
Abstract
Fibronectin type III domain-containing 5 protein (Fndc5) is responsible for producing a secretory protein termed, “irisin.” A modified expression of Fndc5 has been reported in different tissues during development, differentiation processes, as well as in metabolic events such as exercise. One of the important issues to be fixed is whether Fndc5 RNA level and protein content are concerted and modified hand in hand. Therefore, the aim of this study is to assess Fndc5 RNA and protein levels in various tissues of mouse and rat with emphasis on brain. Biopsies from various parts of neonatal and adult mouse and rat tissues were simultaneously assessed for transcript levels of Fndc5 and compared with the respective protein levels at the same time. Data indicated, unlike in muscle, no concerted fluctuations were observed for Fndc5 RNA and protein, especially in brain. Further look at four regions of brain (cerebellum, putamen, hippocampus, and cortex) revealed a similar discrepant expression. To hypothesize whether such discrepancy is arisen by miRNAs, we selected three main miRNAs, which were predicted to target Fndc5 and their expression levels were assessed in central nervous system (CNS) of mouse and hippocampus of rat. miRNA levels showed an antiparallel correlation with protein level of Fndc5, interpreting a putative role in regulating Fndc5 protein content in CNS. This phenomenon may represent the importance of governing Fndc5 content in neural cells, which seems to be crucial for neural function and differentiation.
Introduction
F
Due to detection of high level of Fndc5 RNA in mouse hippocampus (Wrann et al., 2013), more experiments are needed to identify the expression and function of this protein, especially in the hippocampus. Therefore, at the first step to perform such assessments, we aimed to evaluate the Fndc5 expression pattern at both RNA and protein levels in the brain, with special attention to the hippocampus.
Materials and Methods
Animals
The experiments were conducted according to the guidelines approved by the ethics committees of Royan Institute and Isfahan University of Medical Sciences. Eight-week-old male NMRI mice and Wistar rats were maintained under 12 h light–12 h dark cycle at a relative humidity of 40–60% at 18–23°C. Food and tap water were available ad libitum.
Tissue collection
Adult mice were anesthetized by 75 mg/kg ketamine-HCl and 10 mg/kg xylazine-HCl and their skeletal muscle (gastrocnemius), intra-abdominal visceral fat, brown fat, subcutaneous fat, brain (cerebellum, putamen, hippocampus, and cortex), kidney, liver, lung, skin, and intestine were collected and immediately frozen in liquid nitrogen and stored at −80°C for further analysis. The hippocampus from adult and neonate rats was furthermore dissected. In addition, different brain regions (cerebellum, putamen, hippocampus, and cortex) of 1-day mouse pups were dissected.
RNA isolation and reverse transcription quantitative polymerase chain reaction
Total tissue RNA was isolated by homogenizing the tissue in TRIzol reagent (Ambion, Austin, TX). It was then reverse transcribed to cDNA using random hexamer and the PrimeScript™ RT reagent Kit (TaKaRa BIO, Shiga, Japan). Synthesized cDNA was used for real-time PCR in the total volume of 10 μL containing SYBR Green Master Mix (TaKaRa, Tokyo, Japan). To perform reverse transcription quantitative polymerase chain reaction (RT-qPCR), the specific primers mentioned in Table 1 were used. Primers were designed by Beacon Designer software (Version 7.2) and were obtained from Metabion (Germany). RT-qPCR was carried out in a thermal cycler Rotor gene 6000 (Corbett, Australia). The expression of Fndc5 was assessed related to the housekeeping gene Gapdh according to the 2−ΔΔCT method. The expression level of Fndc5 in different tissues was calibrated to its expression in gastrocnemius tissue. All experiments were performed at least in three biological independent repeats with a technical triplicate in each performance.
r.Fndc5 primer used for rat samples and m.Fndc5 primer used for mouse samples. Gapdh primer was used as a housekeeping gene for both sample types. F and R are referred to forward and reverse primers, respectively. Ta stands for annealing temperature.
