
Abstract
Peroxisomes are membrane-bound organelles that are best known for their roles in lipid metabolism. Mounting evidence indicates that they are also important nodes for antiviral signaling. While research over the past few decades has revealed effective viral strategies to block antiviral signalling pathways from the plasma membrane, mitochondria and/or the nucleus, until recently, very little was known about how viruses interfere with peroxisome-based antiviral signaling. In this essay, we review how viruses use a variety of strategies to interfere with peroxisome biogenesis, a phenomenon that has implications for evasion of the host immune system as well as pathogenesis.
Introduction
H
Among the host factors that have been linked to HAND is altered expression of miRNAs (Asahchop et al., 2016). These small noncoding RNAs affect the expression of many host as well as viral genes (Guo et al., 2010). As such, changes in cellular miRNA levels have been observed in a wide range of pathological conditions (Montano, 2011), including HAND (Eacker et al., 2009; Sonntag, 2010; Zhou et al., 2012; Pacifici et al., 2013). To this end, a study originally designed to identify HAND-associated miRNA profiles in brain tissue of HIV-infected patients revealed that HIV-1 infection upregulates a group of miRNAs that target mRNAs encoding proteins required for peroxisome formation (Xu et al., 2017).
As well as promoting loss of peroxisome assembly factors, including Pex2, 7, 11B, and 13, HIV-1 infection, was found to cause dramatic loss of peroxisomes in multiple cell types. Of note, inherited peroxisomal membrane transporter disorders, such as adrenoleukodystrophy resemble severe HAND in terms of clinical, radiological, and neuropathological features (Poll-The and Engelen, 2012). Because peroxisomes are neuroprotective and play critical roles in brain function (Smith et al., 1999), it is tempting to speculate that loss of peroxisomes in brain tissue of patients with HIV-1 infection contributes to HAND. However, given that peroxisomes have recently been recognized to play roles in early antiviral defense (Dixit et al., 2010) and depletion of peroxisomes has also been observed during flavivirus infection (You et al., 2015), it is also likely that targeted elimination of this organelle is a common strategy among RNA viruses at least.
