
Abstract
CYP24A1 is overexpressed in colorectal cancer, and the reason for the dysregulation of CYP24A1 in colorectal cancer is still unknown. In the present study, experiments were designed to test whether CYP24A1 inhibition facilitated the antiproliferative effect of 1,25(OH)2D3. In addition, the role of methylation in the regulation of CYP24A1 expression in human colorectal cancer was investigated. The expression of CYP24A1 in SW480 and Caco2 colorectal cancer cells was inhibited by RNAi. CYP24A1 inhibition significantly increased the antiproliferative effects of 1,25(OH)2D3 in SW480 cells compared with 1,25(OH)2D3 treatment alone (16.78% ± 2.08% vs. 33.53% ± 2.47%, p < 0.05). In addition, CYP24A1 inhibition sensitized Caco2 cells to 1,25(OH)2D3. We also found that CYP24A1 inhibition induced β-catenin to translocate from the nucleus to the plasma membrane in SW480 cells and enhanced the inhibitory effect of 1,25(OH)2D3 on C-myc. Furthermore, CYP24A1 mRNA expression in Caco2 cells was increased after demethylation treatment, and the expression of CYP24A1 induced by 1,25(OH)2D3 was significantly higher in cells treated with 5-aza-2′-deoxycytidine (DAC) than in an untreated group. In conclusion, inhibition of CYP24A1 expression enhances the antitumor effect of 1,25(OH)2D3 in colorectal cancer, and DNA methylation is involved in the regulation of CYP24A1 expression in a cell-dependent manner.
Introduction
V
24-Hydroxylase, encoded by the CYP24A1 gene, is the rate-limiting enzyme that catabolizes 25(OH)D3 and 1,25(OH)2D3 to the less active 24,25(OH)2D3 and 1α,24,25(OH)3D3, respectively, and eventually to calcitroic acid. CYP24A1 plays an important regulatory role in the antitumor effect of 1,25(OH)2D3. Studies have shown that CYP24A1 is a potential oncogene; aberrant expression of CYP24A1 in prostate cancer, lung cancer, and esophageal cancer is closely related to tumor malignancy and poor prognosis (Mimori et al., 2004; Chen et al., 2011; Tannour-Louet et al., 2014). We have found that CYP24A1 is also overexpressed in colorectal cancer tissues, and colorectal cancer patients with high CYP24A1 expression showed significantly reduced overall survival and disease-free survival compared with patients with low CYP24A1 expression in a previous study (Sun et al., 2016). The underlying mechanism might be that the elevated expression of CYP24A1 accelerates inactivation of 1,25(OH)2D3, thus inhibiting specific signaling pathways that limit the growth of colorectal cancer.
However, the reason for the dysregulation of CYP24A1 in colorectal cancer is still unclear. The presence of a CpG island in CYP24A1 promoter region (bases −470 to +1263) suggests that DNA methylation may be involved in the dysregulation of CYP24A1 gene expression (Fetahu et al., 2014). The results of an in vitro reporter assay indicated that promoter methylation downregulated basal transcription and abolished 1,25(OH)2D3-mediated activation. In prostate cancer, promoter hypermethylation resulted in CYP24A1 downregulation (Novakovic et al., 2009; Luo et al., 2010). However, little is known about the epigenetic mechanism that regulates CYP24A1 expression in colorectal cancer.
Since CYP24A1 is abnormally expressed in colorectal cancer, and it is important for the regulation of 1,25(OH)2D3 levels, experiments were designed to test whether CYP24A1 promotes colorectal cancer growth by inhibiting 1,25(OH)2D3-mediated transcriptional regulation in the present study. In addition, we investigated the role of methylation in the regulation of CYP24A1 expression in human colorectal cancer.
Materials and Methods
Cell lines and tissue samples
Human colorectal cancer cell lines, SW480 and Caco2, were obtained from American Type Culture Collection and maintained in our laboratory. Both cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco, Thermo Fisher Scientific, Massachusetts, MA) supplemented with 10% fetal bovine serum (Thermo Fisher Scientific) and 100 U/mL penicillin–streptomycin at 37°C in a humidified atmosphere containing 5% CO2.
Five paired human fresh colorectal cancer tissues and matched normal tissues were obtained from the tissue bank in the China–Japan Union Hospital of Jilin University. The present study was approved by the Ethics Committee of the China–Japan Union Hospital of Jilin University (No. 2016-nsfc014).
CYP24A1-specific siRNA and transfection
Three specific siRNA sequences were designed to silence CYP24A1 expression; the sequences were as follows (5′-3′): siRNA-1-sense: GCUGCAGAUUCUCUGGAAATT, siRNA-1-anti-sense: UUUCCAGAGAAUCUGCAGCTT; siRNA-2-sense: GCCUCGUGUUGUAUGAGAATT, siRNA-2-anti-sense: UUCUCAUACAACACGAGGCTT; siRNA-3-sense: GCAGAAGAUUUGAGGAAUATT, siRNA-3-anti-sense: UAUUCCUCAAAUCUUCUGCTT, negative control-sense: UUCUCCGAACGUGUCACGUTT, and negative control anti-sense: ACGUGACACGUUCGGAGAATT. SW480 and Caco2 cells (2 × 105/well) were seeded into six-well plates in DMEM complete growth medium without antibiotics. When the cell confluency reached ∼50%, cells were transfected with CYP24A1 siRNA or negative control siRNA using Lipofectamine2000 Transfection reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. Forty-eight hours after transfection, cells were rinsed with PBS and collected, and then RNA was extracted by TRIzol reagent (Invitrogen) to determine which siRNA had the best interference effect and detect the expression of target genes.