Protein isolation and Western blot analysis
For protein isolation, tissues were homogenized in RIPA buffer (200 mM Tris/HCl [pH: 7.4], 130 mM NaCl, 10% [v/v] glycerol, 0.1% [v/v] sodium dodecyl sulfate [SDS], 1% [v/v] Triton X-100, and10 mM MgCl2) with protease inhibitor cocktail (Melford, Ipswich, United Kingdom). Homogenates were centrifuged for 15 min at 14,000 g at 4°C and supernatants were frozen at −80°C. Solubilized protein fractions of tissues (30 μg) were loaded on 12% SDS-polyacrylamide gel electrophoresis (SDS-PAGE). Proteins were then transferred to a polyvinylidene difluoride (PVDF) membrane (Biorad, Hercules, CA). Membranes were first blocked in 10% nonfat dry milk overnight at 4°C and then incubated with primary antibodies, rabbit recombinant monoclonal FNDC5 antibody, 1:3000 (Abcam, Cambridge, MA) and mouse monoclonal anti-glyceraldehyde-3-phosphate dehydrogenase antibody GAPDH, 1:5000 (Cat. No. MAB374, RRID:AB_2107445; Millipore). The blots were then washed and incubated with secondary HRP-conjugated antibodies, goat anti-rabbit IgG-HRP, 1:16,000 (Santa Cruz Biotechnology, Santa Cruz, CA), and polyclonal goat anti mouse immunoglobulins/HRP, 1:5000 (Dako, Cambridge, United Kingdom), respectively. The chemiluminescence reaction was detected using the ECL kit (GE Healthcare, Barcelona, Spain) and then exposed to a Kodak X-OMAT film.
miRNA selection and RT-qPCR
Three different microRNAs were selected based on the criteria that will be elucidated briefly. Based on an intense bibliography on published articles, miR-34a-5p was shown to repress the Fndc5 expression in C2C12 myoblasts (Ge et al., 2017). Therefore, it was selected as a valid microRNA for targeting Fndc5. On the other hand, during a search on the miRNA-related websites such as TargetScan (
Statistical analysis
Statistical analysis was performed by SPSS (Version 23.0) and data are shown as mean ± standard error of the mean (SEM) for each group. Comparisons between the groups were analyzed by Kruskal–Wallis test and between the two groups by Mann–Whitney test, and considered to be significant at p ≤ 0.05.
Results
Fndc5 transcript and protein content assessment in different adult mouse tissues
In this study, we investigated the transcription level of Fndc5 in different tissues of adult mouse. Gastrocnemius muscle was set as “calibrator,” and the fold change in relative expression of Fndc5/Gapdh was calculated for various tissues based on this calibrator. Among tissues studied, the brain presented the highest level of Fndc5 expression (Fig. 1A). The Fndc5 protein expression was observed in brown fat, skin, and stomach, as well as in gastrocnemius muscle, but not in the other examined adult mouse tissues, including the brain containing high level of Fndc5 transcript. The secreted form of Fndc5, irisin, was only detected in brown fat tissue (Fig. 1B). Experiments were conducted at least in triplicate, and their protein content was semiquantified, and mean ± SEM for protein expression of Fndc5 is shown in a graph (Fig. 1C).
