
Abstract
The protein O-glucosyltransferase 1 (Poglut1) links O-glucose to epidermal growth factor-like repeats harboring the C1X
Introduction
D
Although SC express numerous membrane proteins, which distinguish them from surrounding cell types (Yin et al., 2013), the paired-box transcription factor Pax7 represents their universal marker (Seale and Rudnicki, 2000). Pax7 plays a key role in early postnatal myogenesis through regulation of muscle precursor cell proliferation. Knockout of Pax7 in mice results in an absence of SCs, which severely impacts muscle mass and leads to death (Seale et al., 2000). Moreover, inhibition of Pax7 in mouse myoblasts (MBs) induces a diminution of MyoD expression (von Maltzahn et al., 2013).
MyoD is a myogenic determination factor, which regulates muscle cell differentiation by inducing cell cycle arrest, a prerequisite for myogenic initiation, and whose expression is activated by Pax7 (Buckingham, 2007). Mice knocked out for MyoD have altered muscle regeneration due to defects of SCs to initiate the process of differentiation (Megeney et al., 1996). Muscle cells can be readily distinguished by the expression of Pax7 and MyoD transcription factors. Quiescent SCs are Pax7+MyoD−, proliferating cells, Pax7+MyoD+, and differentiating cells, Pax7−MyoD+ (Zammit et al., 2006; Shan et al., 2016). Thus, Pax7/MyoD ratio is critical in cell fate determination (Wen et al., 2012).
The Notch signaling pathway is highly conserved in animals; in mammals, there are four Notch receptors (Notch1 to Notch4) and five ligands (Dll1, Dll3, Dll4, Jagged1, and Jagged2). The canonical Notch signaling pathway is initiated by the interaction between the extracellular domains of a Notch receptor and its ligand located on an opposite cell (de Celis, 2013). Sequential enzymatic cleavages by ADAM proteases and the ϒ-secretase complex lead to a release of the Notch intracellular domain (NICD) into the cytoplasm. Cleaved NICD represents the active form of the receptor, which is translocated to the nucleus. It interacts with RBP-Jk, the primary transcriptional mediator of Notch signaling, by displacing corepressors and recruiting coactivators to enhance the expression of target genes, which belong to the Hes and Hey family genes such as Hes1 and Hey1 (Iso et al., 2003).
Both extracellular domains of Notch receptor (NECD) and ligands contain numerous epidermal growth factor (EGF)-like repeats. EGF-like repeats are cysteine-containing motifs of about 40 amino acids found in many transmembrane and secreted proteins, among which Notch is the most studied (Takeuchi and Haltiwanger, 2014; Haltom and Jafar-Nejad, 2015). They are posttranslationally modified by a variety of N- and O-glycans, thus regulating the activity of Notch signaling pathway (Stanley and Okajima, 2010). On the Notch receptor, three unusual forms of O-glycosylation can occur at consensus sites located on EGF-like repeats: O-fucosylation, O-glucosylation, and O-GlcNAcylation (Takeuchi and Haltiwanger, 2014; Vasudevan and Haltiwanger, 2014; Harvey et al., 2016). They are involved in correct folding of the receptor and ligand binding (Jafar-Nejad et al., 2010; Taylor et al., 2014).
Some O-fucosylated EGF-like repeats, as EGF 12 of mouse NOTCH1, were shown to be necessary for ligand binding (Rampal et al., 2005). However, the impact of O-glycosylation on Notch activity is definitively more complex with, for example, on the murine NOTCH1, 20 potential sites of O-fucosylation, 17 sites of O-glucosylation, and 11 sites of O-GlcNAcylation (Sawaguchi et al., 2017). Protein O-fucosyltransferase 1 (Pofut1), protein O-glucosyltransferase 1 (Poglut1), and EGF repeat-specific O-GlcNAc transferase (eOgt) are the enzymes whose the activity toward Notch receptor has been demonstrated (Stahl et al., 2008; Fernandez-Valdivia et al., 2011; Sakaidani et al., 2011).
Perturbed O-glycosylation of Notch receptor can dramatically affect its activity for both ligand binding and receptor activation (Stanley and Okajima, 2010; Takeuchi and Haltiwanger, 2010), whereas O-fucosylation of ligands, as murine DLL1, has been shown dispensable (Müller et al., 2014). In mouse, Pofut1 knockout induces embryonic lethality at E9.5 and a phenotype similar to the lack of core downstream Notch signaling components (Shi and Stanley, 2003). Embryos lacking Pofut1 are especially defective in somitogenesis.
Recently, we demonstrated that Pofut1 knockdown reduces Notch signaling in mouse myogenic C2C12 cell line (Der Vartanian et al., 2015). In addition, reduction of Pofut1 expression decreases Pax7 expression and disrupts the expression of myogenic regulatory factors (MRFs) such as MyoD. This results in the depletion of progenitor cells, PAX7+MYOD−, which is associated to an earlier myogenic program entrance. The differentiation of SC-derived MBs isolated from Pofut1 cax/cax mice, presenting hypomorphic alleles of Pofut1, shows similar results (Al Jaam et al., 2016). In vivo, these mice exhibited postnatal skeletal muscle hypertrophy without hyperplasia, and a decrease in SC number due to an earlier entrance in differentiation.
Poglut1 adds O-glucose to a serine residue included in the C1X
In vivo, Poglut1−/− mouse embryos die before E9.5 with posterior axis truncation and severe defects in neural tube development, somitogenesis, cardiogenesis, and vascular remodeling (Fernandez-Valdivia et al., 2011). Their phenotype is more dramatically affected than in case of Notch mutants, suggesting that Poglut1 acts earlier through other protein targets (Ramkumar et al., 2015). Recently, a missense mutation in human POGLUT1 gene was associated with muscular dystrophy (Servián-Morilla et al., 2016). In vitro and in vivo experiments demonstrate that the mutation significantly reduces the O-glucosyltransferase activity on Notch and the pool of quiescent PAX7+ cells.
In this study, we provide evidence that Poglut1 downregulation disturbed myogenic program through the Notch signaling, resulting in a premature and increased differentiation with altered myotube phenotypes. Expression of Pax7 and MyoD was modified and the PAX7/MYOD ratio was reduced, which led to depletion in the pool of unfused cells. Together, our results indicate that POGLUT1 has a crucial role on Notch pathway activation during myogenic differentiation.
Materials and Methods
Cell culture
C2C12 mouse MBs (American Type Culture Collection–ATCC, Manassas, VA) were cultured at 37°C and 5% CO2 in a growth medium (GM) with DMEM (Gibco®, Life Technologies™, Carlsbad, CA) supplemented with 10% fetal calf serum (Eurobio, Courtaboeuf, France), 4 mM L-glutamine, 50 U/mL penicillin, and 50 μg/mL streptomycin. Cells were plated at a density of 10,000 cells per cm2. When MBs reached confluence growth, the medium was removed and cells were cultured in a differentiation medium (DM). DM consists of DMEM supplemented with 2% horse serum, 4 mM L-glutamine, 50 U/mL penicillin, and 50 μg/mL streptomycin (Gibco) and was routinely changed every 24 h. For experimental purposes, cells were harvested by trypsinization (0.125% trypsin and 0.125 mM EDTA) for 5 min at 37°C.
Samples named MB corresponded to C2C12 cells at time 0 h, defined by substitution of GM by DM, and myoblast in differentiation (MBd) corresponded to C2C12 cells from 12 h to 48 h in DM. At 72 h and 120 h after DM addition, separation of myotubes (MT) and reserve cells (RC) was achieved by a short trypsinization (0.1% trypsin and 0.1 mM EDTA, 30 s) that specifically removed MT and only left RC adherent to the flask. Nuclei number per myotube and total nuclei number were counted in differentiated cells after immunostaining, by using ImageJ 1.45 s software (Abramoff et al., 2004) to estimate the fusion percentage.
