
Abstract
Duchenne muscular dystrophy is the most common and severe form of progressive muscular dystrophy. Previous results showed an increased survival in double knockout mice (dko) when treated with adipose-derived CD146+ cells. In this study, we analyzed the effect of CD146+ cells compared to mesenchymal stem/stromal cells (MSCs) derived from the same human adipose sample when injected in the dko mouse model without immunosuppression. Both CD146+ cells and MSCs increased the survival of treated mice when compared to vehicle-injected mice, with a more prominent effect of CD146+ cells than MSCs. Both CD146+ cells and MSCs suppressed peripheral blood mononuclear cell proliferation, indicating immunomodulatory properties. Co-culture experiments showed that MSCs have a more inflammatory profile expression, and angiogenesis assay showed that CD146+ cells can improve blood vessel formation. CD146+ cells can extend survival of muscular dystrophy mice more efficiently than MSCs, possibly due to immunomodulatory and angiogenic properties. Further investigations focusing on exogenous CD146+ cell role in vivo will improve cell therapy understanding and effectiveness.
Introduction
P
The potential for muscular differentiation of adipose-derived mesenchymal stromal cells was already reported both in vitro (Vieira et al., 2008a) and in vivo (Rodriguez et al., 2005; Vieira et al., 2008b, 2012; Zhang et al., 2015). Mesenchymal stem/stromal cells (MSCs) are capable of secreting paracrine factors of clinical importance with diverse biological functions such as immunomodulation, antiscarring, chemoattraction, angiogenesis support, and antiapoptotic effect (reviewed in Meirelles et al., 2009; Caplan, 2017). According to a more recent review, it is suggested that MSCs should stand for Medicinal Signaling Cells (Caplan, 2017).
The classical protocol to obtain an MSC lineage consists basically in tissue digestion with collagenase, followed by the selection of adherent cells and discarding of culture media on the following days after cell plating. However, nowadays it is well known that the resultant population is highly heterogeneous. We observed that MSCs secrete different proteins even when they are obtained from the same tissue of different donors or from different tissues of the same donor (Assoni et al., 2017).
Attempts to isolate and study specific subpopulations from the stromal fraction of MSCs are ongoing. According to Zimmerlin et al. (2013), thee major subpopulation of cells can be found in human adipose stromal fractions: pericytes (CD45−, CD31−, and CD146+), endothelial progenitor cells (CD45−, CD31+, and CD34+), and supra adventitial adipose stromal cells (CD45−, CD31−, CD146−, and CD34+).
CD146+ cells, also known as pericytes, are perivascular cells currently under evaluation for the treatment of different pathologies (Dellavalle et al., 2007; Chen et al., 2013; Birbrair et al., 2014a; Valadares et al., 2014). CD146+ cells share common features with MSCs such as the expression of specific surface markers, secretion of paracrine factors and the ability to differentiate (Crisan et al., 2008; Corselli et al., 2010). Evidences indicate that MSCs are derived from these CD146+ cells, which are liberated from their basement membrane upon injury or inflammation (Caplan and Sorrell, 2015).
We and others have already reported therapeutic benefits in muscular dystrophy models when treated with MSCs from different sources, with a better effect of adipose-derived stromal cells when compared with umbilical cord-derived stromal cells (Vieira et al., 2008b, 2010, 2012; Rouger et al., 2011; Zucconi et al., 2011; Secco et al., 2013). Also, pericytes from adipose tissue, but not from other sources, significantly increased the life span of dko mice (Valadares et al., 2014). Although both MSCs and CD146+ cells derived from adipose tissue were found to be beneficial for muscular dystrophy preclinical treatment in mice models, we are not aware of studies comparing the clinical effect of both lines in vivo.
In this study, we compared the effect of CD146+ cells and MSCs derived from a single donor human adipose sample, when injected in the dko mouse model without immunosuppression.