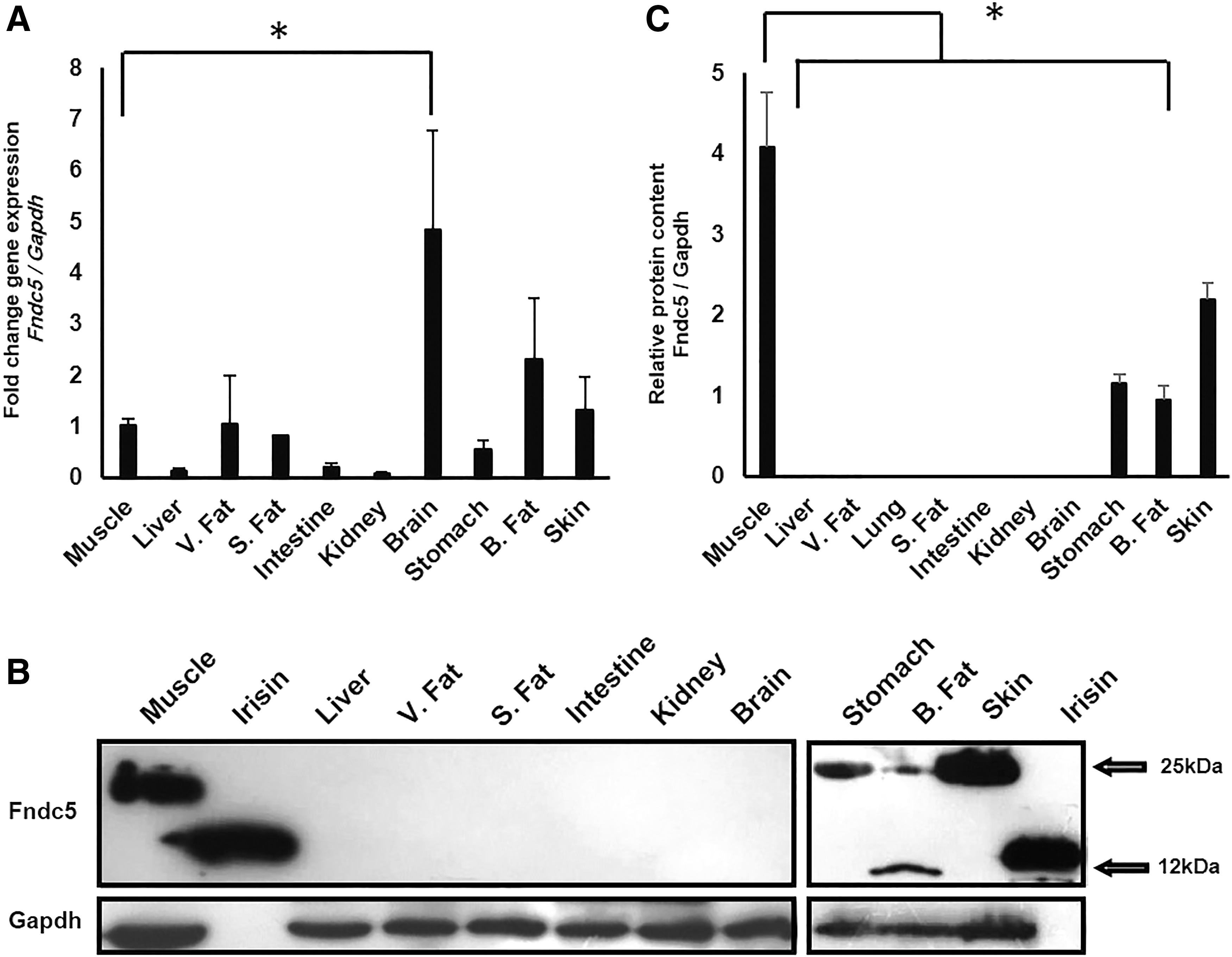
mRNA and protein levels of Fndc5 in different tissues of adult mice.
A discordance was observed between Fndc5 RNA and protein levels in different parts of mouse brain
Neonatal and adult mice were used to assess the Fndc5 expression pattern in different regions of brain. Setting gastrocnemius muscle as “calibrator,” transcriptional levels of Fndc5 were significantly higher in the cerebellum, hippocampus, and putamen of adult mice in comparison to the same respective tissues in neonates (Fig. 2A). On the contrary, Fndc5 protein expression levels were significantly higher in neonates compared to adults (Fig. 2B, C). This observation also holds true when the whole brain of embryo was compared between adult and neonate for RNA and protein content (Fig. 2A, B). It is important to note that no irisin expression band with the expected size of 12 kDa was observed. Similar results were obtained when the hippocampus of neonatal and adult rats was assessed for Fndc5 RNA and protein levels (Fig. 2D–F).