Generation of Poglut1 knockup cell lines
Poglut1 short hairpin RNAs (shRNA) were designed according to rules described by Reynolds et al., 2004. The Poglut1 knockdown cell lines were obtained using electroporation (Gene PulserXcell™ transfection system; Biorad, Hercules, CA), by transfecting 3.106 C2C12 MB with 3 μg of pSilencer 2.1-U6-neoconstruct (Ambion, Life Technologies, Carlsbad, CA) containing an insert encoding a small interfering RNA (siRNA) against the mouse Poglut1 3′UTR region (5′-GATCCTCGGGTCATCTACGTGTGTTATTCAAGAGATAACACACGTAGATGACCCGATTTTTTGGAAA-3′) for knockdown 1 (Pg- KD1) or exon 2 (5′-GATCCATGAACCATGTTCAAGTCAAATTCAAGAGATTTGACTTGAACATGGTTCATTTTTTTGGAAA-3′) for knockdown 2 (Pg- KD2). Clone selection was performed using 1 mg/mL neomycin (Roche Applied Science, Mannheim, Germany). Wild-type C2C12 and two stably transfected cell lines previously modified by Der Vartanian et al. (2015), a control C2C12, with a negative control siRNA template (Ctrl shRNA), and a knockdown C2C12 for Pofut1 (Po-), were also used.
Phenotypic analysis
Cells were fixed with 4% paraformaldehyde (PFA) for 20 min, permeabilized with 0.1% Triton X-100-phosphate-buffered saline (PBS) for 30 min at 4°C, and blocked with PBS containing 20% of goat serum for 1 h at room temperature. Cells were then incubated overnight at 4°C with the mouse anti-myosin Heavy Chain antibody (dilution 1:2,000; Sigma-Aldrich Corp., St Louis, MO). After three washings with 0.1% Tween-20 PBS, cells were incubated for 1 h at room temperature with Alexa Fluor® 488 goat anti-mouse IgG (H+L) (Invitrogen, Life Technologies, Carlsbad, CA) and washed thrice with 0.01% Tween 20-PBS. To complete the staining, cell nuclei were labeled for 5 min, at room temperature, in 1 μg/mL DAPI solution (Sigma-Aldrich Corp.). Cells were rinsed five times with PBS, mounted with Mowiol® 4–88, and sealed with glass coverslips. For each sample, eight randomized visual fields were acquired with Leica fluorescence microscope at 40 × magnification. The fusion percentage corresponded to the percentage of nuclei-included myotubes. Myotubes were categorized into five different groups according to their nuclei content (<5, ≥5–10, >10–20, >20–40, >40). The morphometric parameters (diameter and length) were measured with ImageJ.
Gene expressions and data analysis
Total RNA was extracted using the RNeasy mini kit (Qiagen, Inc., Hilden, Germany). Quality and quantity of total RNA were measured using an Agilent 2100 bioanalyzer (Santa Clara, CA) and a Nanodrop 1000 spectrophotometer (Wilmington, DE), respectively. Two micrograms of total RNA were used to synthesize cDNA with the High Capacity cDNA Reverse Transcription Kit (Applied Biosystems®, Life Technologies, Carlsbad, CA). The amplification reactions were performed with 2 ng of cDNA following the manufacturer's instructions. Gene expressions were determined by semiquantitative real-time RT-PCR on an ABI Prism 7900 Sequence Detector System using the TaqMan® probe-based chemistry (Applied Biosystems). TaqMan primers and probe sets used in this study were as follows: 18S (Hs99999901_s1), GAPDH (Mm99999915_g1), Pofut1 (Mm00475567_m1), Poglut1 (Mm00552419_m1), Pax7 (Mm03053796_s1), MyoD1 (Mm00440387_m1), Rbpj (Mm00770450_m1), Hes1 (Mm00468601_m1), and Hey1 (Mm00468865_m1).
Data were collected using the QuantStudio Analysis software (Applied Biosystems). mRNA relative quantity (RQ) values were calculated only for a threshold cycle (Ct) lower than 38 and compared using the ΔΔCt method, as previously described (Der Vartanian et al., 2015) with 18S and GAPDH reference genes. mRNA relative quantity (RQ) was quantified by comparison of each differentiation time (from 0 to 120 h) relative to 0 h for WT cells, which was set as 1.
Western blot analysis
Proteins were extracted from C2C12 cell lines with RIPA extraction buffer (50 mM Tris–HCl, 150 mM NaCl, 0.5% sodium deoxycholate, 1% NP-40, and 0.1% SDS, pH 8) containing a protease inhibitor cocktail (Roche Applied Science). Protein lysates were then centrifuged at 12,000 g, 10 min at 4°C. Supernatant was recovered and protein quantity was estimated using the bicinchoninic acid protein (BCA) assay (Sigma-Aldrich Corp.) with bovine serum albumin (BSA) as a standard. Fifty micrograms or 100 μg (in case of cleaved NICD detection) of extracted proteins were separated under denaturing and reducing conditions into 12% or 6% (NICD) polyacrylamide gel, and then transferred to Hybond C-Extra Nitrocellulose membrane (GE Healthcare, Buckinghamshire, United Kingdom). To prevent unspecific antibody binding, membrane was saturated 2 h at room temperature with 5% nonfat dried milk in 0.1% Tween-20 Tris-buffered saline (TBST, 50 mM Tris–HCl, 150 mM NaCl, and 0.1% Tween-20, pH 7.4) or blocking buffer (5% BSA in 0.1% TBST) for cleaved NICD detection. Specific primary antibodies were diluted in their blocking buffer overnight at 4°C.
The primary antibodies detecting the following proteins were used: cleaved NOTCH 1 (Val 1744) from Cell Signaling Technology (Danvers, MA), GAPDH from R&D Systems (Minneapolis, MN), MYOD1 (clone 5.8A) from Dako Cytomation (Glostrup, Copenhagen, Denmark), PAX7 from Developmental Studies Hybridoma Bank (University of Iowa, IA), POFUT1 (Der Vartanian et al., 2015) and POGLUT1, an anti-peptide produced and purified by Agro-Bio (La Ferté St-Aubin, France). After three washes of 5 min each in 0.1% TBST, HRP-coupled secondary antibodies were incubated in 2.5% nonfat dry milk in 0.1% TBST or 2.5% BSA in 0.1% TBST for cleaved NICD, for 1 h at RT, and washed thrice. Immunoblots were revealed by enhanced chemiluminescence using BM Chemiluminescence western blotting substrate (peroxidase, POD; Roche Applied Science) and exposed (Hyperfilm ECL; GE Healthcare, Buckinghamshire, United Kingdom). For relative quantification, analysis of band intensities was carried out using ImageJ. All band intensities were normalized to the corresponding GAPDH band intensity. Then, band intensities were compared to WT C2C12 MB one at 0 h, settled to 1.
Statistical analysis
All experiments were performed in biological triplicates and represented by the mean ± SEM. Statistical comparisons were performed using one-way ANOVA or ANCOVA test implemented in Past®, exe 3.1 (Hammer et al., 2001). If ANOVA revealed significant effects, it was corrected by the Bonferroni test. A p-value of 0.05 or less was considered statistically significant. On figures, no asterisk denoted p > 0.05, one asterisk p < 0.05, double asterisk p < 0.01, and triple asterisk p < 0.001.
Results
Stable C2C12 cell lines knockdown for Poglut1
To investigate the role of protein-O-glucosyltransferase 1, POGLUT1, during the onset myogenic differentiation, we established stable Poglut1 knockdown cell lines with two different shRNAs, shRNA 1 targeting the 3′UTR region of Poglut1 and shRNA 2 targeting Poglut1 coding region. Four stable clones were obtained with shRNA 1 and six with shRNA 2. By semiquantitative real-time PCR, we evaluated the mRNA expression levels of Poglut1 in the different clones. Three shRNA 1 C2C12 clones showed significant downexpression of Poglut1, between 53% and 81%, relative to WT C2C12; four shRNA 2 C2C12 clones downexpressed Poglut1, between 19% and 42% (data not shown). For further studies, we chose the clone that displayed the strongest downregulation of Poglut1 (81%), this clone was named Pg- KD1, and a clone that presented a more moderated downregulation (42%), this clone was named Pg- KD2 (Fig. 1A). We also used, for comparison, a C2C12 cell line knockdown for Pofut1, Po- (40% downexpression), since both POFUT1 and POGLUT1 participate to Notch O-glycosylation on EGF-like domains (Moloney et al., 2000; Takeuchi et al., 2017). We had therefore controlled that the downexpression of Poglut1 had no effect on Pofut1 expression and vice versa (Fig. 1B). All the significant downexpressions of Poglut1 and Pofut1 were confirmed at the protein level (Fig. 1C, D).