Cell proliferation assays
Cell proliferation was assessed by Cell Counting Kit-8 (CCK8; KeyGen, Nanjing, China). At 24 h post siRNA transfection, cells were treated with vehicle (0.1% ethanol) or 100 nM 1,25(OH)2D3 (Sigma-Aldrich, St. Louis, MO) for 2 days. Medium was changed and, 10 μL of CCK8 reagent was added to each well. The plates were measured in a microplate reader (Bio-Tek) at 450 nm after 2 h incubation at 37°C.
Quantitative real-time PCR
Using the GoScript Reverse Transcription System Kit (A5000; Promega), 500 ng RNA extracted from different cell lines and tissues was converted into cDNA in accordance with the manufacturer's instructions. mRNA expression levels were quantified by qPCR using the SYBR Premix Ex Taq II (TaKaRa Bio, Otsu, Shiga, Japan) with the following primers: CYP24A1: forward primer 5′-GCCGTATTTAAAAGCCTGTCTGAA-3′ and reverse primer 5′-ACCTGGGTATTTAGCATGAGCACTG-3′. C-myc: forward primer 5′-GGCTCCTGGCAAAAGGTCA-3′ and reverse primer 5′-CTGCGTAGTTGTGCTGATGT-3′. Cyclin D1: forward primer 5′-GTGTATCGAGAGGCCAAAGG-3′, and reverse primer 5′-GCAACCAGAAATGCACAGAC-3′. GAPDH: forward primer 5′-AGAAGGCTGGGGCTCATTTG-3′ and reverse primer 5′-AGGGGCCATCCACAGTCTTC-3′. β-Actin: forward primer 5′-CATGTACGTTGCTATCCAGGC-3′ and reverse primer 5′-CTCCTTAATGTCACGCACGAT-3′.
The reaction was performed with the Eppendorf Mastercycler ep Realplex (initial incubation at 95°C for 30 s followed by 40 cycles of 95°C for 5 s, 60°C for 30 s and melting curve). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and β-actin mRNA levels were used to standardize the measurements of target genes. Relative mRNA levels were assessed by the comparative 2−ΔΔCt method.
Western blot
The protein extractions from SW480 and Caco2 cells were isolated using RIPA Lysis Buffer (P0013B; Beyotime, China); nuclear proteins were extracted with the Nuclear Protein Extraction Kit (R0050; Solarbio, China). Then, 50–100 μg protein was separated by 12% SDS-polyacrylamide gel electrophoresis and electroblotted onto polyvinylidene difluoride membranes (Millipore). After blocking with 5% fat-free dry milk in PBS, the blots were incubated overnight at 4°C with primary antibodies, including anti-CYP24A1 (ab175976, 1:300; Abcam), anti-VDR (ab3508, 1:100; Abcam), anti-β-catenin (D10A8, 1:1000; CST, Danvers, MA), and anti-C-myc (D3 N8F, 1:1000; CST); anti-GAPDH (D16H11, 1:1000; CST) and anti-TBP (D5C9H, 1:1000; CST) antibodies were used as internal controls for the total proteins and nuclear proteins, respectively. Then, the membranes were incubated with horseradish peroxidase-conjugated secondary antibody for 1 h at room temperature, and protein bands were visualized using an Odyssey infrared imaging system (Gene Company).
Immunostaining
SW480 cells were transfected with CYP24A1 siRNA or negative control siRNA, and 100 nM 1,25(OH)2D3 was added at 24 h after transfection. Twenty-four hours after 1,25(OH)2D3 treatment, cells were rinsed three times with PBS and then fixed in cold 4% paraformaldehyde for 20 min. After the cells were washed with PBS three times, nonspecific staining was blocked by incubation with 10% normal goat serum for 30 min at room temperature. Cells were then incubated overnight at 4°C with the primary antibody diluted at 1:100 (D10A8; CST). After three washes with PBST (PBS containing 0.1% Tween), cells were incubated with TRITC AffiniPure secondary antibodies (E031320; EARTHOX, San Francisco, CA) for 1 h at 37°C and rinsed three times with PBST; then, DAPI was added for 5 min at room temperature to stain nuclei. After the cells were washed three times with PBST, the Operetta High Content Screening System (PerkinElmer) was used to observe cell staining and obtain photographs. Images were analyzed using Columbus 2.2 software (PerkinElmer).
Bisulfite DNA sequencing
SW480 and Caco2 cells were seeded at 1 × 105 cells/well in a six-well plate overnight, and then cells were treated with 5-aza-2′-deoxycytidine (DAC) at 0.5, 1.0, and 2.0 μM for 3 consecutive days, followed by the addition of 1,25(OH)2D3 (100 nM) for an additional 12 h.
