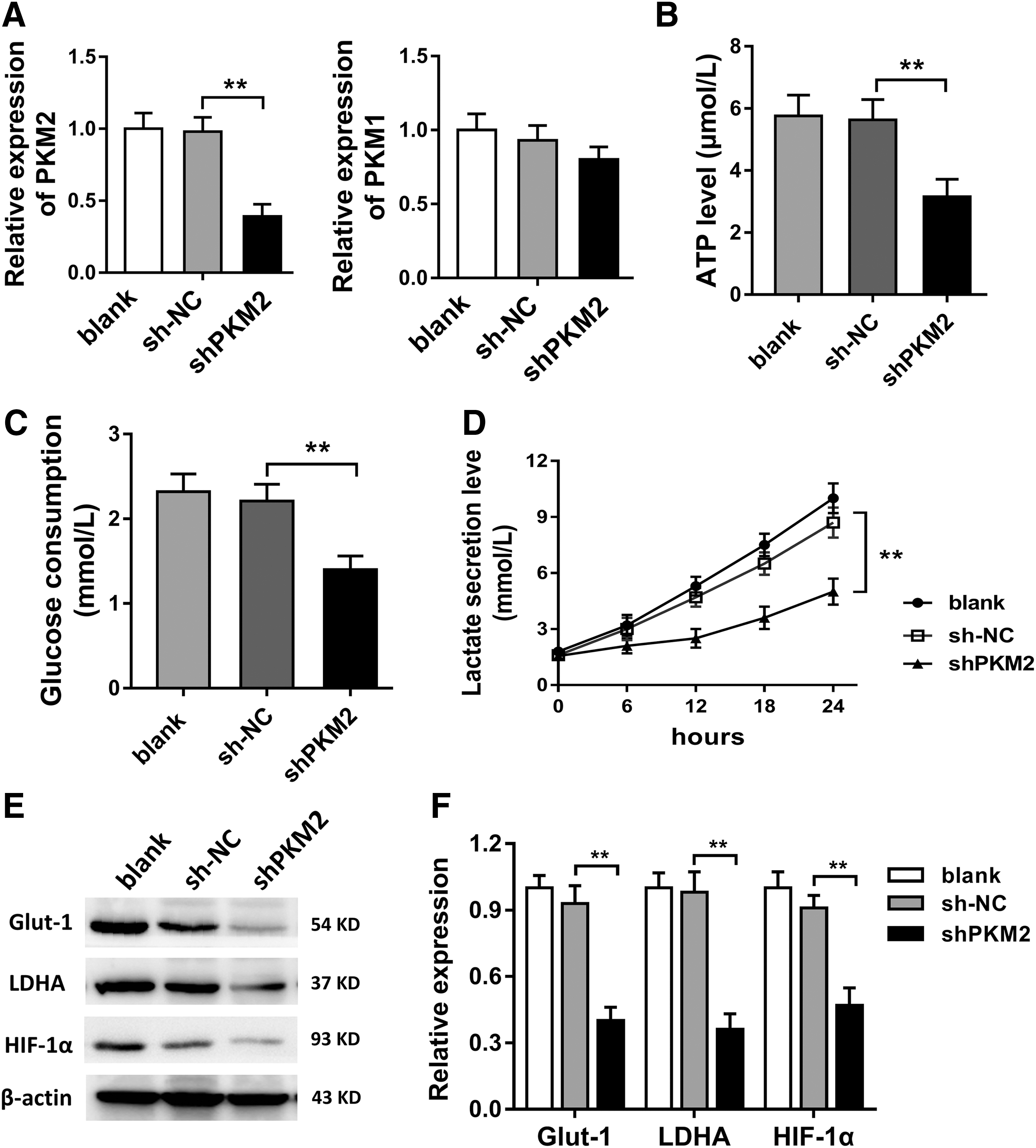
PKM2 knockdown suppressed the glycolysis in OA chondrocyte.
PKM2 knockdown suppressed the proliferation of chondrocyte in OA
Prior experiments had revealed that PKM2 knockdown suppressed the glycolysis of OA chondrocyte. For the phenotype of OA chondrocyte, functional experiments were performed to verify the proliferation characteristic. CCK-8 showed that PKM2 knockdown downregulated the proliferation vitality of OA chondrocyte compared with blank and sh-NC group (Fig. 3A). Flow cytometry showed that PKM2 knockdown increased the apoptotic cells compared with other groups (Fig. 3B, C). TUNEL staining showed that the TUNEL positive cell rate was significantly higher in PKM2 knockdown group than blank and sh-NC group, indicating the higher apoptotic rate (Fig. 3D, E). Taken together, results revealed that PKM2 knockdown suppressed the proliferation and promoted the apoptosis of chondrocyte in OA.