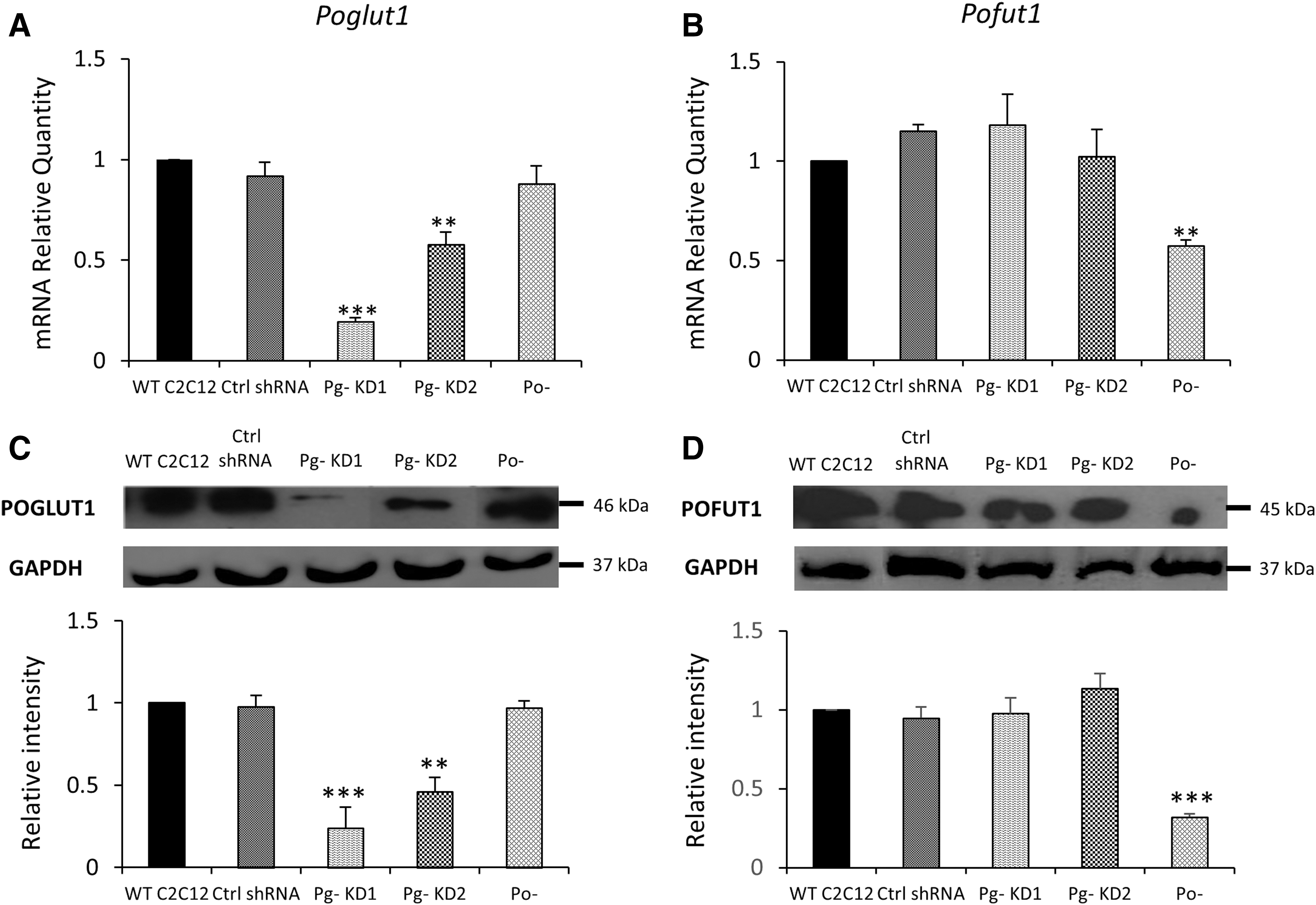
Characterization of the Poglut1 knockdown C2C12 cell lines. Relative quantities (RQ) of Poglut1
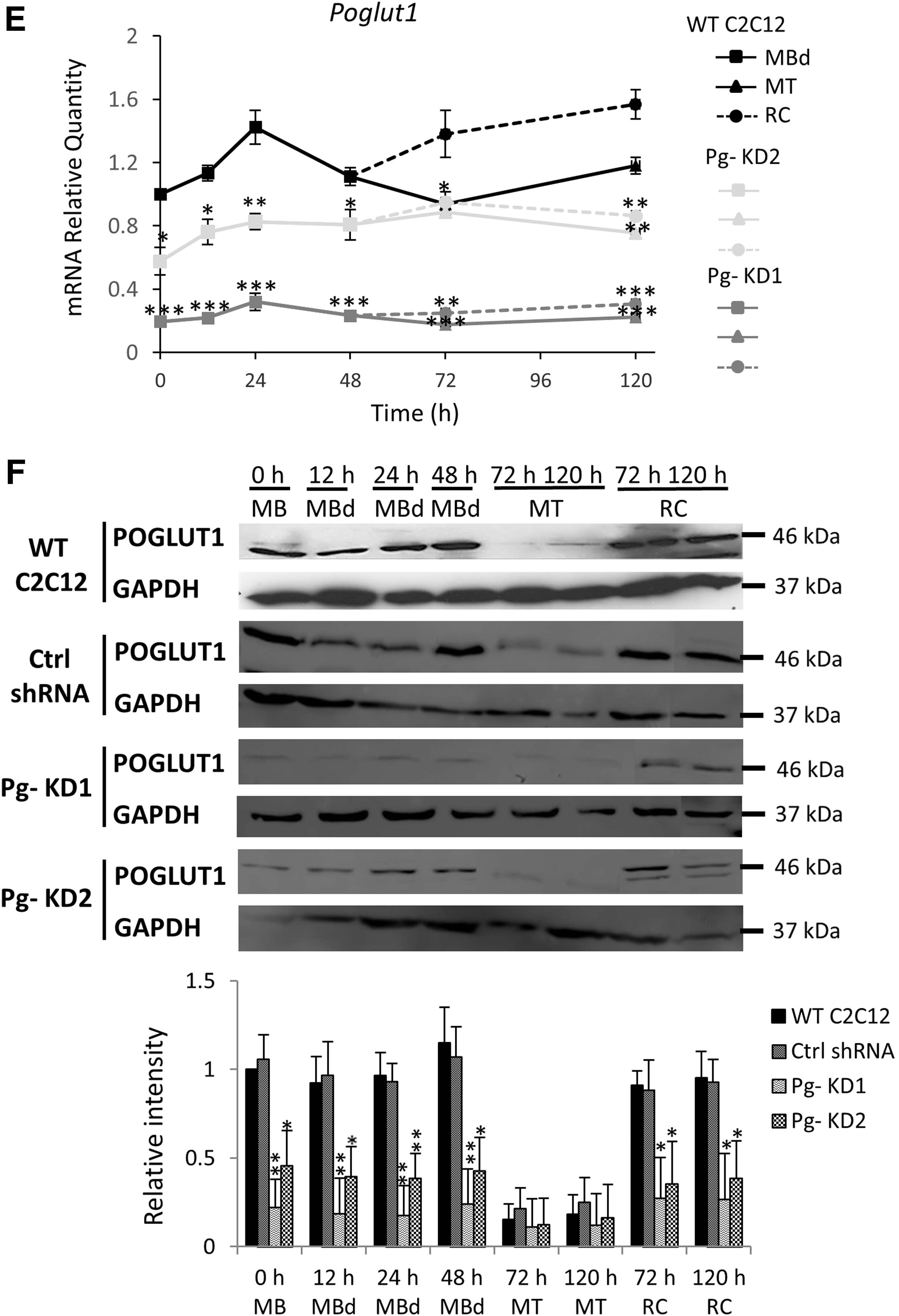
Poglut1 expression was followed during the first 120 h of differentiation time course (Fig. 1E), in which differentiated myoblasts (MBd) fuse into multinucleated cells, myotubes (MT), leaving some undifferentiated cells, RC, which can start a new differentiation process if replated. In WT C2C12, Poglut1 expression increased during the first 24 h. Beyond 72 h of the differentiation time course, where two distinct populations of cells (MT and RC) can be separated, it was significantly more expressed in RC than in MT. In Pg- KD1 and Pg- KD2 cell lines, Poglut1 was significantly downexpressed compared to WT C2C12, except for time 72 h in Pg- KD2 MT. Expression in RC was always higher compared to MT, but no more statistically supported (Fig. 1E). During the differentiation time course, downregulation of Poglut1 was significantly stronger in Pg- KD1 than in Pg- KD2. Poglut1 expression was reduced by 80% and 71% in Pg- KD1 MT and by 74% and 64% in Pg- KD1 RC, at 72 and 120 h respectively, relative to Pg- KD2. Poglut1 expression profiles were confirmed at the protein level (Fig. 1F) with a significant reduction of POGLUT1 in both Pg- cell lines from 0 h to 48 h. POGLUT1 was poorly detected in MT and unlike mRNA expression, did not show any significant difference between cell lines. Otherwise, Pg- KD1 and Pg- KD2 RC showed around three to four times less of POGLUT1 at 72 and 120 h, relative to WT RC. The Poglut1 downexpression in Pg- KD1 and Pg- KD2 cells was maintained along the first 120 h of the myogenic process, however, without a significant difference between MT and RC, which could suggest modification in the cell differentiation program.