Genomic DNA extracted from colorectal cells and tissues using a Genomic DNA Purification Kit (A1120; Promega, Madison, WI) was treated with bisulfite with a Bisulfite Conversion Kit (No. 55016; Active Motif, Carlsbad, CA). PCR amplification was performed using EpiTaq™HS (TaKaRa Bio); primers were designed by MethPrimer and were as follows (Li and Dahiya, 2002): forward: 5′-AAAATTATTTTAGTTTAGGTTGGGG-3′, reverse: 5′-AAATAACCCCCAAAA A ATCATAC-3′. PCR products were purified using a DNA Gel Extraction Kit (Sangon, Shanghai, China) and cloned into the pUCm-T vector using the TA Cloning Kit (Sangon, Shanghai, China) for sequencing. A total of 5 clones from each sample were sequenced, and results were analyzed with BiQ Analyzer software. The methylation status of each CpG site was evaluated by the ratio of C (methylated) to T (unmethylated).
Statistical analysis
The differences in mRNA expression and cell proliferation between groups were assessed by independent sample t-test or Mann–Whitney U-test (two-tailed); t-test was used when the data were normally distributed; otherwise, Mann–Whitney U-test was used. The correlation between CYP24A1 mRNA expression and the methylation level was examined using Spearman correlation coefficients by rank test. Statistical analysis was performed using SPSS18.0 (SPSS, Chicago, IL); a p-value less than 0.05 was considered statistically significant.
Results
Inhibition of CYP24A1 expression on the antiproliferative effect of 1,25(OH)2D3
Three specific siRNA sequences were designed to inhibit CYP24A1 expression, and Caco2 and SW480 cells were transfected with the three siRNAs at a concentration of 50 nM. siRNA1, siRNA2, and siRNA3 transfection reduced CYP24A1 expression by 57.0%, 24.7%, and 88.3%, respectively, in Caco2 cells compared with transfection with negative control. However, an inhibitory effect of siRNA1 and siRNA2 was not found in SW480 cells; only siRNA3 transfection reduced CYP24A1 expression by 81.6% in SW480 cells (Fig. 1A). Therefore, siRNA3 was selected for subsequent experiments.
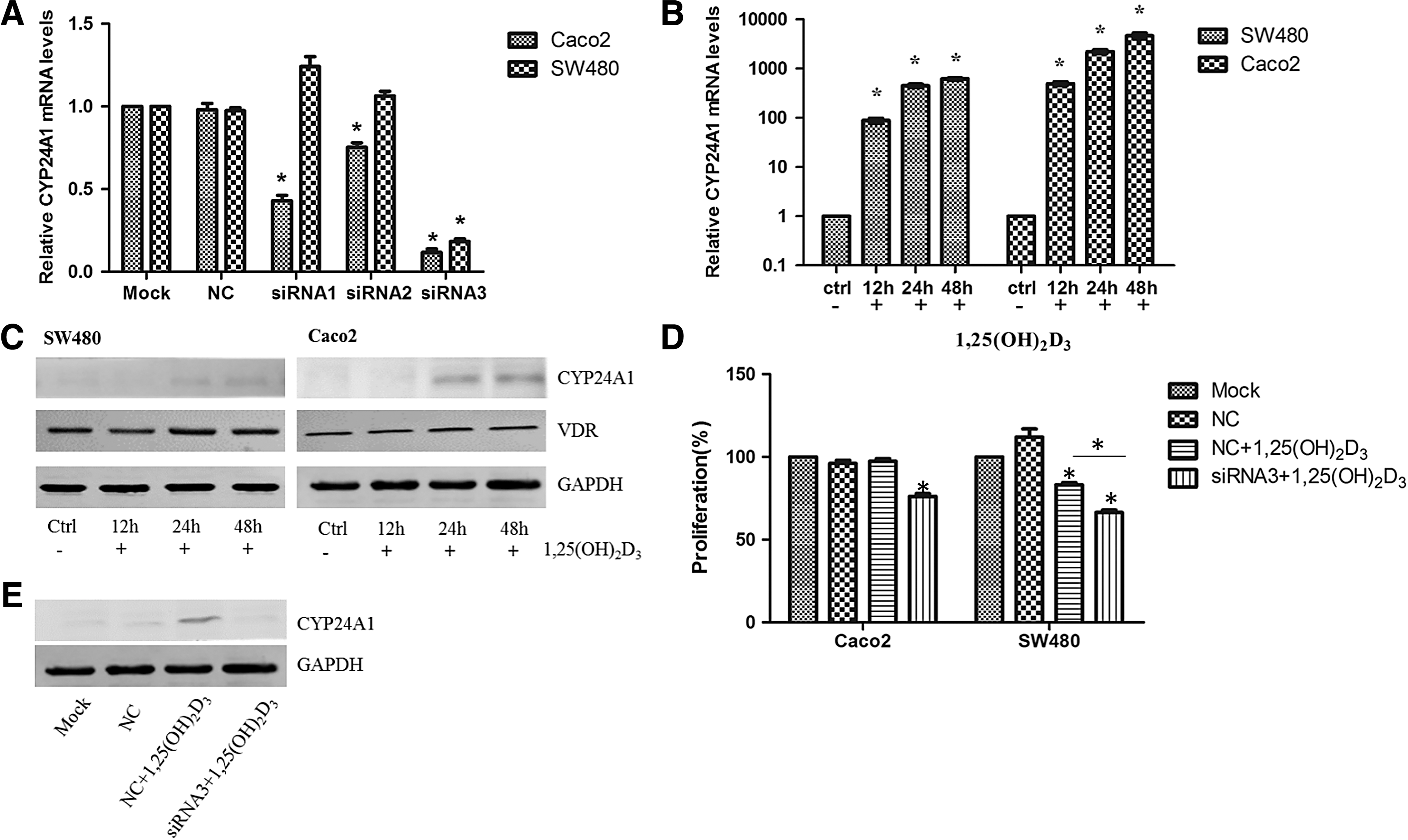
Impact of CYP24A1 inhibition on the antiproliferative effect of 1,25(OH)2D3 in colorectal cells.