Peroxisomes and antiviral defense
Peroxisomes are membranous organelles that are present in all eukaryotic cells. Genetic mutations that affect peroxisome function and/or biogenesis in humans are almost invariably incompatible with a normal lifespan due to serious neurological disorders and other developmental defects [reviewed in Smith and Aitchison (2013)]. Formation of peroxisomes requires at least 14 biogenesis factors or Pex proteins and can arise by de novo or endoplasmic reticulum-associated pathways (Fig. 1).
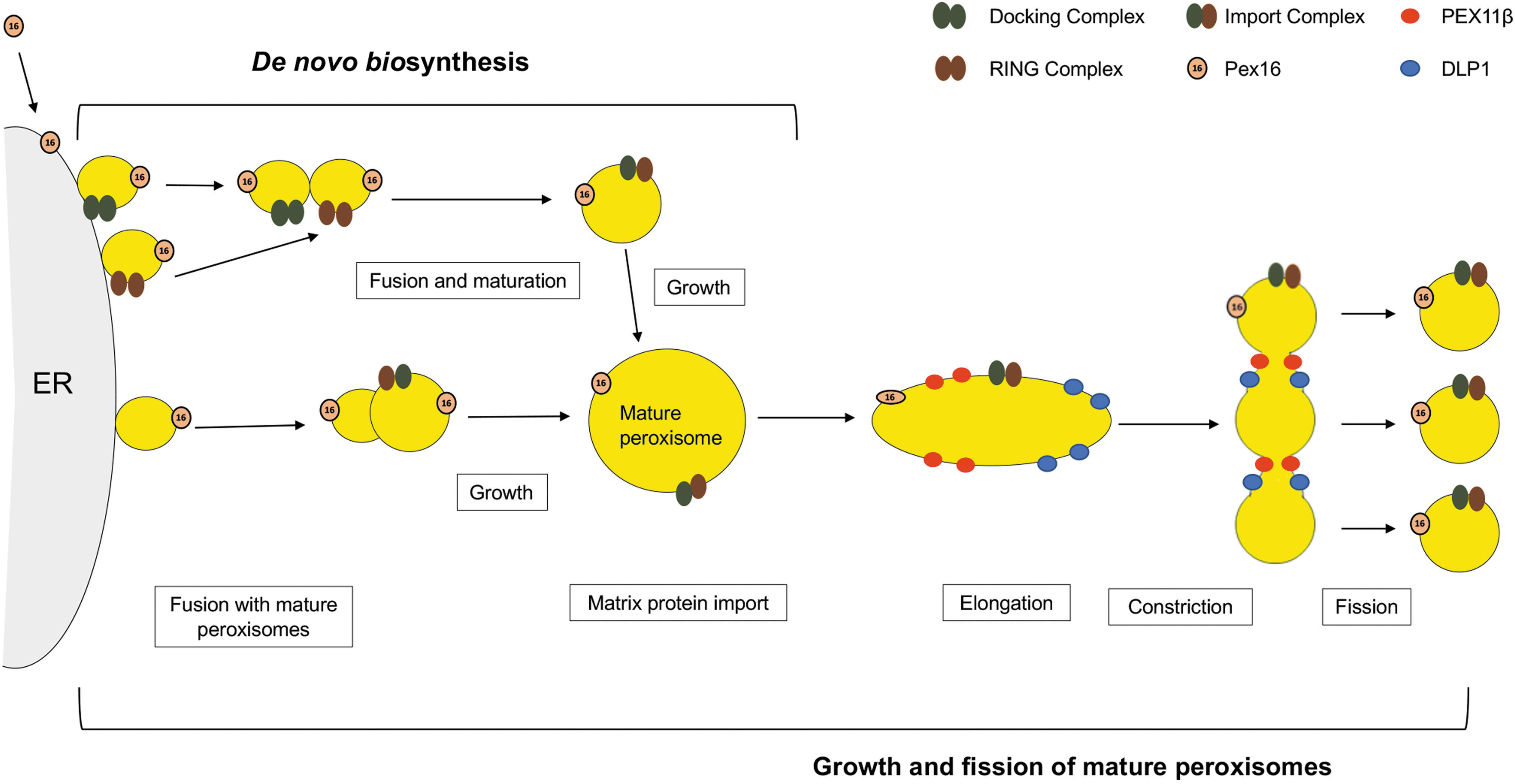
There are two primary pathways for peroxisome formation in mammalian cells: de novo biogenesis growth and fission of mature peroxisomes. In the de novo biogenesis pathway, the integral membrane protein Pex16 is required for budding of two forms of preperoxisomal vesicles from specialized regions of the ER. The preperoxisomal vesicles, which contain either docking complexes (Pex13/Pex14) or RING complexes (Pex2/Pex10/Pex12), fuse with each other in a Pex1/Pex6-dependent manner. Maturation and growth of these nascent peroxisomes occur by importing matrix proteins. Peroxisomes can also be formed by fusion of ER-derived preperoxisomal vesicles that lack docking or ring complexes, with preexisting mature peroxisomes. Growth and elongation of mature peroxisomes is mediated by Pex11 family and DRP-interacting proteins. Subsequent membrane constriction and fission are facilitated by DLP1 (Smith and Aitchison, 2013). DLP1, DRP, dynamin-like protein 1; dynamin-related proteins; ER, endoplasmic reticulum.
Within the peroxisomal matrix, there are more than 50 different enzymes that carry out critical metabolic functions, including production and degradation of hydrogen peroxide, oxidation of fatty acids, and synthesis of specialized lipids, all of which are important for development and function of the central nervous system (Wanders and Waterham, 2006). More recently, peroxisomes have emerged as signaling hubs for early antiviral defense pathways, including the IFN response (Dixit et al., 2010). Given that pervasive activation of type 1 IFN expression is common in patients with HAND (Rustagi and Gale, 2014), it is tempting to speculate that deregulation of the IFN system at the level of peroxisomes is a contributing factor to neurological disease in HIV patients.