Poglut1 reduction affects MB fusion
Poglut1 and Pofut1 are enzymes known to impact activation of Notch signaling pathway involved in cell commitment (Takeuchi and Haltiwanger, 2014). In our study, phenotypic consequences of Poglut1 knockdown were appreciated during myogenic differentiation. First, we visualized, by fluorescence microscopy, myotube phenotypes during the first 120 h of differentiation. MT at 120 h showed no apparent difference between WT and control shRNA C2C12 cells, as revealed by DAPI and myosin staining (Fig. 2A). WT C2C12 MT are long, fusiform, and elongated in several directions. The strong downregulation of Poglut1 in Pg- KD1 cells induced earlier apparition of branched MT compared to controls. They were clearly thicker and longer with agglomerated nuclei, different from the even distribution seen in control cells. Pg- KD2 MT also appeared thicker than controls, but less than in Pg- KD1 and much shorter, with apparently lower number of nuclei per MT. Their phenotypes were different from Po- ones, which presented more elongated and thinner MT with a low number of nuclei (Der Vartanian et al., 2015). All these phenotypes reflected modified fusion processes.

Poglut1 knockdown promotes myogenic differentiation in C2C12 cells and decreases the pool of reserve cells.
The fusion percentages were then calculated by determining the percentage of nuclei in myotubes relative to the total number of nuclei, during 120 h of differentiation. No significant difference was observed between WT and control shRNA C2C12 cells (Fig. 2B). In Pg- KD cell lines, downregulation of Poglut1 increased the fusion percentage, compared to control cells. In Pg- KD1 cells, where Poglut1 expression was strongly decreased (by 81% in MB), the fusion percentage was significantly increased from 24 h (12% vs. 4% in WT C2C12) to 120 h (59% vs. 39% in WT C2C12) of differentiation. In Pg- KD2 cells, which displayed a moderate Poglut1 downregulation (by 42% in MB), the fusion was also increased, but only significantly at 48 h and 72 h (17% and 29% vs. 10% and 25% in WT C2C12). A significant increase of fusion percentage was also observed in Po- cells from 24 h (9% vs. 4% in WT C2C12) to 120 h (48% vs. 39% in WT C2C12).
Globally, the differentiation process occurred earlier in Pg- cell lines compared to WT or control shRNA C2C12 cell lines and was more pronounced when the downregulation of Poglut1 was strong. After addition of differentiation medium, cell proliferation in controls remained persistent up to 48 h, whereas in Pg- KD1 and Pg- KD2 cell lines, it immediately dropped, suggesting a premature differentiation, which could explain the increase of fusion percentage (data not shown). This precocious cell cycle exit would explain the significant decrease in number of mononucleated cells at 72 h and 120 h of differentiation in Pg- KD, compared to control cell lines.
Poglut1 reduction alters myotube phenotype
Computation of fusion percentages did not allow appreciating the heterogeneity in the number of nuclei per MT. Consequently, to better characterize the myotube phenotypes, we counted the number of nuclei per MT in all cell lines. WT and control shRNA C2C12 cell lines had a mean of 9 and 8 nuclei per MT, respectively, at 120 h, whereas Pg- KD1 had an average of 26 nuclei per MT. In Pg- KD2 cells, where Poglut1 downregulation was more moderated, only 6 nuclei were counted per MT at 120 h of differentiation. The results obtained for Pg- KD cells reflect important fusion defects. A similar (even more serious) disruption of MB fusion was observed in Po- cells with an average of four nuclei per MT at 120 h.
To clarify the phenotype changes, we categorized myotubes into five groups (<5, ≥5–10, >10–20, >20–40, >40 nuclei per MT) for each cell line at 72 h and 120 h of differentiation (Fig. 3A). No significant differences were observed between WT C2C12 and shRNA control cells. Pg- KD1 cells presented a distribution of MT categories significantly different from controls with a diminution of the category with less than five nuclei (38% and 9% in Pg- KD1 vs. 52% and 35% in WT C2C12, at 72 h and 120 h, respectively), whereas the categories with more than 20 nuclei were significantly increased (14% and 53% in Pg- KD1 vs. 2% and 4% in WT C2C12, at 72 h and 120 h, respectively). In contrast, Pg- KD2 cells had significantly more MT with a small number of nuclei (70% and 60% of MT had less than five nuclei, at 72 h and 120 h, respectively). No MT with more than 20 nuclei was observed in Pg- KD2 at 72 h and less than 1% at 120 h. Compared to Po- cells at 120 h, the distribution of Pg- KD2 MT categories was not significantly different.