Material and Methods
Ethics statement
This study was approved by the human research ethics committee (Comitê de ética em pesquisa—seres humanos—CEP), permit number 177/2013 and with informed consent of donor, and approved by the animal research ethics committee (Comissão de ética no uso de animais em experimentação—CEUA) of Institute of Bioscience, permit number 139/2013.
Tissue and cell preparation
One tissue specimen from human adipose tissue was obtained from total hysterectomy procedure. Tissue digestion and cell sorting by FACS were performed as reported in Valadares et al. (2014). After digestion, one portion of the stromal vascular fraction of cells was placed in culture, and by its adherent and in vitro differentiation capacities was classified as MSCs. The other portion was submitted to cell sorting, before culture, to enrich pericyte population according to the markers CD34−, CD45−, CD56−, and CD146+.
Animals
For the main experiment, 8-week-old dko mice were divided in three groups (n = 20 per group on average) that received weekly transplantation of CD146+ cells, MSCs, or Hanks' Balanced Salt Solution (HBSS, vehicle). Each injection contained 106 viable cells in 100 μL HBSS and were delivered intraperitoneally for a total of 8 weeks, without any immunosuppression.
Physical performance tests
Treadmill
Initial speed was 3 m/min and every minute, the speed was accelerated 3 m/min. The complete test lasted 10 min with a final speed of 30 m/min.
Kyphosis ratio
As described by Lima et al. (2010), a full body X-ray of each mouse was digitized and the measurements were taken using RadiAnt DICOM viewer 1.8. Spine kyphosis ratio (straight line distance/cervical-thoracic segment length) was used to score the animals according to the severity of the skeletal manifestation (the smaller the ratio, the more severe the manifestation).
In vitro myogenic differentiation
For myogenic differentiation, when cells reached a confluence of 80–90%, the proliferation media were changed to induction media, which consisted of Dulbecco's modified Eagle's medium (DMEM)-F12 with 2% horse serum for 15 days. Cells were then analyzed for expression of myogenic proteins by immunofluorescence. Briefly, cells were fixed in cold methanol for 5 min and then were blocked with phosphate-buffered saline (PBS) containing 10% SFB and 5% bovine serum albumin (BSA) for 1 h and then incubated for 1 h at 37°C with mouse anti- myosin heavy chain 2× (1:500, DHSB-MF20) and rabbit anti-dystrophin (1:200, 15277; Abcam). After three washes with PBS for 5 min, cells were incubated with secondary antibody against mouse (Alexa 488, 1:400; Invitrogen) and rabbit (Alexa 594, 1:400; Invitrogen). After incubation, cells were washed thrice with PBS and incubated with Hoechst 33342 (Sigma-Aldrich) for 5 min to visualize the nuclei. Fluorescence signal was examined in Axiovert 200 (
In vivo myogenic differentiation
The protocol was as described in Crisan et al. (2008). Black/SCID mice were anesthetized with xylazine/ketamine and an injury using 15 μg of cardiotoxin was made in the gastrocnemius of each mouse. Three hours later, animals were reanesthetized and received either 2 × 104 cells locally in the injured gastrocnemius or 105 cells intraperitoneally. After 21 days, animals were sacrificed and both gastrocnemius of each mouse were cryopreserved. Mice that did not receive any cells were used as negative controls. For the positive control, cells were transplanted into mice that were sacrificed right after local injection.
Immunofluorescence was performed to identify human cell with the briefly described protocol: fixation for 20 min in PFA 4%, wash thrice with PBS, permeabilization solution for 15 min (containing BSA and Triton X-100), block for 1 h (solution containing BSA, fetal bovine serum [FBS], and Triton X-100), incubated overnight with anti-human lamin A/C antibody (VP-L550 1:400; Vector), washed thrice with PBS, and incubated for 1 h with secondary antibody anti-mouse Cy3 (Cat. No. 111-165-003; Jackson ImmunoResearch).