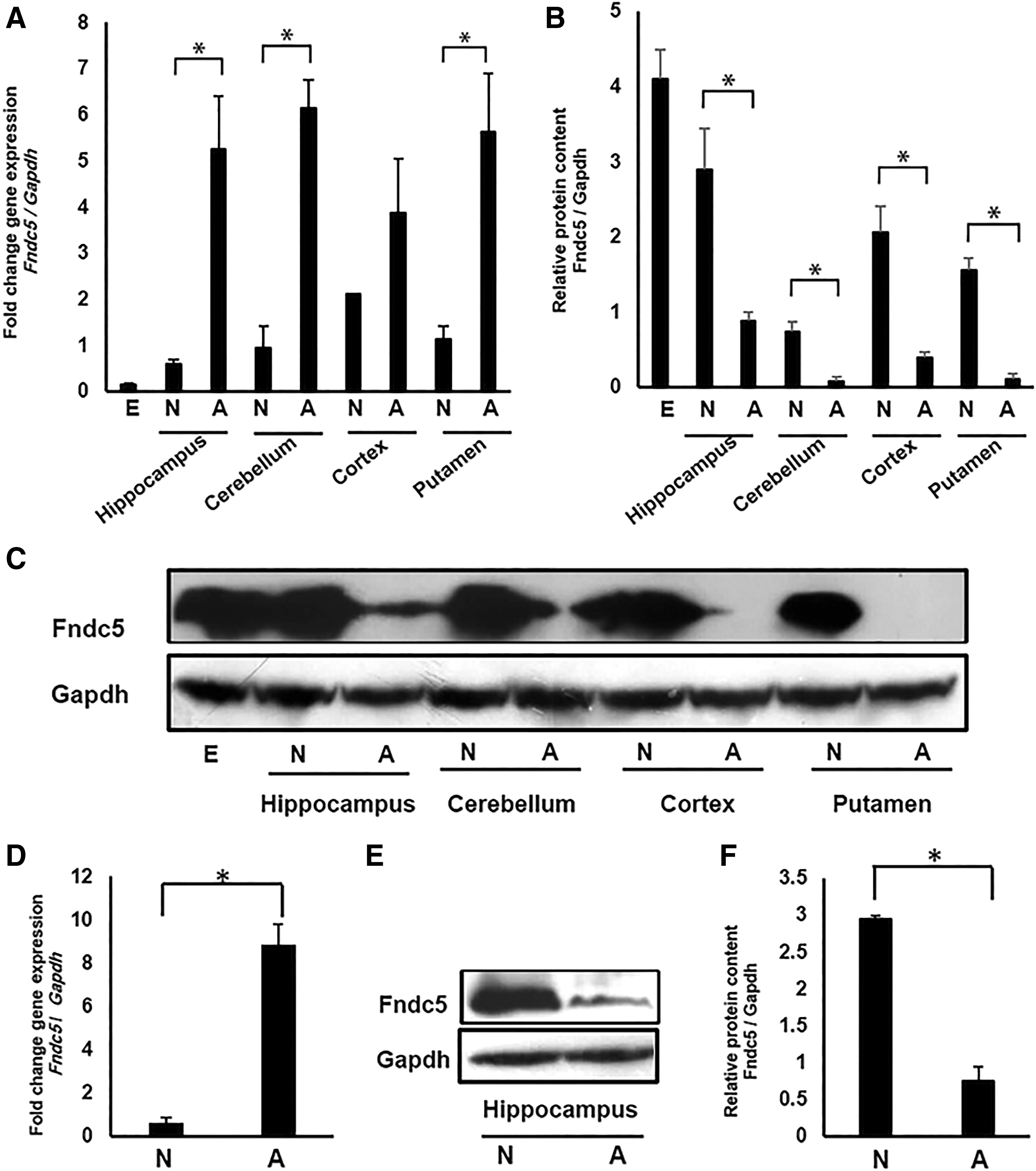
Fndc5 transcript and protein expression pattern in different parts of mouse and rat brains in neonatal and adult stages.