To examine the effects of CYP24A1 inhibition on colorectal cell proliferation, CCK8 assays were conducted. When 100 nM 1,25(OH)2D3 was used alone, SW480 cell growth was inhibited by 16.78%, whereas Caco2 cells were insensitive to 1,25(OH)2D3 treatment. Expression of CYP24A1 and VDR in SW480 and Caco2 cells was detected at 12, 24, and 48 h after 100 nM 1,25(OH)2D3 treatment (Fig. 1B, C). 1,25(OH)2D3 induced CYP24A1 mRNA expression 12 h after treatment, and CYP24A1 protein expression also increased 24 h after treatment in both cell lines. However, VDR protein level remained stable upon 1,25(OH)2D3 treatment. Moreover, the increasing trend of CYP24A1 in Caco2 cells was more obvious according to western blot analysis. CYP24A1 siRNA significantly increased the antiproliferative effects of 1,25(OH)2D3 in SW480 cells compared with 1,25(OH)2D3 treatment alone (16.78% ± 2.08% vs. 33.53% ± 2.47%, p < 0.05). In addition, CYP24A1 inhibition sensitized Caco2 cells to 1,25(OH)2D3.
An inhibitory effect of 1,25(OH)2D3 was observed after CYP24A1 expression was inhibited by siRNA in Caco2 cells; the average proliferation rate of the Caco2 cells decreased to 76.23%, which was significantly lower than that of the control group (Fig. 1D). To further confirm that the inhibition of proliferation was associated with the interference of CYP24A1, protein levels of CYP24A1 were evaluated by western blot in different treatment groups, and the results showed that the protein level in the interference group was significantly lower than that in the noninterference group upon 1,25(OH)2D3 treatment (Fig. 1E).
CYP24A1 inhibition promotes 1,25(OH)2D3-mediated β-catenin nuclear export and reduces its target gene expression
Studies have demonstrated that 1,25(OH)2D3 inhibits the WNT/β-catenin pathway, which is often aberrantly activated in colorectal cancer, in colorectal cancer cells, thereby inhibiting cell proliferation and preserving the differentiated phenotype (Palmer et al., 2001; Larriba et al., 2013). We tested the effects of CYP24A1 inhibition on β-catenin nuclear export. Immunostaining results indicated that CYP24A1 inhibition induced β-catenin translocation from the nucleus to the plasma membrane in SW480 cells (Fig. 2A). CYP24A1 inhibition significantly reduced β-catenin nuclear fluorescence intensity (Fig. 2B). Nuclear β-catenin expression was also detected by western blot; results showed the same tendency with immunostaining (Fig. 2C). 