In vitro angiogenic assay
Matrigel (356234; Corning) was defrosted in 4°C for at least 1 h. Fifty microliters of matrigel were plated per well in a 96-well plate. After Matrigel polymerization at 37°C for 30 min, 2 × 104 cells (human umbilical vein endothelial cell [HUVEC], CD146+ cells, or MSCs) were add to each well. When co-culture was performed, 2 × 104 of HUVEC were plated with 7.5 × 103 of CD146+ cells or MSCs. Cells were plated in duplicates for 24 h, time when endothelial tubular networks are undone. HUVEC were commercially obtained (Lonza).
In vivo angiogenic assay
The used protocol was similar to that described elsewhere (Birbrair et al., 2014b). Briefly, Matrigel-reduced growth factor (356230; Corning) was defrost in 4°C for at least 1 h. The cell suspension was mixed with 500 μL of matrigel at ratio 1:1, generating matrigel plugs, which were injected subcutaneously into dorsal region of nude mice. Suspended cells consisted of 4 × 106 HUVEC with either 5 × 105 MSCs or CD146+ cells. Therefore, angiogenic assay in vivo comprised three groups (two animals per group), where each group received a matrigel plug with the following cells: HUVEC alone; HUVEC and MSCs; and HUVEC and CD146+ cells. Matrigel plugs were recovered 3 weeks later.
Lymphocyte proliferation assay
Peripheral blood mononuclear cells (PBMCs) were collected from mouse peripheral blood. MSCs and CD146+ cells were cultivated in different cell concentrations per well (103, 5 × 103, 104, 5 × 104, and 105 cells) and then treated with mitomycin C for 3 h. Cells were washed twice with PBS and 104 PBMCs, previously activated with antibody anti-CD3 (Cat. No. 100340; Biolegend), were added to each well. The assay was performed in triplicates and activated lymphocytes cultured alone served as controls. The cells were maintained in RPMI medium at 37°C and 5% CO2 for 5 days. Thereafter, the lymphocytes were resuspended and CFDA staining was evaluated in a Guava EasyCyte flow cytometer running Guava Express Pro software (Guava Technologies).
Co-culture assay and cytokine analysis
To investigate MSCs' and CD146+ cells' cytokine profiles in response to dko environment, an in vitro co-culture of these cells was established. Murine cells were obtained from a 2-month-old female mouse that underwent euthanasia, when gastrocnemius was collected. Briefly, for tissue explants, gastrocnemius is washed twice with PBS, minced in small parts that are binded to culture flask with FBS overnight before contact to growth media (DMEM high glucose with 20% FBS). Growth media were changed twice a week and when cells reached a confluence of 80–90% in the culture flask, cells were detached using Trypsin-EDTA solution (0.05%) to perform a cell passage. For co-culture assay, a 24-well plate was used plating 4 × 104 murine muscle cells and 2 × 104 human CD146+ cells or MSCs. Murine and human cells were separated by transwells, with no contact among cells. Murine cells, MSCs, and CD146+ cells were also cultured alone as control of cytokine expression.
After 96 h, conditioned media were collected from co-cultures and analyzed by Luminex technology comprising 27 cytokines (Bio-Plex Pro Human Cytokine Standard 27-plex, Group I; Cat. No. M500KCAF0Y; BioRad). Also, media alone (without cell contact) were used as control.
Statistical analysis
The following tests were applied.
Weight: one-way analysis of variance test.
Survival curve by log-rank trend test: to verify whether there was a trend in the survival curves (indicating a difference between them), the dko mice were divided into three groups: CD146+ cell-treated group n = 19; MSC-treated group n = 22; and vehicle-treated group n = 20.
Survival curve by log-rank (Mantel-Cox) test and Gehan-Breslow-Wilcoxon test: to evaluate whether the survival curves after administration of the specific cells were significantly different (p < 0.05).
Spinal curvature ratio and Treadmill: Mann–Whitney test.
All data used in the survival curves were measured in days after the onset of treatment. Outliers within each group were excluded using Grubbs' test (ESD method) considering the value for alpha as 0.05.