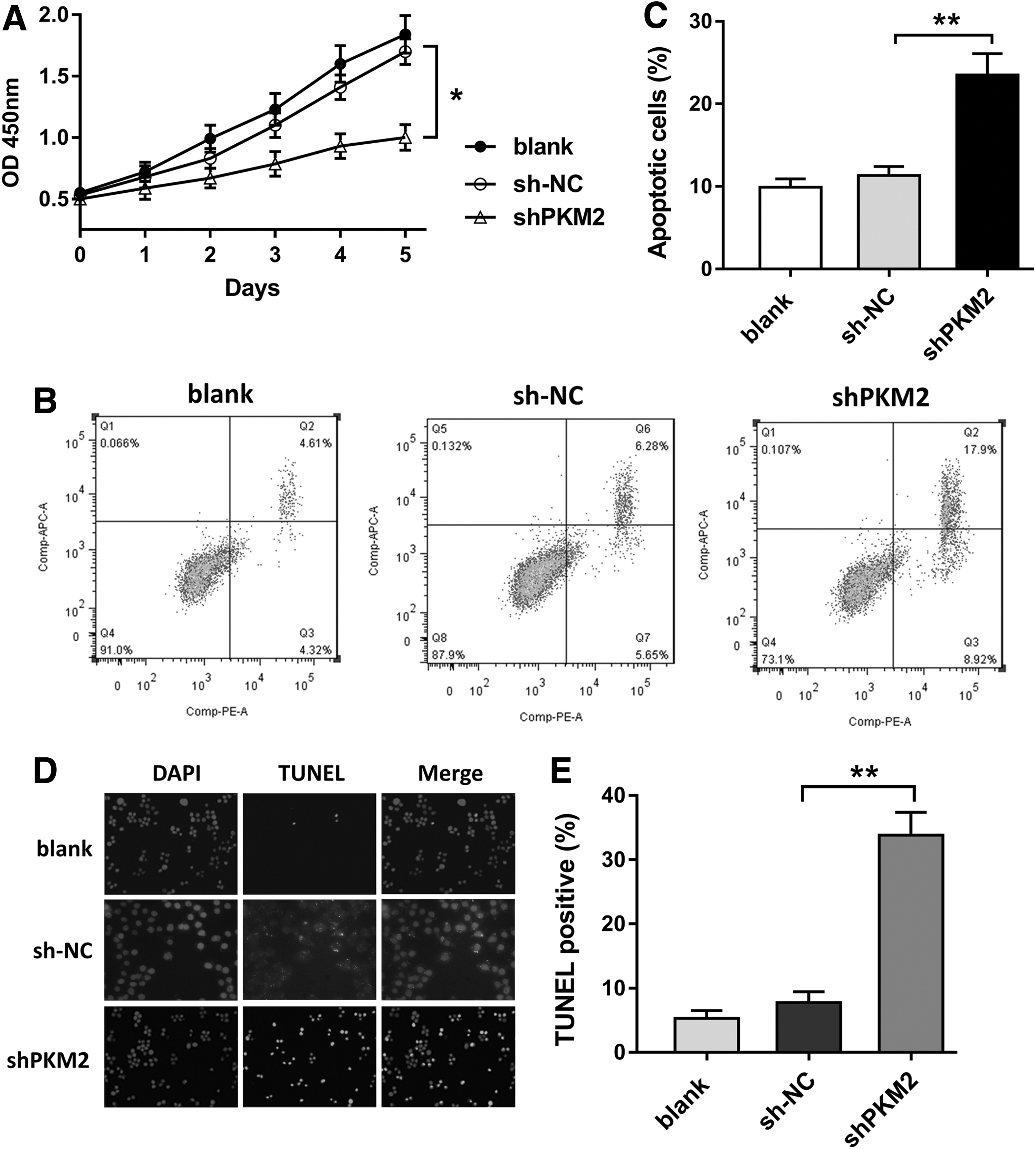
PKM2 knockdown suppressed the proliferation of chondrocyte in OA.
PKM2 knockdown suppressed the ECM generation of chondrocyte in OA
To investigate the role of PKM2 on articular cartilage ECM generation, we performed western blot to measure the expression levels of COL2A1, SOX-9, ADAMTS-5, and ADAMTS-4. Results showed that PKM2 knockdown downregulated the expression of COL2A1 and SOX-9 in OA chondrocyte (Fig. 4A, B). PKM2 knockdown upregulated the expression level of ADAMTS-5. Immunocytochemical staining showed that the COL2A1 expression was lower in PKM2 knockdown group than that in sh-NC group (Fig. 4C, D). Overall, results showed that PKM2 knockdown suppressed the ECM generation of chondrocyte in OA.