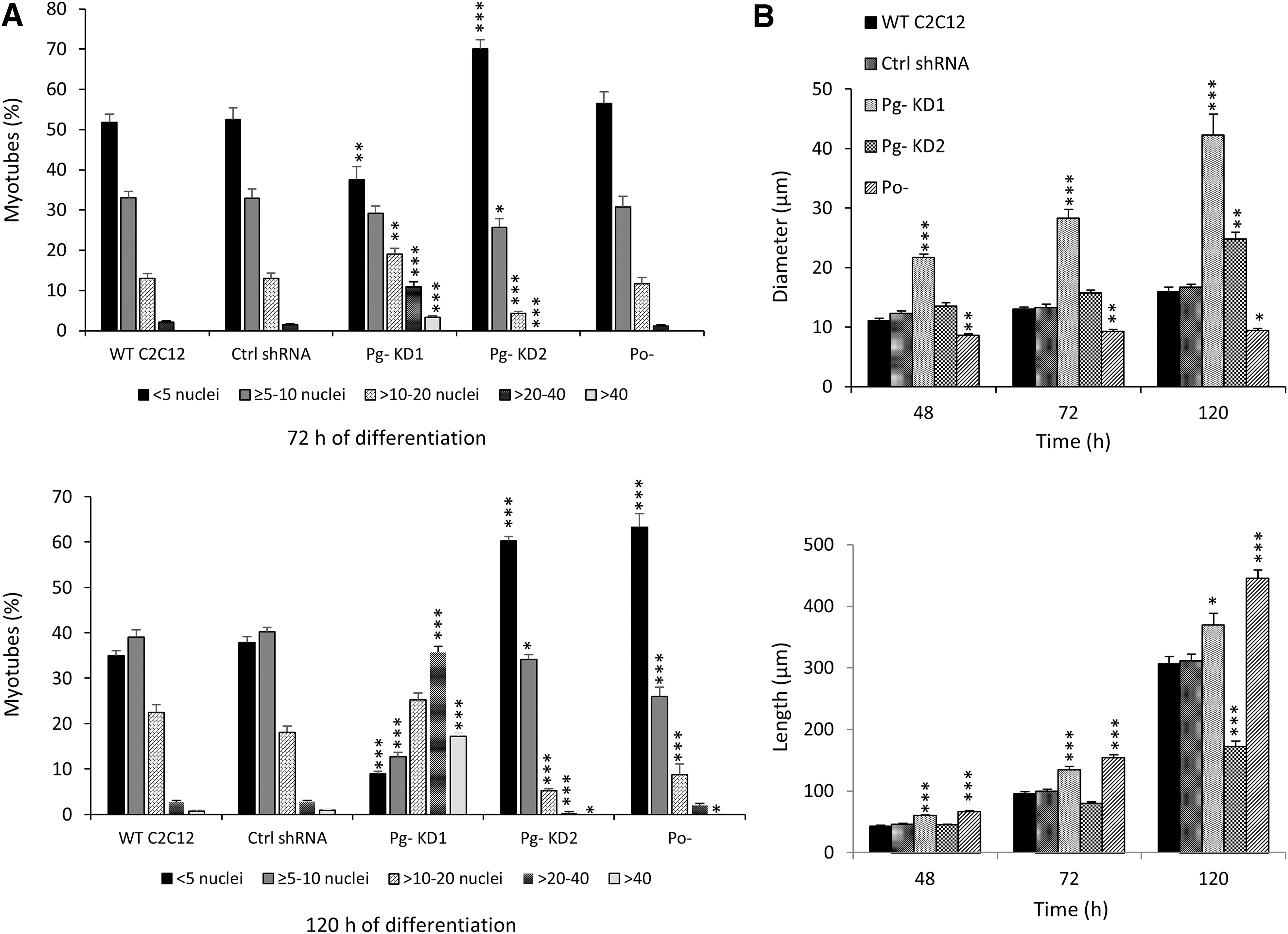
Poglut1 knockdown alters myotube phenotype during myogenic differentiation of C2C12 cells.
Therefore, a moderate downregulation of Poglut1 or Pofut1 expression (by around 40%) leads to the same reduced capacity of cell fusion into multinucleated myotubes. This phenomenon was not accentuated in Pg- KD1 cells, where Poglut1 was strongly reduced. In that case, compared to all cell lines, MB fusion seemed to be promoted since MT with more than 10 nuclei were more represented.
To complete our phenotypic observations, we measured the diameters and the lengths of MT at 48 h, 72 h, and 120 h of differentiation in WT, control shRNA, Pg- KD, and Po- C2C12 cell lines (Fig. 3B). No significant difference was observed between WT and control shRNA C2C12 cells. At 120 h, WT C2C12 MT had an average of 16 μm of diameter and 307 μm of length. In Pg- KD1 cells, MT diameter and length were significantly greater than controls from 48 h to 120 h. Pg- KD1 MT had a mean of 42 μm of diameter and 369 μm of length at 120 h. In Pg- KD2 cells, MT length was shorter than controls, only being significant at 120 h with 172 μm. However, their diameter was significantly greater than controls at 120 h, with 25 μm. Compared to Pg-, Po- MT were characterized by their small diameter (9 μm) and stretched shape (445 μm long).
Taken together, our results showed that MT phenotypes were differently affected, depending on the level of Poglut1 deregulation. When it is moderated, MB fusion is severely reduced, whereas when Poglut1 is strongly downregulated, MB fusion higher than in control cells is allowed. We can hypothesize that beyond an important level of Poglut1 downexpression, regulation mechanisms of MB fusion are defective and this could explain the runaway of cells toward a differentiated state.
Knockdown of Poglut1 reduces Notch signaling pathway
Altered MT phenotypes could result from abnormal activation of Notch receptor. So, the active form of Notch receptor, the cleaved Notch intracellular domain (NICD) was followed by Western Blot during 120 h of differentiation time course. NICD was strongly detected from 0 h to 48 h in WT differentiating MBs and then, mostly detected in RC (Fig. 4A). In Pg- cell lines, the NICD profiles were significantly lowered by 50% to 95% compared to WT C2C12 cells, depending on the differentiation time and cell type considered. The decrease was already observed for both Pg- KD at the start of differentiation and more accentuated in Pg- KD1 compared to Pg- KD2, which correlated with the difference of phenotypes. Cleaved NICD was also more abundant in RC than in MT. However, its presence did not prove that the Notch signaling pathway was correctly activated.
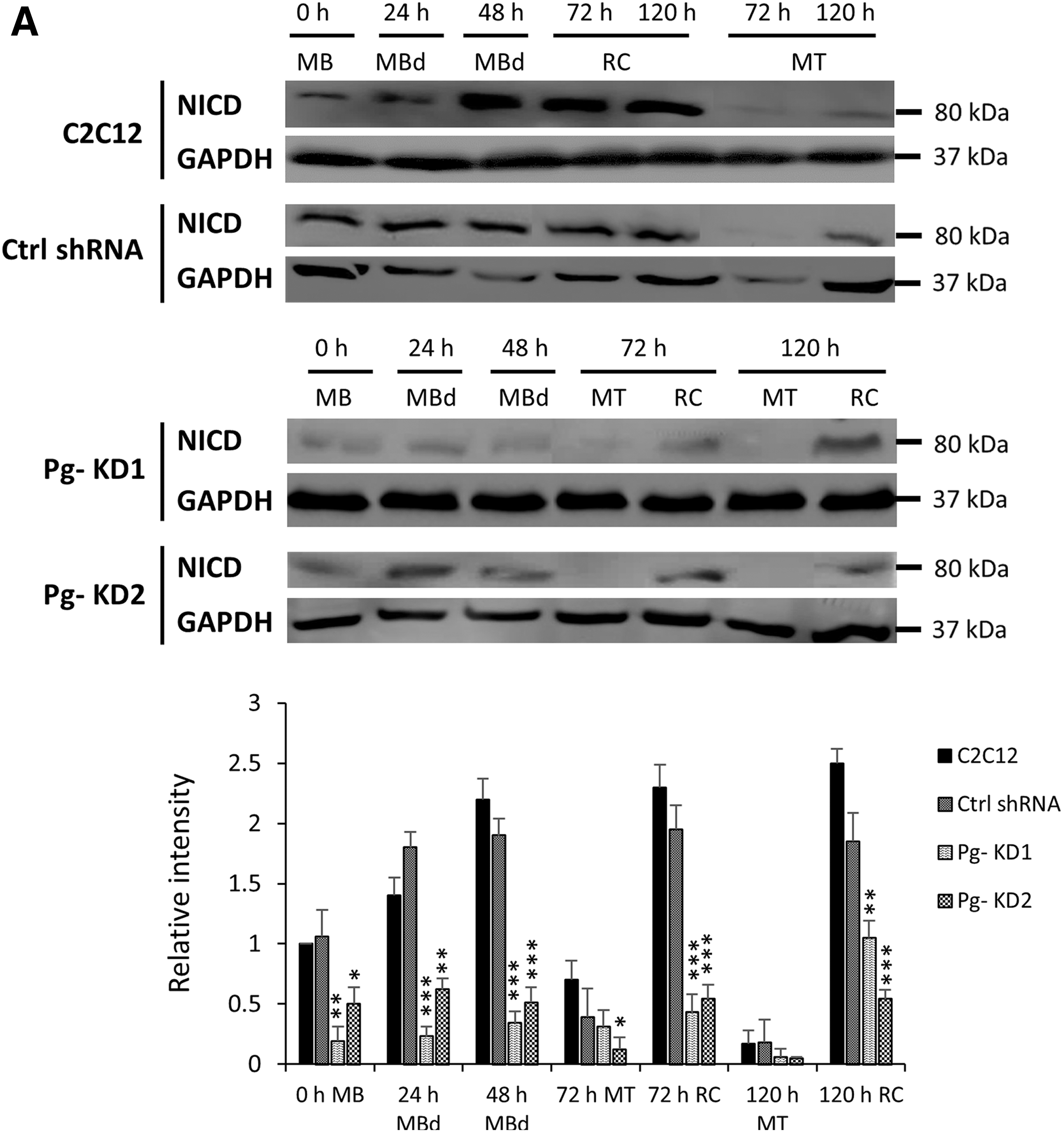
The knockdown of Poglut1 affects the Notch signaling pathway activation.