It is widely recognized that mitochondria, which interact extensively with peroxisomes, plays critical roles in antiviral defense. One of the canonical antiviral signaling pathways that intersect with this organelle (Fig. 2) is activated following detection of viral RNA by RIG-I-like receptors (RLRs), such as RIG-I and MDA-5. RLRs are RNA helicases that recognize long double-strand RNA, a replication intermediate of RNA viruses or the structures found on the 5′ ends of viral RNA (Cui et al., 2008; Jensen and Thomsen, 2012). Binding to viral RNA induces conformational changes in RLRs, which allow them to interact with MAVS, an adaptor protein that resides on the surface of mitochondria (Kawai et al., 2005; Seth et al., 2005). Subsequently, MAVS signals downstream in parallel pathways to activate the antiviral transcription factors, IRF-3 and NF-kB, which activates transcription of IFN-β, a highly potent cytokine that in turn stimulates transcription of hundreds of other antiviral genes (Seth et al., 2005; Nakhaei et al., 2009).
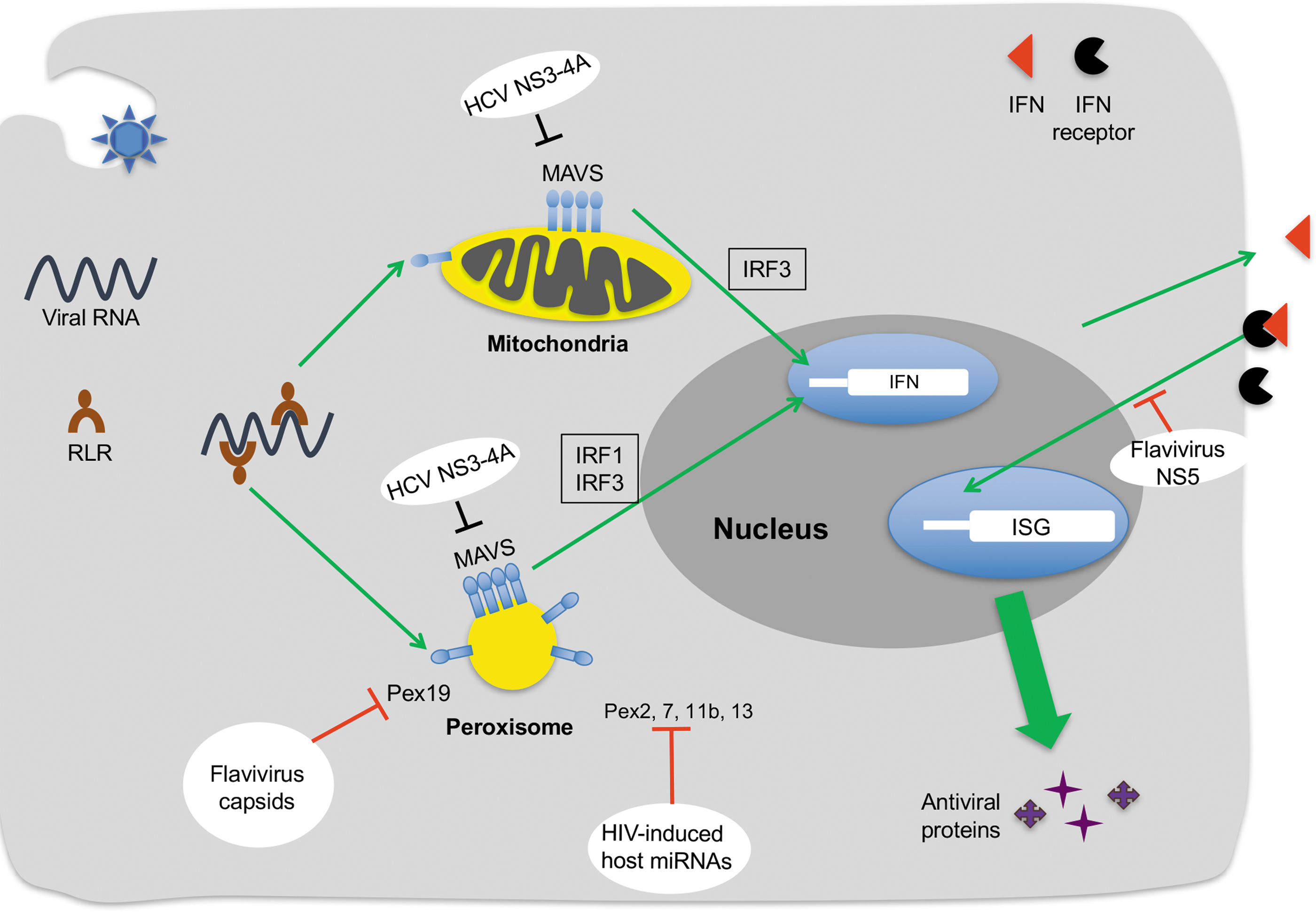
Viral interference with peroxisome-dependent antiviral signaling. Viral RNA in the cytosol is recognized by RLR, which then activates MAVS on peroxisomes and mitochondria. Peroxisomal MAVS may induce IRF1-IRF3-dependent expression of antiviral proteins encoded by ISGs through two different mechanisms. One of the pathways appears to be independent of IFN and results in very rapid induction of some ISGs and their encoded antiviral proteins (Dixit et al., 2010). The other is similar to signaling from mitochondrial MAVS which mediates the sustained expression of antiviral proteins through the type I IFN pathway. Flaviviruses, such as West Nile and Dengue viruses, inhibit peroxisome biogenesis in part by capsid protein-dependent sequestration and degradation of the peroxisomal biogenesis factor PEX19 (You et al., 2015). In contrast, the NS3-4A protease of the related pathogen, HCV impairs early antiviral signaling by cleaving MAVS on both mitochondria and peroxisomes (Horner et al., 2011; Bender et al., 2015; Ferreira et al., 2016). NS5 proteins of multiple flaviviruses block IFN signaling by causing degradation of STAT2 (Ashour, Laurent-Rolle et al., 2009; Grant, Ponia et al., 2016; Kumar, Hou et al., 2016). HIV-1 also impairs peroxisome biogenesis by inducing expression of host miRNAs that suppress expression of the peroxisome biogenesis factors PEX2, 7, 11b and 13 (Xu et al., 2017). Some DNA viruses (not shown) are known to interfere with the peroxisome-mediated antiviral response. The vMIA protein of human cytomegalovirus localizes to peroxisomes through interaction with PEX19 and interferes with antiviral signaling downstream from MAVS (Magalhaes et al., 2016). HSV1 dampens peroxisomal MAVS-dependent induction of IFN genes, a process that is attributed to the viral tegument protein VP16 (Zheng and Su, 2017). HCV, hepatitis C virus; HSV1, Herpes simplex virus-1; ISGs, interferon-stimulated genes; RLR, RIG-I-like receptor.
More recently, it was reported that RLRs can also signal through a pool of MAVS localized to peroxisomes following detection of viral RNA (Fig. 2). In this case, however, as well as IRF-3, another antiviral transcription factor, IRF-1 is activated followed by induction of type III IFN (Dixit et al., 2010). Further evidence of peroxisomal involvement in antiviral signaling stems from the observation that overexpression of Pex11β, which increases peroxisome numbers by stimulating elongation before division, induces expression of IFN-λ mRNA expression but not IFN-β during Sendai virus infection (Odendall et al., 2014). Conversely, knockdown of the peroxisome biogenesis factor Pex19 reduces expression of IFN-λ in response to the viral double-strand RNA mimics (You et al., 2015). Interestingly, in mouse embryonic fibroblasts, lack of peroxisomes does not affect induction of type I or III IFN expression in response to Sendai virus infection (Bender et al., 2015). Whether this indicates a fundamental difference between rodent and human antiviral signaling remains to be determined.