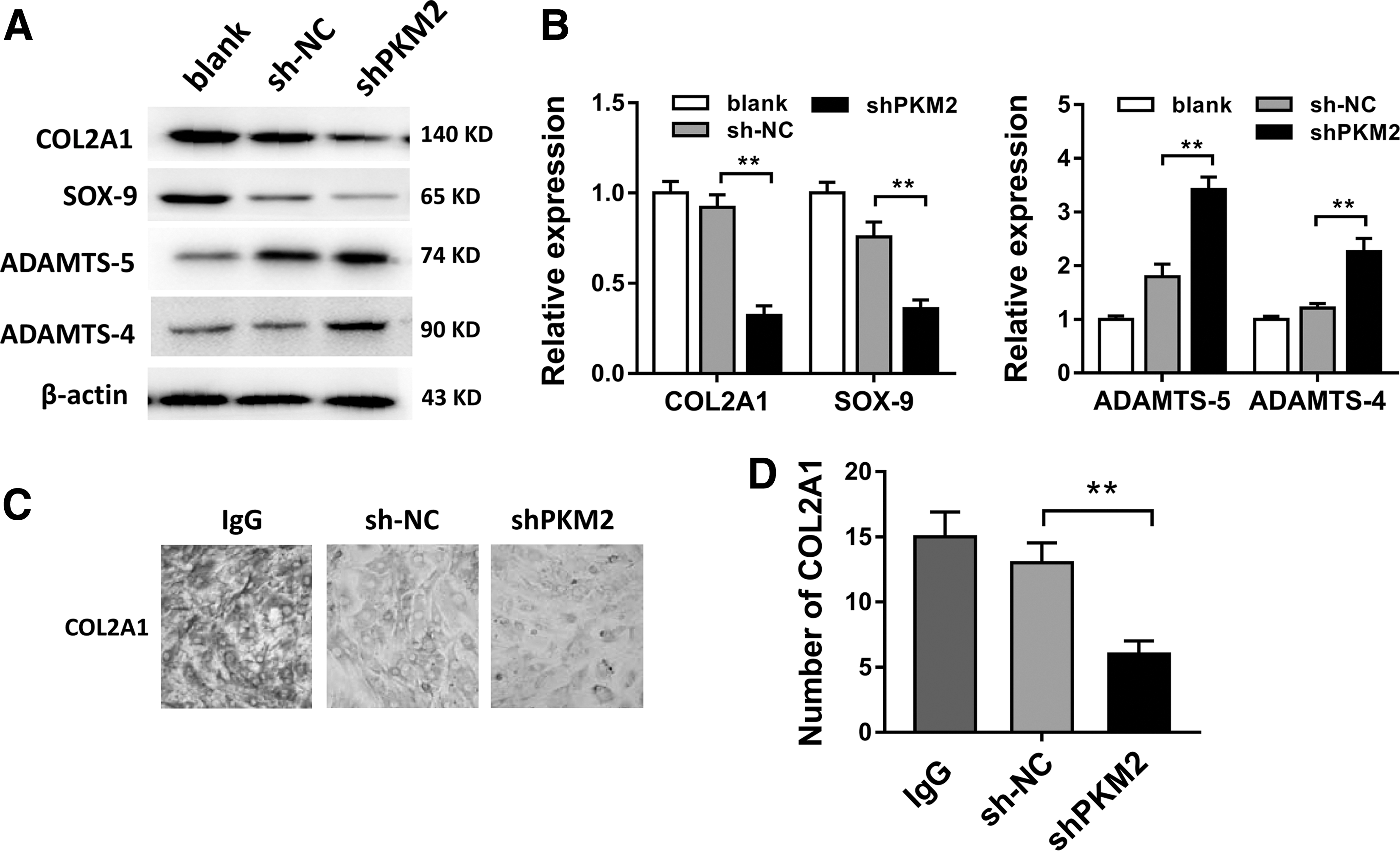
PKM2 knockdown suppressed the extracellular matrix generation of chondrocyte in OA.
Discussion
OA is a kind of retrogressive arthropathy accompanied by increasing morbidity as patients grow older (Aaron et al., 2017; Rahmati et al., 2017). There are series of factors in the pathogenetic process; however, the accurate molecular mechanism is still unclear. Abundant researches confirm that the degeneration of articular cartilage is one of the major pathogeneses for OA. Chondrocytes is the only cell in the articular cartilage, whose biological characteristic variation is closely correlated with the occurrence and development of OA (Li et al., 2017b). In present study, we focus on the energy supplying of chondrocytes and investigate the role of PKM2 on chondrocyte glycolysis.