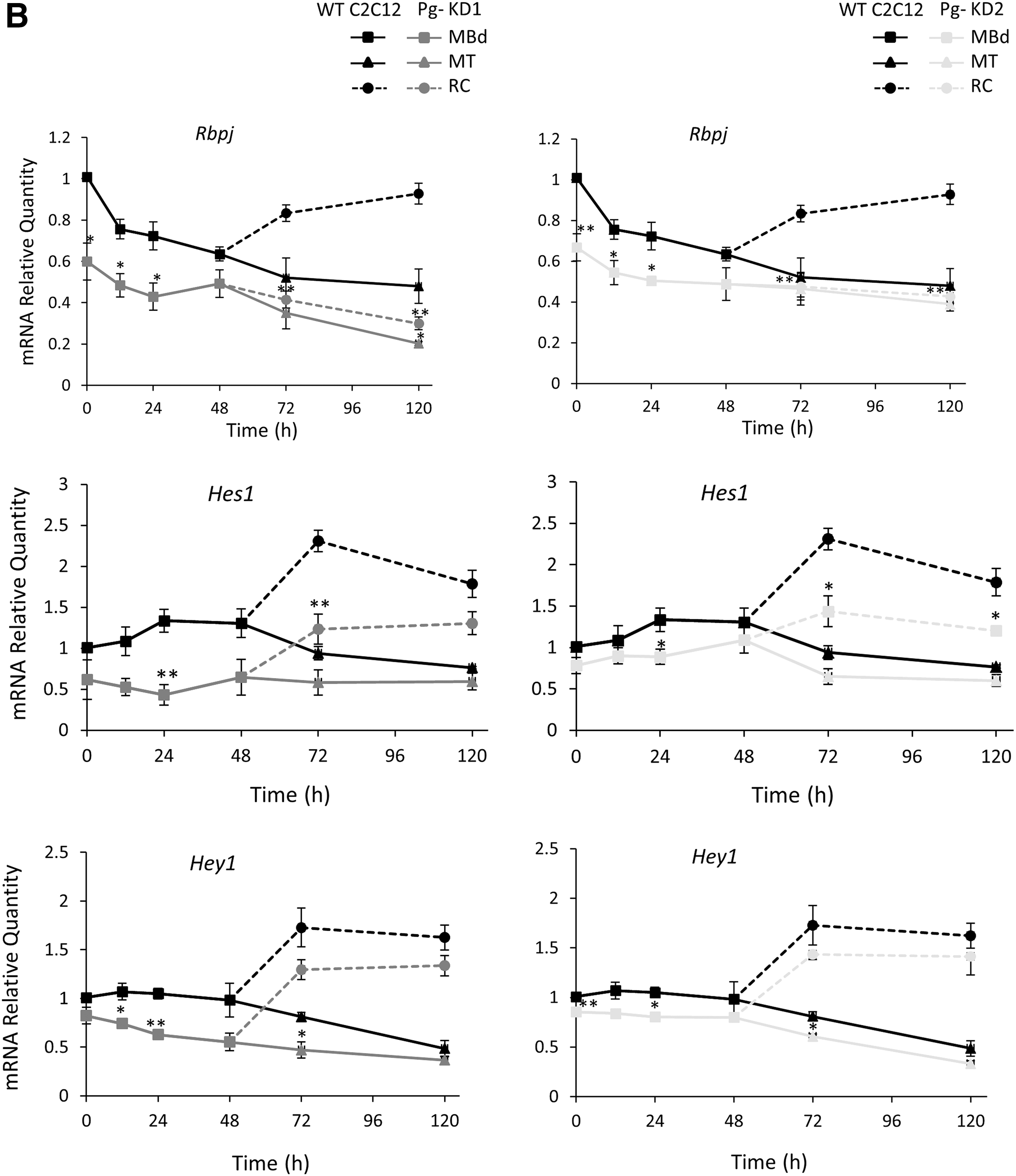
Thus, we analyzed expression of Rbpj (DNA binding transcription factor), a nuclear partner, which forms with NICD a transcriptional coactivation complex in the nucleus. Final activation of Notch signaling was measured by expression of some downstream Hes and Hey family genes (Hairy/Enhancer of split genes encoding transcriptional inhibitors). In all cell lines, expression of Rbpj decreased throughout the differentiation process and was higher in RC than in MT (Fig. 4B). In WT C2C12 cells, Rbpj expression had decreased by 30% to 52% from 24 h to 120 h. In Pg- KD cells, the phenomenon was accentuated with a decrease of 40% and 33% in Pg- KD1 and Pg- KD2, respectively, at the beginning of the time course, compared to WT. A significant reduction of Rbpj expression was also observed in RC (50% and 68% in Pg- KD1 and 43% and 54% in Pg- KD2, at 72 h and 120 h, respectively, relative to WT C2C12).
Expressions of Hes1, Hes6, and Hey1 were also modified during C2C12 differentiation. Hes1 and Hey1 whose promoters are targets for NICD were more expressed in RC than in MT, regardless of the C2C12 cell line considered (Fig. 4B). Hes1 expression decreased in both Pg- KD cells, however, more significantly at the onset of differentiation for Pg- KD1 and in RC for both, compared to WT. In Pg- KD1 cells, Hes1 expression was significantly decreased from 12 h of differentiation by 52% compared to WT, and at 72 h by 38% in MT and by 48% in RC. In Pg- KD2, the decrease of Hes1 expression was less pronounced (reduction by 17% at 12 h, and by 31% in MT and by 38% in RC at 72 h). Hey1 expression was reduced in both Pg- KD cells and it was more pronounced in Pg- KD1 than Pg- KD2 cells. As a control, Hes6 expression (not induced by cleaved NICD) was enhanced during the first 120 h of differentiation with a higher level in MT compared to RC, but without significant differences between WT and Pg- cells (data not shown).
During myogenesis, Hes1 and Hey1 have inhibitory activities on the expression of myogenic genes (Buas and Kadesch, 2010) and in muscle progenitor cells, Hes1 represses expression of cyclin-dependent kinase inhibitor p57, essential for cell growth arrest (Zalc et al., 2014). Their downexpression in Pg- KD could explain the early cell cycle exit of proliferative MBs and their commitment into myocytes ready to initiate the fusion process, a phenomenon particularly accentuated in Pg- KD1 cells.