Positive correlation between miRNAs 34a-5p, 129-5p, and 29b-3p levels with Fndc5 expression may suggest a posttranscriptional regulation mechanism for this gene in central nervous system
Discordance between Fndc5 mRNA and protein levels in different brain tissues led us to assess whether alternative mechanisms, including miRNAs, are involved. Computational analyses revealed three candidate microRNAs (miR-34a-5p, miR-129-5p, and miR-29b-3p), which potentially can target Fndc5 transcripts in brain (Table 2). All the three candidate microRNAs showed a significantly higher expression in mouse central nervous system (CNS) and rat adult hippocampus compared to their neonate counterparts (Fig. 3A).
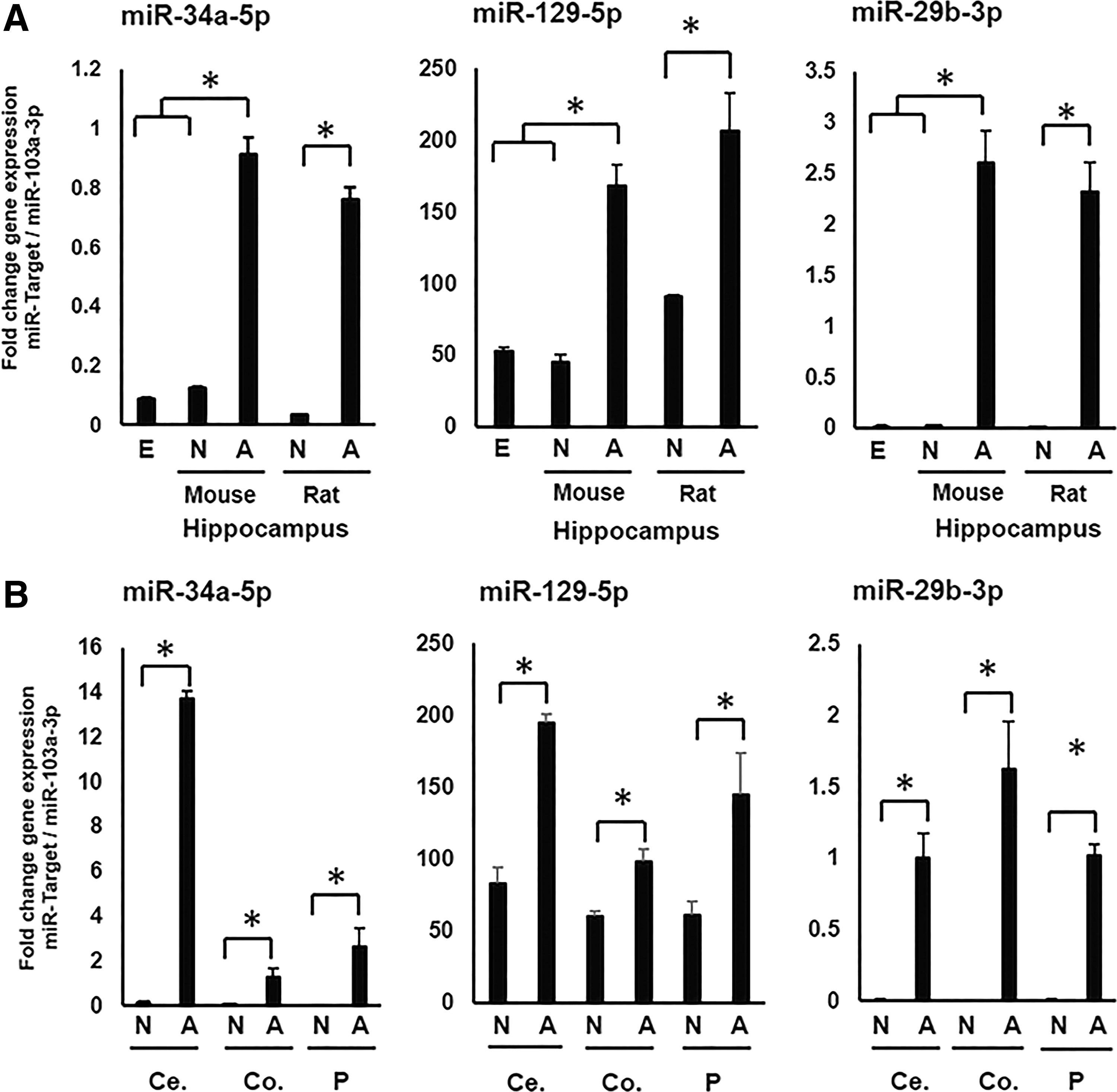
Quantification of miRs 34a-5p, 129-5p, and 29b-3p in different regions of mouse brain and hippocampus of rat, in both neonatal and adult specimens The expression of microRNAs was studied in neonatal and adult hippocampus of mice and rats
Predicted target sequence are represented based on the data obtained in TargetScan.