Results
Cell line characterization
Both cell lines were characterized through their in vitro differentiation potential. MSCs were able to differentiate into osteoblasts, chondroblasts, and adipocytes, while CD146+ cells were able to differentiate only in osteoblasts and chondroblasts (data not shown). Both lineages were also analyzed by flow cytometry to evaluate their immunophenotypic profile in passage 8, the same used for cell transplantation in vivo. CD146+ cells and MSCs presented low expression of hematopoietic markers (CD34 and CD45) and low expression of muscle progenitor marker CD56 (Supplementary Table S1; Supplementary Data are available online at
Physical performance tests and survival analysis
Both CD146+ cells and MSCs tend to improve the survival of treated mice when compared to vehicle-injected mice, with a better effect of CD146+ cells than MSCs (Fig. 1). However, the trend was statistically significant only during the treatment period, suggesting that these cells have a short-term effect (Fig. 1).
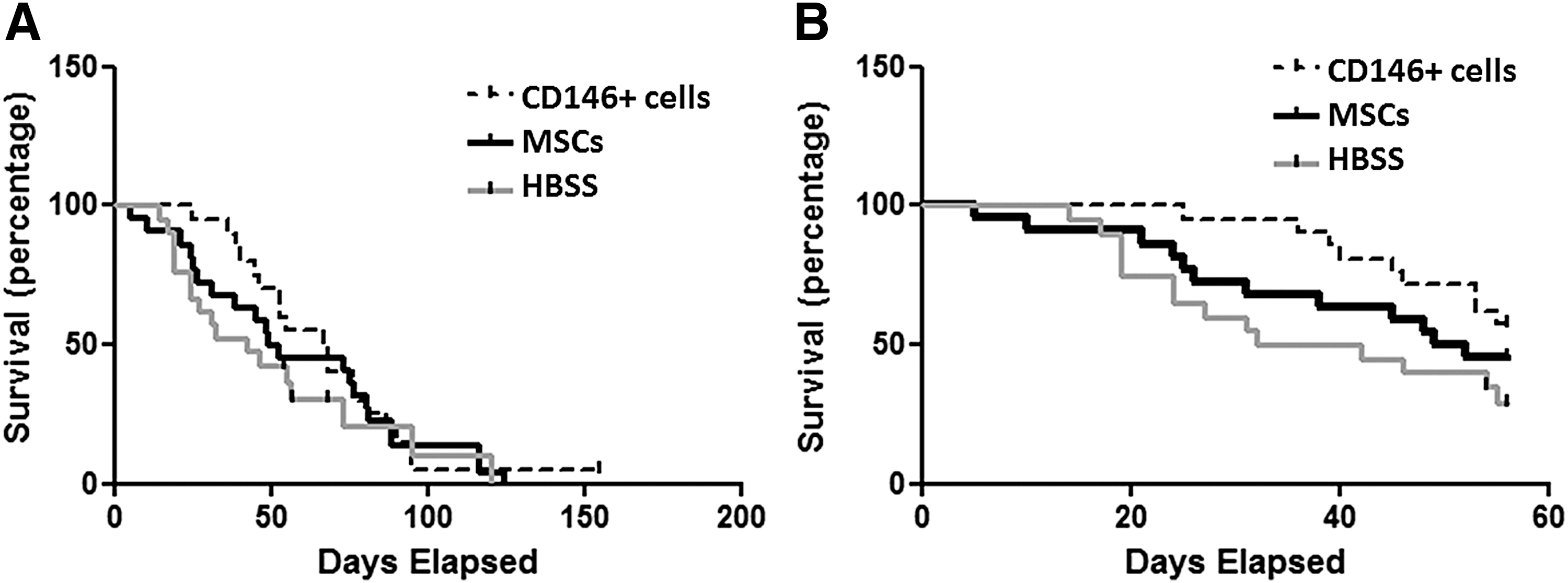
Day 0 represents first day of treatment. Survival curves of all animal life spans starting
Weight, kyphosis ratio, and treadmill performance did not differ among experimental groups (Supplementary Figs. S1–S3). Therefore, we concentrated our conclusion on the survival data.