The Knockdown of Poglut1 affects the Pax7/MyoD expression balance
Pax7/MyoD ratio is decisive in muscle cell fate commitment
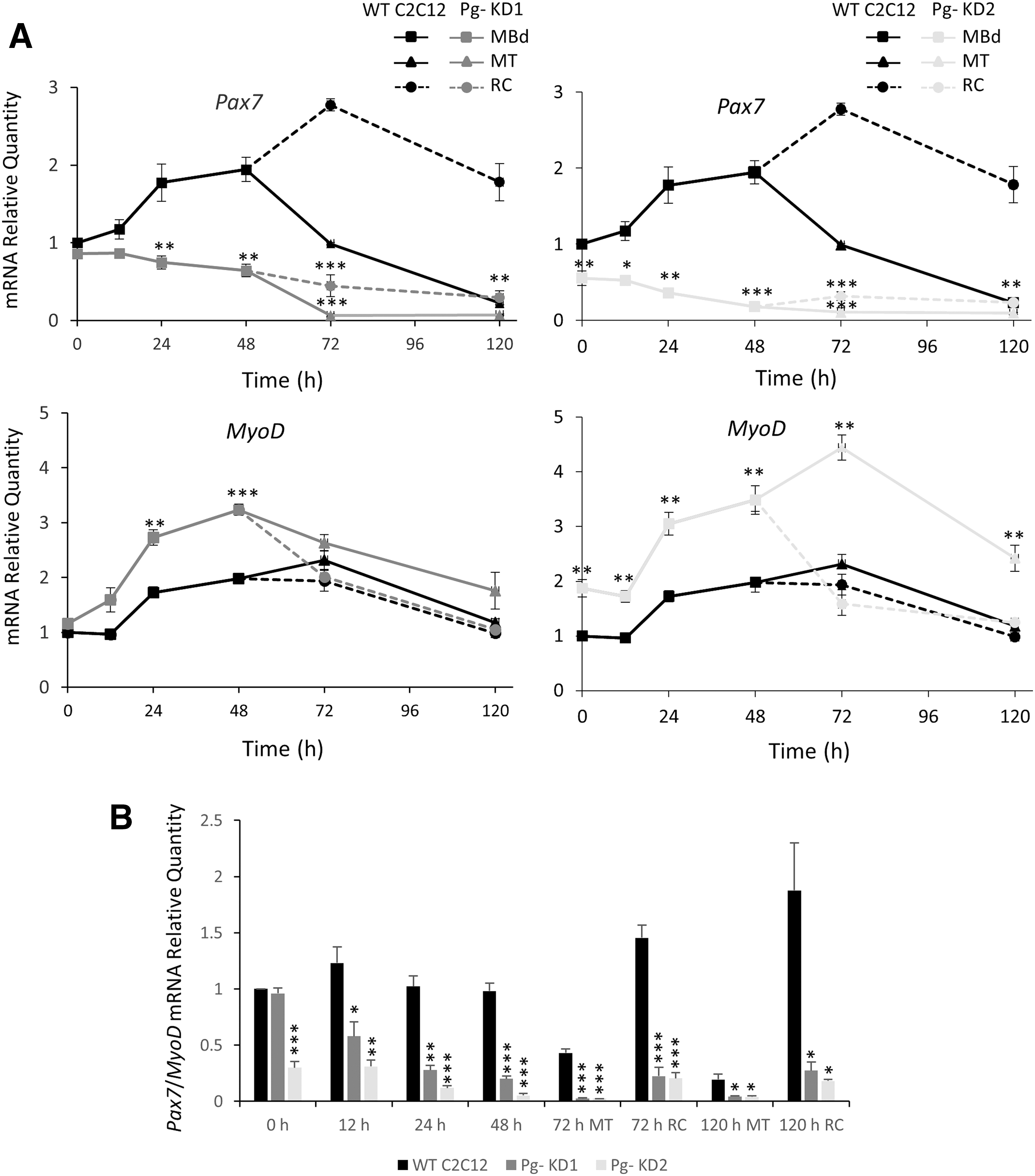
The knockdown of Poglut1 affects the timing of myogenic differentiation.
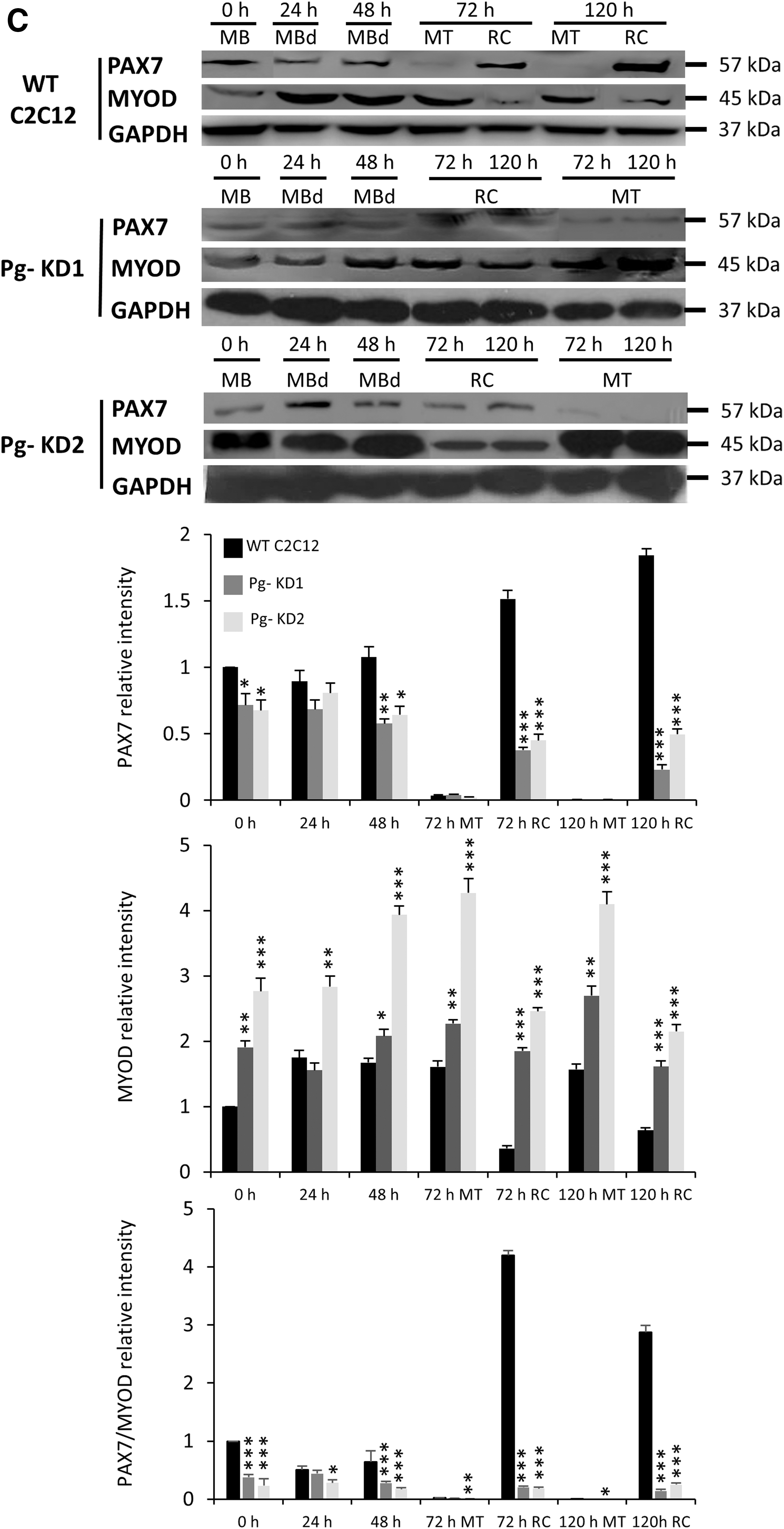
Western Blot experiments showed that PAX7 was poorly detected in MT and, unlike mRNA expression, did not show any significant difference between cell lines (Fig. 5C). Pg- KD1 RC expressed 88% and Pg- KD2 73% less of PAX7, relative to WT C2C12 RC at 120 h. MYOD was more present in MT than in RC, independent of the cell line considered. It was always significantly more expressed in Pg- KD2 than in Pg- KD1. In RC, MYOD was significantly increased compared to WT C2C12, with for example, 2.5 and 3.4 times more of MYOD in Pg- KD1 and Pg- KD2 at 120 h, respectively. Disruption of PAX7/MYOD ratio confirmed that Poglut1 knockdown alters the myogenic program. Our results showed that RC are particularly sensitive to decreased Poglut1 expression.
In summary, Poglut1 knockdown in mouse C2C12 cells results in a significant reduction of Notch signaling, which leads to a drastic decrease of Pax7 expression, critical for RC maintenance. A decreased pool of proliferative PAX7+ cells induces a premature and increased differentiation observed by the MYOD increase, which disturbs myogenic process, alters myotube phenotypes, and could impair the ability of RC to reengage a myogenic differentiation program.
Discussion
In this study, we present evidence that POGLUT1, an enzyme responsible for O-glucosylation of proteins, is crucial for Notch pathway activation by which it governs, in murine C2C12 MBs, cell fate decisions during myogenic differentiation. Our in vitro experiments show that the knockdown of Poglut1 in C2C12 cells alters Notch signaling pathway by a significant reduction of quantity of cleaved Notch intracellular domain (NICD), a decrease in the expression of Rbpj and Notch downstream genes (Hes1 and Hey1), and finally a disruption of Pax7/MyoD expression balance.