In addition, we assessed the correlation between Fndc5 protein and three microRNAs, and a negative significant correlation was observed for all the three microRNAs (miR-34a-5p [r = −0.444, p = 0.012], miR-129-5p [r = −0.609, p = 0.000], and miR-29b-3p [r = −0.519, p = 0.003]).
A negative correlation was observed between miR-34a-5p and Fndc5 expression levels in muscle
Assessment of Fndc5 in both neonate and adult rat and mouse muscle samples revealed that the Fndc5 expression was higher in neonates. Concomitant analysis of miRNAs 34a-5p revealed that this miRNA was lower in neonates compared to adult muscle. These differences were shown to be significant in rats (Fig. 4).
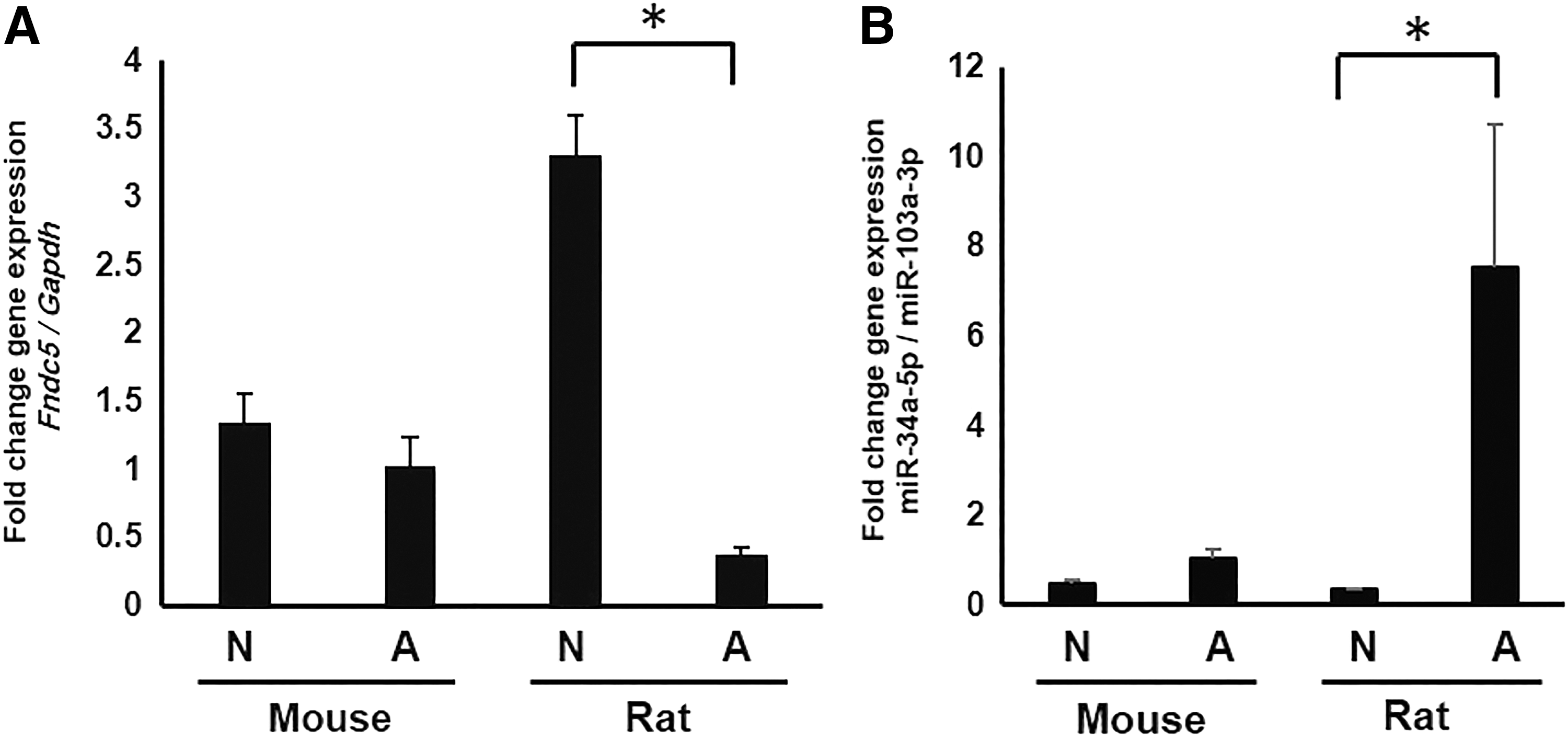
Quantification of Fndc5 and miR-34a-5p in neonatal and adult mouse and rat muscle. The expression of Fndc5
Discussion
Irisin, a cleaved form of Fndc5, is considered myokine and adipokine, regulating metabolism homeostasis through different biological functions, including fat browning and sensitivity to insulin (Roca-Rivada et al., 2013; Schumacher et al., 2013). It also plays a role in numerous physiological processes. The expression of Fndc5 is controlled by (Pgc-1α), which is a coactivator of many transcription factors, including those related to mitochondrial biogenesis (Bostrom et al., 2012; Wrann et al., 2013; Tiano et al., 2015). Previous studies suggested high energy-demanding tissues, such as skeletal muscle, brain, and heart, expressing high level of Pgc-1α and Fndc5 (Kelly, 2012). The expression of Pgc-1α and Fndc5 has been studied during differentiation, and they play an important role in several processes, including cardiogenesis and neurogenesis. During neural differentiation, it has been shown that retinoic acid and bFGF stimulate phosphorylation of ERK, leading to phosphorylation of CREB (cAMP response element binding protein). Phosphorylated CREB increases the expression of Pgc-1α and Fndc5, which are considered to regulate the expression of neurotrophic factors such as Bdnf (Hosseini Farahabadi et al., 2015). Based on these studies, we aimed to assess the relative expression of Fndc5 at RNA and protein levels in different tissues, especially the brain.