PKM2 is the major regulatory enzyme in glycolysis progression, which catalyzes phosphoenolpyruvic acid (PEP, ADP) to pyruvic acid and ATP. In tumorous cells, the glycolysis is active and accounts for the main energy source. Besides, PKM2 acts as the key regulatory factor in “Warburg effect” (Kremer et al., 2017; Panchabhai et al., 2017). Palsson-McDermott et al. (2015) reported that LPS-induced PKM2 enters into a complex with Hif-1α by directly binding to the IL-1β promoter, and activation of PKM2 by TEPP-46 in vivo inhibited LPS and Salmonella Typhimurium-induced IL-1β production, promoting the inflammatory response. In our study, we found that PKM2 was upregulated in the isolated OA chondrocytes; moreover, the ATP level in OA chondrocytes was decreased compared with control chondrocytes. Therefore, we assume that PKM2 participates in the ATP generation regulation. When OA chondrocyte was treated with 2-DG (glycolytic inhibitor, 20 mM), ATP level was significantly decreased; however, ATP level remained the same when treated with OLIG (oxidative phosphorylation inhibitors, 20 μM). Thus, we confirmed that the major manner of energy source is glycolysis, instead of oxidative phosphorylation. The reason might be due to the hypoxia environment of OA chondrocyte. Besides, western blot showed that the PKM2 protein expression was enriched in cytoplasm and nucleus, especially cytoplasm. Previous studies have indicated that the cytoplasmic PKM2, in tumors, is mainly tetramer form, indicating the prominent function on glycolysis of PKM2 (Giannoni et al., 2015).
Because PKM2 is upregulated in OA chondrocyte, we construct PKM2 knockdown using lentivirus transfection to validate the role of PKM2 knockdown on OA chondrocyte. Results showed that the ATP level in shPKM2 group was lower compared with sh-NC and blank group. Moreover, glucose consumption, lactate secretion levels, Glut-1, LDHA, and HIF-1α were decreased in shPKM2 group. HIF-1α is a common component that controls the cellular metabolism and has a role on cellular effector functions, acting as a key reprogrammer of metabolism in inflammatory cells that promotes inflammatory gene expression (Corcoran and O'Neill, 2016). Thus, we conclude that PKM2 knockdown suppresses the glycolysis in OA chondrocyte. Although glycolysis is the major manner to provide energy for chondrocyte, the mitochondria oxidative phosphorylation still coexists in chondrocyte (Lee et al., 2016; Hosseinzadeh et al., 2017). Besides, the calorific value and energy accommodate of oxidative phosphorylation are more effective than glycolysis (Ye et al., 2016; Ma et al., 2017). Therefore, when the PKM2 and glycolysis are decreased, the oxidative phosphorylation recovers and provides more ATP than ever.
Moreover, our results revealed that PKM2 knockdown suppressed OA chondrocyte proliferation and promoted apoptosis. The chondrocyte ATP acts as the energy material and plays important role in chondrocyte proliferation, differentiation, and function maintenance (Fang et al., 2016). Besides, the glycolysis abnormal might lead to endothelial dysfunction, even chondrocyte fertilization (Xing et al., 2017). Matsumoto et al. (2017a, 2017b) reported that the glycolysis dysfunction could cause chondrocyte fertilization, ECM degradation, and arthropathy. In general, the ATP quantity generated by anaerobic glycolysis is markedly lower compared with mitochondria aerobic oxidation. In pathological OA chondrocyte, the excessive enhancement of glycolysis led to the reduction of energy acquisition and then suppresses the proliferation, differentiation, and cell viability (Qu et al., 2016).
The pathogenesis of OA is mainly due to the hydrolysis of cartilage ECM protein, and the main ingredients of cartilage ECM are polysaccharide (Hadipour-Jahromy and Mozaffari-Kermani, 2010). Our results revealed that PKM2 knockdown decreased collagen matrix generation in OA chondrocyte. Type II collagen was the main constituent of cartilage collagen. SOX-9 had high expression in embryonic cartilage and controlled the II collagen synthesis, thus affecting the cartilage generation. Polysaccharide protein enzyme family (ADAMTS) was the main degradation enzyme for proteoglycan in cartilage matrix proteins. Therefore, we conclude that PKM2 knockdown decreased the energy accommodation and then attenuated extracellular collagen matrix generation in OA chondrocyte, providing articular cartilage degeneration in OA.
In summary, our study investigates the role of PKM2 on the energy metabolism of chondrocytes in OA pathogenesis. Results validate the function of PKM2 knockdown on the proliferation, apoptosis, and glycolysis of chondrocytes, providing a novel therapeutic target for OA.
Footnotes
Acknowledgments
This work was supported by Zhejiang Provincial Natural Science Foundation of China (LY15H060003) and National Natural Science Foundation of China (81401785).
Disclosure Statement
No competing financial interests exist.