Viral interference with peroxisome function and biogenesis
The notion that peroxisomes are key players in antiviral defense is still a relatively new concept (Dixit et al., 2010). As such, we are only now beginning to understand how viruses interfere with antiviral signaling pathways that intersect with this organelle. One of the first indications of this phenomenon was the discovery that flaviviruses, such as West Nile and Dengue viruses, eliminate peroxisomes in part by capsid protein-dependent sequestration and degradation of the peroxisomal biogenesis factor Pex19 (You et al., 2015). Loss of peroxisomes may explain in part why induction of type III IFN is impaired during flavivirus infection. The related pathogen, hepatitis C virus, also impairs early antiviral signaling by targeting peroxisomes. However, in this case, the pool of MAVS on mitochondria is cleaved by the NS3-4A protease of hepatitis C virus (Horner et al., 2011; Bender et al., 2015; Ferreira et al., 2016). As of yet, it is not known whether hepatitis C virus infection actually reduces peroxisome numbers.
As mentioned above, HIV-1 impairs peroxisome biogenesis through a different mechanism than flaviviruses (Xu et al., 2017). Rather than directly interacting with proteins required for formation of peroxisomes, HIV-1 infection induces expression of miRNAs (miR-500a-5p, miR-34c-3p, miR-93-3p, and miR-381-3p) that suppress expression of the peroxisome biogenesis factors PEX2, PEX7, PEX11B, and PEX13. Reduction of peroxisomes was observed during acute as well as chronic HIV-1 infection. The latter may indicate that inhibition of peroxisome biogenesis is a key element of HIV-1 persistence and/or pathogenesis. Specifically, the dramatic reduction in peroxisomal proteins in brain tissue from HIV-infected patients may underlie some of the neurological disorders such as HAND or peripheral neuropathy associated with this virus.
Recent reports indicate that DNA viruses have also evolved strategies to evade peroxisome-mediated antiviral defense (Magalhaes et al., 2016; Zheng and Su, 2017). For example, the human cytomegalovirus protein vMIA, which is known to inhibit signaling downstream from mitochondrial MAVS, also localizes to peroxisomes through interaction with PEX19 (Magalhaes et al., 2016). Of note, interference of antiviral signaling by vMIA does not seem to require disruption of peroxisome morphology. Herpes simplex virus-1 infection has also been shown to dampen peroxisomal MAVS-dependent induction of IFN-stimulated genes, a process that is linked to the viral tegument protein VP16 (Zheng and Su, 2017). Finally, given the importance of peroxisomes in nervous system development and function, understanding how viruses manipulate peroxisomes will undoubtedly reveal pathogenic mechanisms that underlie virus-associated neurological disease in both children and adults.
Conclusions and Future Directions
While the metabolic roles of peroxisomes have been the subject of intense study for many years, it has only recently become apparent that this organelle functions in antiviral defense. The fact that multiple viruses use different mechanisms to reduce peroxisome numbers or interfere with their functions further underscores the importance of these organelles in the innate immune response. Importantly, the activity and abundance of peroxisomes can be manipulated through pharmacological means. Accordingly, these organelles may serve as cell-based targets for the development of broad-acting antiviral therapies. Indeed, recent studies in mice indicate that use of the peroxisome proliferator fenofibrate alone or as an adjuvant, showed significant protection from lethal challenge with influenza virus and Japanese encephalitis virus (Sehgal et al., 2012; Xu et al., 2015).
Footnotes
Acknowledgments
Research in the laboratories of TCH and CP is supported in part by a Canadian Institutes of Health Research (CIHR) Emerging Team grant (CP, TCH). CPW is supported by a graduate studentship from Alberta Innovates. CP and TCH are supported by Canada Research Chairs in Neurologic Infection & Immunity (CP) and RNA Viruses and Host Interactions (TCH).
Disclosure Statement
No competing financial interests exist.