1,25(OH)2D3 inhibited expression of C-myc, an important downstream gene of the Wnt/β-catenin signaling pathway, and this inhibitory effect was more obvious after CYP24A1 inhibition in SW480 cells (Fig. 2D, E). We also detected the expression of Cyclin D1; however, expression was not downregulated after CYP24A1 inhibition (data not shown).
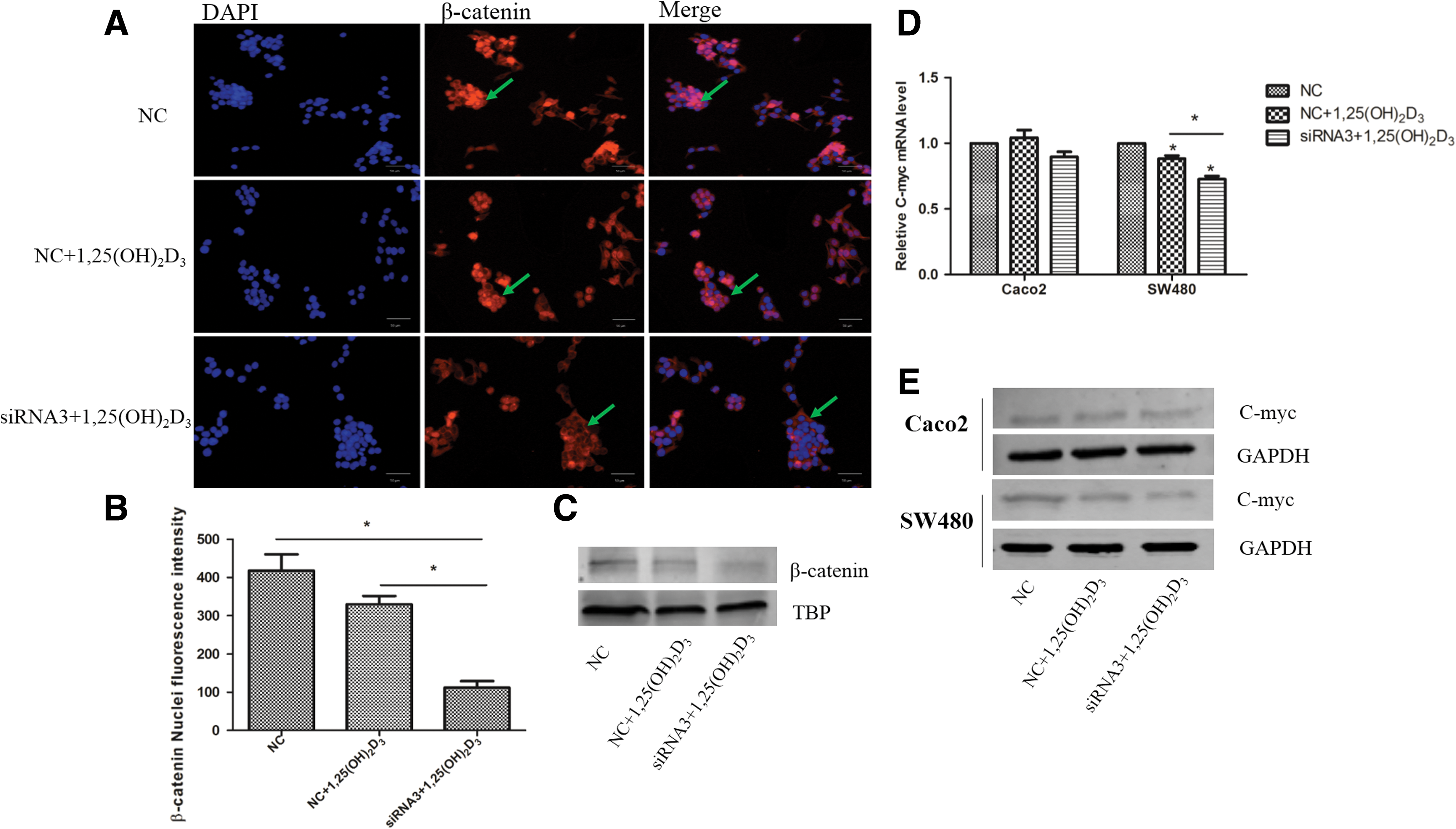
The effect of CYP24A1 inhibition on β-catenin nuclear export and transcriptional activity.
Activation of CYP24A1 expression by DAC in colorectal cancer cells
To investigate the role of methylation in the regulation of CYP24A1 expression, we treated SW480 and Caco2 cells with 0.5, 1.0, or 2.0 μM DAC; CYP24A1 expression was then detected by RT-PCR. The results showed that there was no significant difference in CYP24A1 mRNA expression among the three groups in SW480 cells. However, in Caco2 cells, treatment with different doses of DAC induced CYP24A1 expression in each group compared with the control treatment, and no difference in expression was found between the groups treated with 1.0 and 2.0 μM DAC (Fig. 3A). To confirm the demethylation effect of DAC, bisulfite DNA sequencing was done on Caco2 and SW480 cells. The region analyzed by bisulfite sequencing is localized at the CYP24A1 promoter region (−499 to −244), which contains 29 CpGs (Fig. 3B). We found that promoter methylation of CYP24A1 in Caco2 cells was reduced from 75.2% to 11.7% (Fig. 3C, D) and from 62.8% to 13.1% in SW480 cells (Fig. 3E, F).
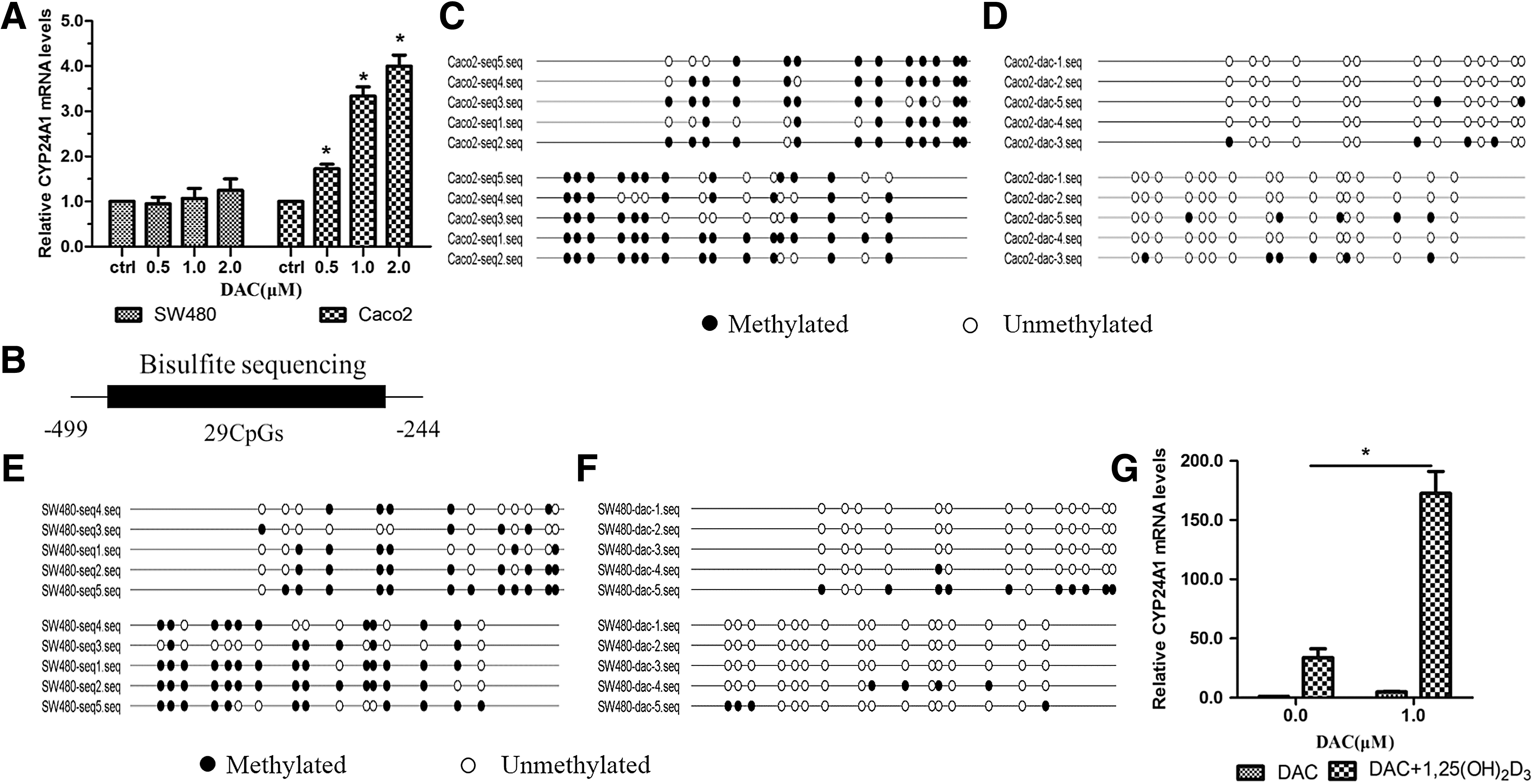
Activation of CYP24A1 expression by DAC in colorectal cancer cells.
We further investigated whether the methylation status of the CYP24A1 promoter region affected 1,25(OH)2D3-induced CYP24A1 expression in colorectal cancer cells. Caco2 cells were treated or not treated with DAC before 1,25(OH)2D3 was added, and qRT-PCR revealed that the expression of CYP24A1 induced by 1,25(OH)2D3 was significantly higher in cells treated with DAC than in cells not treated with DAC; the difference was statistically significant (Fig. 3G).