Myogenic differentiation
As expected, commercial myoblasts used as positive control expressed myosin heavy chain and dystrophin and were able to fuse in myotubes, after 15 days in culture with myogenic inducing media (Supplementary Fig. S4). However, these features were absent in both MSCs and CD146+ cells.
Myogenic potential was also evaluated in vivo in immunosuppressed mice injured with cardiotoxin in their gastrocnemius. Immunosuppressed mice received cells, locally or intraperitoneally, after an injury was induced on their gastrocnemius with cardiotoxin. Three weeks after CD146+ cell or MSC transplantation, muscles were collected for histological evaluation, but no human cells were identified in those tissues (Supplementary Fig. S5).
Angiogenic assays
CD146+ cells alone were able to form endothelial tubular networks in vitro after 2 h of plating, while this ability was absent in MSCs cultured on Matrigel (Fig. 2).
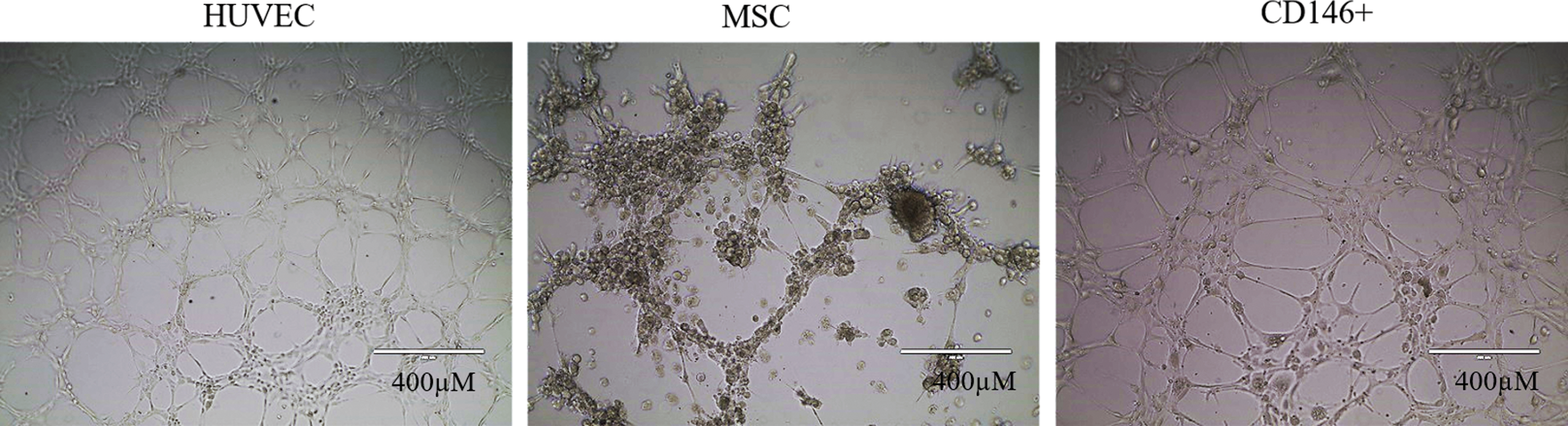
Angiogenic assay. In vitro evaluation with positive control (HUVEC), CD146+ cells, or MSCs. CD146+ cells alone showed angiogenic potential. Tubular structure after 2 h of plating. Scale 1:400 μm (10 × ). HUVEC, human umbilical vein endothelial cell. Color images available online at
Two-month-old nude mice were used to evaluate the angiogenic potential of CD146+ cells and MSCs in vivo. We observed that CD146+ cells, when mixed to HUVEC cells, were able to assist blood vessel formation into Matrigel plugs, while this was not observed for HUVEC or HUVEC plus MSCs (Fig. 3).