The moderate downregulation of Poglut1 expression in Pg- KD2 cell line (around 40% less than in WT C2C12 cells) and the strongest Poglut1 downregulation in Pg- KD1 cell line (around 80% less than in WT C2C12 cells) induce different myotube phenotypes depending on the level of Poglut1 reduction. We used Po- cell line to compare the effects of Pofut1 knockdown to the Poglut1 deregulation in C2C12 cells. Pg- KD2 and Po- cells have the same level of Poglut1 or Pofut1 reduction, but show some different myotube phenotypes. It reflects a different role of glucose or fucose on glycoproteins, even if both sugar residues could exist on the same EGF-like repeat. Based on the consensus sequences, a database screening with their respective consensus O-glycosylation sites (Rana et al., 2011; Schneider et al., 2017) gives around 50 protein targets for mouse POGLUT1 and 100 for POFUT1; among the first, 90% are also demonstrated or predicted targets for POFUT1. Notch receptors and their ligands are the best studied and biologically demonstrated to be highly modified by O-glucose and O-fucose (Takeuchi and Haltiwanger, 2014). However, elimination of a single O-glucosylation site was demonstrated to only have a limited effect on Notch1 activity (Rana et al., 2011), possibly due to compensatory effects of the other O-glucosylated sites. In our study, we determine that Poglut1 knockdown largely affects the Notch signaling pathway, suggesting the absence of several O-glucoses on the 17 potential O-glucosylation sites. Indeed, Fernandez-Valdivia et al. (2011) demonstrated by mass spectroscopy that decrease in Poglut1 expression in C2C12 cells causes a reduction in the level of O-glucosylation of mouse Notch2 EGF-like repeats. The differences found between Pg- KD1 and Pg- KD2 cells may be explained by the involvement of other O-glucosylated targets. Essentially, knockout of Poglut1 in mouse induces an embryonic lethality with phenotypic and functional defects similar to a global loss of Notch signaling, but occurring slightly earlier than knockout of Notch (Fernandez-Valdivia et al., 2011; Ramkumar et al., 2015), supporting the involvement of other targets.
Among the potential targets of mouse POGLUT1, peptidase domain-containing protein associated with muscle regeneration 1, PAMR1, protocadherin FAT2, which is involved in migration of epidermal cells, but also expressed in skeletal muscles and fibulin-7, and an adhesion glycoprotein that interacts with extracellular matrix molecules in developing teeth, also expressed in muscles, could be specifically involved in peculiar Poglut1 knockdown phenotypes. They all possess a unique potential O-glucosylation site always associated in the same EGF-like repeat with a potential O-fucosylation site. Recently, Takeuchi et al. (2017) demonstrated that the addition of O-glucose and O-fucose glycans stabilizes a single EGF-like repeat in an additive and nonredundant manner, participating to its correct folding and preserving its biological role.
Knockdown of Poglut1 in C2C12 cells induces an earlier entrance in the differentiation process, around 24 h before WT C2C12 cells, causing defects in MB fusion and inducing an altered phenotype of myotubes. MB fusion takes place in two steps: the primary fusion where MBs fuse together, forming nascent myotubes, and the secondary fusion where MBs fuse with immature myotubes to increase the number of nuclei and the size of myotubes. In Pg- KD2 MT, from 12 h of differentiation, we have observed that the increase of MyoD expression is higher than in Pg- KD1 cells and could explain a more premature fusion. Pg- KD2 MT are characterized by a small number of nuclei in MT, suggesting that the primary fusion is not altered, but the secondary fusion is. Pg- KD1 MT present a significant increase of differentiation from 24 h compared to WT C2C12 or Pg- KD2 cells, and have significantly more nuclei in MT than all cell lines, suggesting that both the primary and secondary fusions are favored or at least the secondary fusion is.
We can hypothesize that in Pg- KD1 MT, where nuclear accretion is considerable at the end of time course differentiation, the “brake” acting on MB fusion in Pg- KD2 cells can be lifted when Poglut1 is strongly reduced. So, according to the level of Poglut1 downregulation, O-glucosylation sites could be differently affected, generating different responses. As the knockdown of Poglut1 could affect several O-glucosylated targets, it is possible that the alteration of MT phenotypes comes from subsequent modification of non-O-glucosylated extracellular, cell surface or intracellular proteins. Indeed, various proteins such as cadherins, integrins, stabilin-2, or myomaker are known to influence the fusion of MBs (Abmayr and Pavlath, 2012; Park et al., 2016; Gamage et al., 2017) and would deserve particular attention. It is also interesting to note that compositional changes in the extracellular matrix influence cell proliferation and could therefore be involved in the slow proliferation rate observed in Pg- KD cells.
Recently, it has been reported that a human POGLUT1 mutation, named D233E, causes a muscular dystrophy with reduced Notch signaling and SC loss (Servián-Morilla et al., 2016). In D233E samples, Pax7+MyoD+ (proliferating) and Pax7+MyoD− (self-renewing) cells were significantly reduced, whereas Pax7−MyoD+ (differentiating) cells were increased. These results are coherent with the significant reduction of Pax7 and the increase of MyoD expression observed in Pg- KD cell lines. The Pax7/MyoD ratio influences major cell fate decisions in C2C12 cells, i.e MB, RC, and myotube. The decrease of Pax7/MyoD ratio commits cells to a premature myogenic differentiation and leads to an earlier appearance of MT, 24 h before their presence in control cells.
The Pax7/MyoD ratio is under the control of Notch pathway activation. Cleaved NICD through Rbp-Jk transcription factor directly regulates Pax7 expression, whereas MyoD expression is mediated by Hes and Hey family proteins, whose expressions are dependent on NICD/Rbp-Jk control (Olguín and Olwin, 2004). A conditional knockin mouse model, in which endogenous Notch signaling was specifically blocked in muscle SCs, showed a muscular dystrophy and a defect in SC maintenance, due to a reduced Pax7 expression (Lin et al., 2013). This study highlights the role of Notch signaling in skeletal muscle regeneration through Pax7/MRFs expressions in SCs and muscle cells. So, if cleaved NICD amount is too low, it is not sufficient to maintain Pax7 expression and quiescence of SCs, especially because the NICD half-life is only of about 3 h (Buas and Kadesch, 2010). Consequently, we could hypothesize that in Pg- C2C12 cells, Poglut1 knockdown reduces activation of Notch signaling and Pax7 expression, and influences MB commitment in myotube to the detriment of RC, by upregulation of MyoD, at the origin of the entry into differentiation.
Previously, Der Vartanian et al. (2015) showed that Pofut1 transcript and the corresponding protein are expressed at higher levels in RC relative to MT, both in WT and Po- C2C12 cells. These findings describe the same differential expression of Poglut1 between RC and MT in WT and Pg- KD C2C12 cells. In connection with the Poglut1 expression, we found that Notch signaling was more activated in RC than in MT, which could reveal a modulator role of Poglut1 according to cell types, in the context of myogenic differentiation.
Conclusion
Our findings demonstrate an essential role of Poglut1 in the context of skeletal myogenesis. The expression of Poglut1 is necessary for proper Notch signaling and maintenance of a Pax7/MyoD ratio compatible with a coordinated differentiation process. A strong reduction in Poglut1 expression decreases Pax7/MyoD balance and induces a premature commitment of cells in myogenic differentiation.
Footnotes
Acknowledgments
This work was supported by a French Ministry of Higher Education and Research doctoral fellowship to M.P. The authors thank Dr. Daniel Petit for his help in statistics analyses.
Authors' Contributions
M.P. performed experiments and analyzed the data. A.G. and A.M. designed the research study and interpreted the data. M.P. and A.D.V. carried out Pg- KD cell line constructions. M.P., A.G., and A.M. drafted and critically revised the article. All the authors have corrected and approved the article for publication.
Disclosure Statement
No competing financial interests exist.