Correlation of CYP24A1 expression in human colorectal tissues with DNA promoter methylation status
We analyzed CYP24A1 expression in five matched human colorectal cancer and adjacent colorectal tissues. qRT-PCR indicated that CYP24A1 expression was significantly increased in colorectal malignant tissues compared with its expression in normal tissues (Fig. 4A, C). Bisulfite DNA sequencing revealed that the methylation level of the CYP24A1 promoter region was 27.3%, 16.2%, 8.6%, 0.0%, and 53.9% in the five colorectal cancer tissues; the median methylation level was 21.2%. Meanwhile, the methylation level was 36.0% 26.5%, 42.5%, 0.0%, and 40.0% in the five matched normal tissues, and the median methylation level was 29.0% (Fig. 4B). There was no significant difference between the levels of methylation in colorectal cancer tissue and adjacent mucosal tissue (Fig. 4D). In addition, CYP24A1 mRNA expression did not correlate with its promoter DNA methylation status (Fig. 4E).
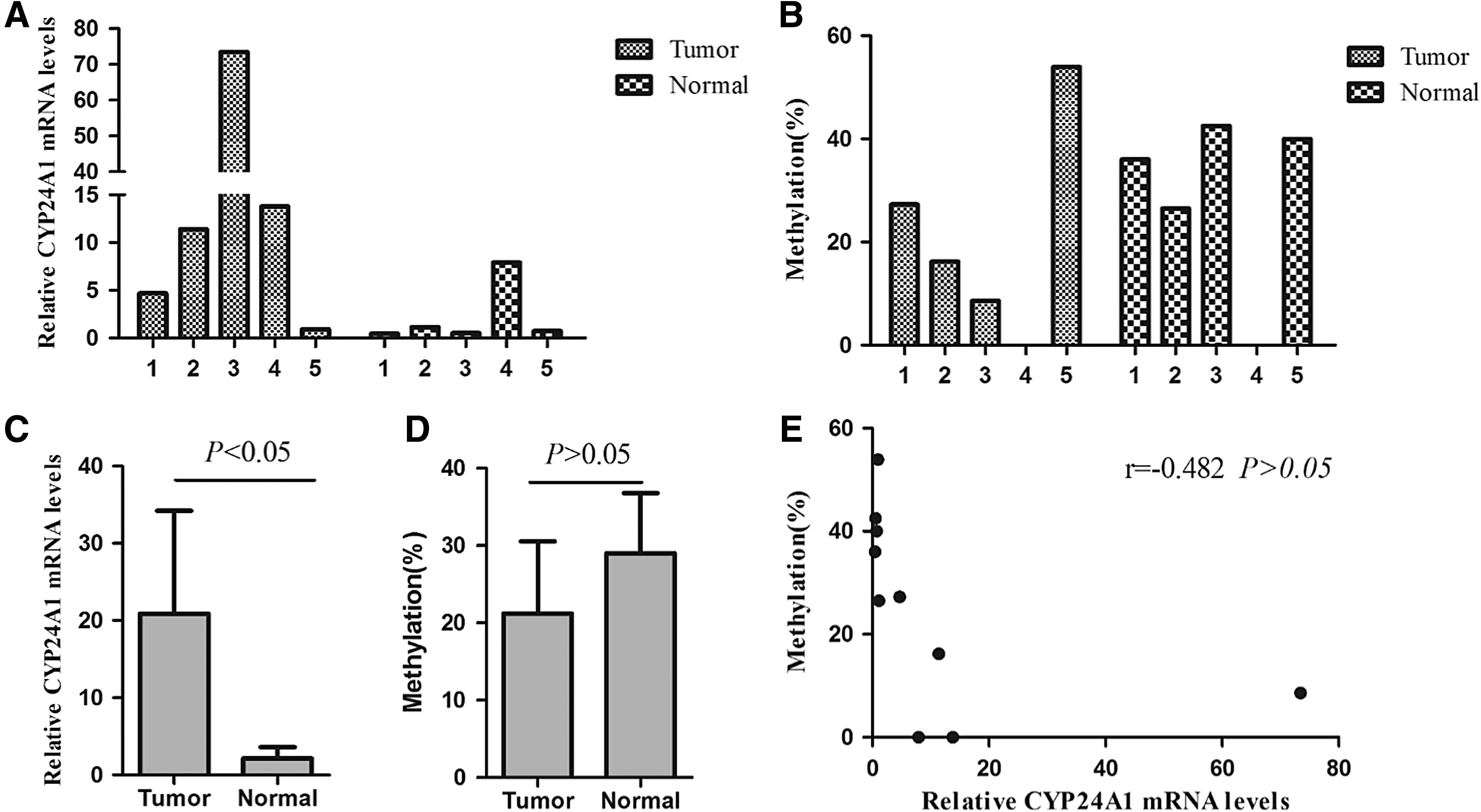
Correlation of CYP24A1 expression in human colorectal tissues with promoter DNA methylation status.
Discussion
As a key vitamin D3 catabolizing enzyme, CYP24A1 is aberrantly increased in several cancer cell types and is associated with cancer development and progression (Friedrich et al., 2003; Mimori et al., 2004; Mitschele et al., 2004; Cross et al., 2005; Parise et al., 2006; Tannour-Louet et al., 2014). Studies have shown that CYP24A1 is a potential oncogene. Aberrant expression of CYP24A1 is closely related to inflammatory bowel disease and colorectal cancer (Cross et al., 2011). Horváth et al. (2010) compared CYP24A1 expression in normal human colon mucosa, colorectal adenomas, and adenocarcinomas, and the results showed that CYP24A1 was a potential biomarker for colorectal tumorigenesis. We further investigated the prognostic value of the expression of CYP24A1 in colorectal cancer patients in a previous study and found that CYP24A1 was a prognostic biomarker of human colorectal carcinoma (Sun et al., 2016). Höbaus et al. (2016) demonstrated that aggressiveness and the proliferative potential of colorectal tumors were higher in CYP24A1-overexpressing xenografts than in controls. We hypothesized the elevated intratumoral levels of CYP24A1 may accelerate inactivation of 1,25(OH)2D3, thus abrogating the biological actions of 1,25(OH)2D3 in inhibiting specific signaling pathways that promote the progression of colorectal cancer.