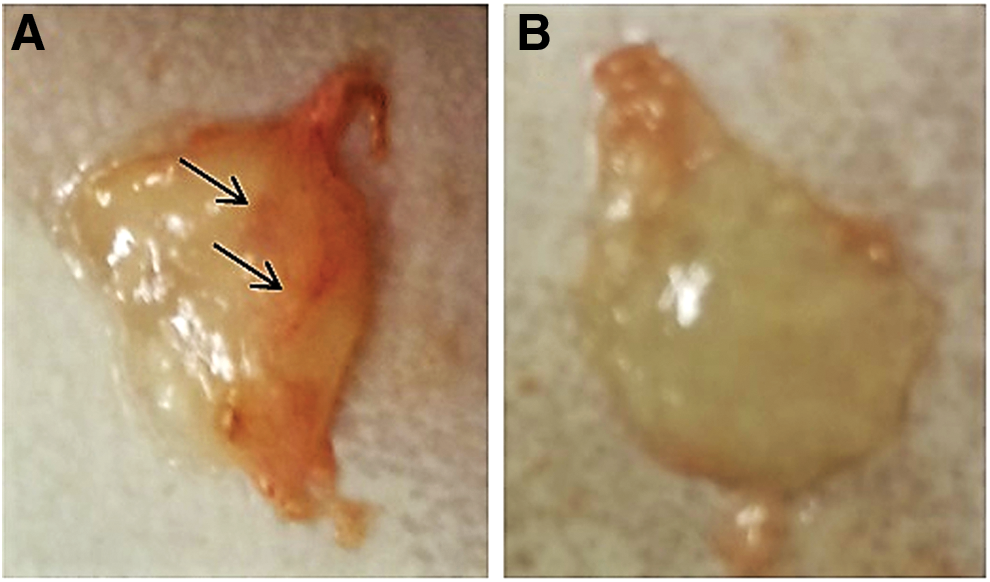
Angiogenic assay in vivo. Matrigel plugs were constituted of HUVEC and CD146+ cells
Immunomodulation ability
To evaluate the extension of the paracrine effect of CD146+ cells and MSCs, the immunomodulatory potential was evaluated through lymphocyte proliferation assay and cytokine quantification.
We observed that both CD146+ cells and MSCs were equally able to suppress lymphocyte proliferation in a dose-dependent manner (Supplementary Fig. S6).
Next, to further evaluate which cytokines are produced by each cell line, we performed co-culture assay, using dko muscle cells to represent the environment of dystrophic muscle and CD146+ cells or MSCs, which were separated by transwell inserts. Murine muscle cells, MSCs, and CD146+ cells were also cultured alone as controls of cytokine expression. After 96 h, conditioned media were collected from co-cultures and analyzed by Luminex technology to detect human cytokines. Our results showed that some cytokines and chemokines, such as IL12p70, IL6, IL8, MCP1, and RANTES, were produced in higher rates by MSCs than CD146+ cells (Supplementary Fig. S7). Despite some of them can act as proinflammatory or anti-inflammatory cytokine, IL12p70, MCP1, and RANTES are proinflammatory.
Discussion
DMD is a genetic disease caused by mutations in the X-linked dystrophin gene, resulting in protein deficiency causing muscle degeneration and early death in affected boys. Involvement of the immune system in dystrophinopathies has been suggested (Spencer and Tidball, 2001). It is well known, for example, that DMD patients present accumulation of immune cells in their muscles (Arahata and Engel, 1984). The fact that immunosuppressive drugs, such as glucocorticoids, can alleviate inflammation and muscle weakness in DMD patients and muscular dystrophy in animal models, despite their numerous side effects, supports these observations (Davies and Grounds, 2006; Bushby et al., 2010; Serra et al., 2012; Arpan et al., 2014). Currently, there is no efficient treatment for DMD patients; the potential benefits of cell therapy are being evaluated.
In this study, we compared the therapeutic potential of MSCs and pericytes obtained from the adipose tissue of the same human donor. The clinical impact of mouse cells transplanted into mouse recipients should be also evaluated in future studies. However, the use of human cells in preclinical studies with mice allows to observe the fate of injected cells in vivo, aiding the design of future clinical trials in humans.