Similar to previous studies in humans and mice, our results revealed the highest expression of Fndc5 in mouse brain compared to other tissues. However, to our surprise, the protein expression of Fndc5 was undetectable in mouse brain, while it was relatively high in muscle. It is important to note that based on the high protein expression of Fndc5 in our study and other studies, we decided to assess the expression of Fndc5 RNA in all other tissues relative to muscle (as a calibrator). The discordance between RNA and protein expression guided us to assess Fndc5 expression in different parts of neonatal and adult mouse brain. It was interesting to note that, in contrary to neonates, in adult brain tissue, RNA expression was higher unlike protein expression, which was higher in the neonatal brain. This type of discordance between RNA and protein expression for Fndc5 was not reported previously. Therefore, we also assessed RNA and protein of Fndc5 in neonatal and adult hippocampus of rat brain. Similar results were also observed in rats, indicating that the observed discordance expression of RNA/protein exists across different species. To our knowledge, there are no studies utilizing Western blot analysis to assess the expression of Fndc5 or irisin in brain. In this regard, there are two studies in the literature assessing Fndc5 expression by immunocytochemistry. Dun et al. (2013) showed that Fndc5 was expressed by GAD-positive Purkinje cells of cerebellum. Aydin et al. (2014) showed that Fndc5 was present in some cells of rat brain, and based on the images provided in their study, a limited number of cells expressed Fndc5, and therefore, it is not surprising to see that the amount of protein expression is low in the brain.
The discordance between mRNA and protein levels of Fndc5 between neonatal and adult mice may represent (1) the presence of an inhibitory mechanism suppressing mRNA translation into protein or (2) separate roles for Fndc5 mRNA and protein in various stages of lifespan. To study the former possibility, we aimed to assess the expression of microRNAs related to Fndc5. The first microRNA that we focused on was miR-34a-5p. It has been shown that overexpression or suppression of this microRNA in muscle downregulates and upregulates the Fndc5 expression, respectively (Ge et al., 2017). Myostatin, regulating muscle growth and development, is shown to control the expression of this microRNA (Ge et al., 2017). In accordance with these results, we also observed that when the expression of muscle Fndc5 was high, the expression of miR-34a-5p is low and vice versa. The second microRNA (miR-29b-3p) was selected based on its hippocampal differential expression between neonatal and adult mice, and the third microRNA (miR-129-5p) was selected according to bioinformatics analysis. The expression level of these microRNAs was higher in adults with high RNA and low protein amount of Fndc5, so that it was vice versa in neonatal CNS tissues. Therefore, it is likely that these miRNAs directly or indirectly act as suppressors for Fndc5 mRNA translation into protein in CNS of adult rodents. These observations are also in line with significant negative correlations observed between these microRNAs and Fndc5 protein. It should be noted that miR-34a-5p controlled by myostatin showed an inverse relationship with Fndc5 expression in skeletal tissue being contradictory to our observation in the brain, but concomitant with that in muscle. This may suggest that the expression of Fndc5 is differentially controlled between skeletal and brain tissues.
By studying irisin in different tissues, Aydin et al. (2014) concluded that fat browning or fat burning is locally regulated by irisin, and they stated that in peripheral and CNS tissues, irisin or Fndc5 could act as a “gatekeeper of metabolic energy regulation”. Therefore, it is likely that neonatal brain energy demand will be higher than the adult brain. Higher Fndc5 protein expression in neonates is in concordance with our previous study, showing a role for Fndc5 during neural differentiation. In this regard, both downregulation and upregulation of Fndc5 in neural precursor cells decelerate and accelerate neural differentiation, respectively (Hashemi et al., 2013; Forouzanfar et al., 2015). Another possibility is that the Fndc5 expression, not irisin, may be related to the developmental process rather than normal maintenance of energy homeostasis. Considering few studies on the role of Fndc5 in developmental processes, it is important to clarify the exact function of Fndc5 during these processes in the future.
Conclusion
The results of this study provide correlative evidence showing a discordance between RNA and protein levels of Fndc5 and the levels of RNA and protein expression between adult and neonatal brain tissue, which is likely to be regulated by microRNAs at the posttranscriptional level. Further studies, including experimental validation of these predicted microRNAs targeting Fndc5 as well as their upregulation or downregulation, are needed to verify this hypothesis.
Footnotes
Acknowledgments
This work was supported by the following grants: 394367, funded by Isfahan University of Medical Sciences, and 91000653, funded by Royan Institute for Biotechnology.
Disclosure Statement
No competing financial interests exist and all authors support the submission of this journal.