The results of other reports using CYP24 inhibitors showed that inhibition of CYP24A1 expression enhanced the antitumor effect of 1,25(OH)2D3 in prostate cancer cells, lung cancer cells, and colon cancer cells (Zhang et al., 2012; Kósa et al., 2013; Luo et al., 2013). Combination of 1,25(OH)2D3 with ketoconazole, a nonselective azole CYP24A1 inhibitor, increases the half-life of 1,25(OH)2D3 and enhances its antiproliferative effects in PC3 prostate cancer cells (Muindi et al., 2010). The CYP24-selective inhibitor CTA091 increased 1,25(OH)2D3 stability and 1,25(OH)2D3-mediated growth inhibition in H292 lung cancer cells (Zhang et al., 2012). Kósa et al. (2013) investigated 13 tetralones to screen out a compound that is effective in CYP24A1 inhibition. The proliferation of Caco2 cells was not inhibited when 1,25(OH)2D3 was used alone, but the cell viability and proliferation were significantly decreased when used in combination with the screened inhibitor KD-35 (Kósa et al., 2013). We also found that Caco2 cells were insensitive to 1,25(OH)2D3 in the present study. The reason for this insensitivity might be that 1,25(OH)2D3 induced the expression of CYP24A1 in a time and dose-dependent manner due to a negative feedback mechanism in Caco2 colorectal cancer cells; thus, the amount of 1,25(OH)2D3 in local tumor cells is too limited to exert an antitumor effect.
The CYP24A1 inhibitors that have been published on to date are azole and non-azole inhibitors or vitamin D analogs, which may affect the other enzymes of cytochrome P450, such as CYP27B1 (Luo et al., 2013). In this study, siRNA was designed to specifically inhibit expression of CYP24A1, and CYP24A1 interference sensitized Caco2 colorectal cancer cells to D3, which was consistent with the results in Kósa's study. CYP24A1 inhibition also enhanced the antitumor effect of 1,25(OH)2D3 in SW480 colorectal cancer cells.
Aberrant activation of WNT/β-catenin signaling, followed by activation of target genes, is the most common alteration in sporadic colorectal cancer. Studies have demonstrated that 1,25(OH)2D3 inhibits the WNT/β-catenin pathway and β-catenin transcriptional activity in colorectal cancer cells (Pereira et al., 2012). We tested whether CYP24A1 inhibition enhanced the inhibitory effect of 1,25(OH)2D3 on the WNT/β-catenin pathway. Our results showed that CYP24A1 inhibition induced β-catenin to translocate from the nucleus to plasma membrane in SW480 cells. In addition, the expression of C-myc was reduced by CYP24A1 inhibition compared with 1,25(OH)2D3 alone. Therefore, CYP24A1 inhibition might enhance the inhibitory effect of 1,25(OH)2D3 through mediating the actions of the WNT/β-catenin pathway.
The mechanism underlying the overexpression of CYP24A1 in colorectal cancer is not clear; the expression of VDR is actually reduced in colorectal cancer tissues (Horvath et al., 2010). Furthermore, 1,25(OH)2D3 induced CYP24A1 expression in SW480 and Caco2 colorectal cancer cells, whereas VDR levels remained stable, which indicates that the high CYP24A1 level is not necessarily the result of the identified physiological transcriptional process in which 1,25(OH)2D3 binds to VDR to activate CYP24A1. Studies have shown that multiple factors might be associated with abnormal regulation of CYP24A1 expression in cancer, including amplification at the CYP24A1 locus, dysregulation of miRNAs, and methylation status of the CYP24A1 promoter (Albertson et al., 2000; Komagata et al., 2009; Fetahu et al., 2014).
A classical 5′ CpG island spans the CYP24A1 promoter, containing two vitamin D response elements (VDRE) and SP1-binding sites that regulate 1,25(OH)2D3-induced and basal transcription, indicates a role for DNA methylation-based regulation. In this study, we treated SW480 and Caco2 cells with demethylating agents and found that CYP24A1 expression was induced in Caco2 cells but not in SW480 cells. In the study of Höbaus et al. (2013), DAC treatment increased the expression of CYP24A1 mRNA by 50-fold in Coga1A and HT-29 colon cancer cells, whereas the expression in Coga13 and Caco2/AQ cells did not change significantly, potentially due to different promoter methylation status in these types of cells. Overall methylation of the CYP24A1 promoter in Caco2 cells was 75.2% and was reduced to 11.7% after DAC treatment.
We further analyzed whether the methylation status of CYP24A1 promoter region affected 1,25(OH)2D3-induced CYP24A1 expression in colorectal cancer cells. The expression of CYP24A1 induced by 1,25(OH)2D3 was significantly higher in cells treated with DAC than in the untreated group. Through promoter methylation analysis, we found demethylation of CpG in the VDRE region after DAC treatment; however, more studies are needed to identify whether hypermethylation of the CYP24A1 promoter prevents VDR from binding to the VDREs and inhibits the transcription of CYP24A1.
Promoter hypermethylation might result in low expression of CYP24A1 in the normal colon, and hypomethylation increases CYP24A1 expression during carcinogenesis. Höbaus et al. (2013) first reported that there was no significant difference between the levels of methylation in colorectal cancer tissue and adjacent mucosal tissue. Moreover, the expression of CYP24A1 did not correlate with the level of methylation of its promoter (Höbaus et al., 2013), and the results of our present study are consistent with this research. Therefore, DNA methylation does not fully account for the difference in CYP24A1 expression in human colorectal cancer and normal tissues. Histone modifications might also play a role in CYP24A1 expression, according to a previous study (Luo et al., 2010). The mechanisms of CYP24A1 abnormal expression in colorectal cancer need to be further elucidated.
In conclusion, inhibition of CYP24A1 expression enhances the antitumor activity of 1,25(OH)2D3 in colorectal cancer, and DNA methylation is involved in the regulation of CYP24A1 expression in a cell-dependent manner. Accumulating data demonstrates that 1,25(OH)2D3 plays an important role in inhibition of tumor growth in colorectal cancer. However, the antitumor effects of 1,25(OH)2D3 have been limited in clinical trials. One possible reason for this limited effect might be the frequent overexpression of CYP24A1 in tumors, which accelerates inactivation of 1,25(OH)2D3 in local tumor tissue. Elucidating the underlying mechanisms of CYP24A1 aberrant expression will provide new strategies for inhibiting CYP24A1 expression. Many preclinical studies have demonstrated that combining 1,25(OH)2D3 with a selective CYP24A1 inhibitor could improve the efficacy of 1,25(OH)2D3 in cancer treatment (Swami et al., 2005; Wang et al., 2012). However, the efficient application of 1,25(OH)2D3 together with CYP24A1 inhibitors in the clinical treatment of colorectal cancer while avoiding hypercalcemia requires further investigation.
Footnotes
Acknowledgment
This study was supported by the National Nature Scientific Foundation of China (Nos. 81472209 and 81602715).
Disclosure Statement
No competing financial interests exist.