In a previous study from our group, pericytes from the adipose tissue, but not from other sources, were able to increase the lifespan of muscular dystrophy mice (Valadares, 2014). In this study, we compared the therapeutic effect of MSCs and a specific subpopulation of these cells, CD146+ cells. In this work, cell characterization revealed that CD146+ cells maintained CD146 expression after eight passages in culture. As for MSCs, we noticed that approximately 40% of cells were positive for CD146+, highlighting the heterogeneity of this cellular vascular fraction.
Logrank-trend test revealed a significant trend of increased survival in CD146+ cell and MSC groups in comparison to vehicle-treated dko mice, this effect being more prominent for mice treated with CD146+ cells alone. This result indicates that, despite being derived from the same healthy donor, biological differences existing in these two lineages may have a different therapeutic potential. To deeply investigate those inherent differences, we compared their differentiation and angiogenic capacities, and their immunomodulatory potential.
Our results showed that neither CD146+ cells nor MSCs could differentiate into muscle cells in vitro or in vivo. Regarding their immunomodulatory potential, we observed that MSCs and CD146+ cells equally suppressed the proliferation of mouse lymphocytes in a dose-dependent manner, starting from 5:1 (human cells/mouse cells) ratio.
Cytokine quantification in the co-culture medium of MSCs or CD146+ cells with dko muscle cells demonstrated that MSCs produced numerous chemokines and cytokines in higher rates than CD146+ cells, similar to the results found in our previous study (Coatti et al., 2017). IL12p70, RANTES, and MCP1 are either cytokines or chemokines with a proinflammatory profile. IL6 and IL8 can act as proinflammatory or anti-inflammatory cytokines (Kyurkchiev et al., 2014; Admyre et al., 2015). These results suggest that MSCs have a proinflammatory profile when co-cultured with dystrophic murine muscle cells, which could accentuate the infiltration of inflammatory cells and fibrosis in the muscle.
It has been observed by us and others that MSCs secrete different proteins even when they are obtained from the same tissue of different donors or from different tissues of the same donor (Assoni et al., 2017). This could be related to the existence of different subpopulations within each cell lineage, such as cells with proinflammatory or anti-inflammatory profile. It is possible that specific stimulus before transplantation could determine which profile these cells will present, improving results (Waterman et al., 2010). To avoid the genetic variation observed between different donors, in this study, we used MSCs and CD146+ cells obtained from a single donor and thus with the same genetic background. Future studies should be performed with other CD146+ lineages obtained from different donors to attest this effect. In addition, a deeper investigation of what molecules are being secreted by these cells is important, allowing researchers to personalize cell therapy for each condition.
Besides the myogenic potential, we evaluated their ability to assist the formation of blood vessels, since a higher vascularization could be beneficial to muscular dystrophy treatment, contributing to injured tissue recovery and a better delivery of precursors associated to vessels (Corselli et al., 2013; Murray et al., 2014). The angiogenic assay revealed that, CD146+ cells, but not MSCs, could promote/assist blood vessel formation in vitro and in vivo, which could at least partially explain the different clinical outcome in treated mice.
Our group showed previously that the beneficial effect of injected adipose pericytes in dko mice (Valadares et al., 2014) was probably due to a paracrine effect through soluble factors, since we were not able to trace injected cells. Similar to these findings, other reports have suggested that such paracrine effect is the main mechanism of tissue repair through transplanted progenitor cells (Lavasani et al., 2012; Coatti, 2017). To provide a more detailed description of the fate of transplanted cells, pericytes obtained from NG2-DsRed mice, which naturally express a red fluorescent protein, could be traced in dko mice after transplantation.
Similar to these results, Valadares et al. (2014) observed that human adipose-derived pericytes were able to increase the life span of dko mice. For that purpose, they obtained the cells from a different donor than the one used in this study (Valadares et al., 2014). Although CD146+ cells are more homogenous than MSCs, they are not completely clonal. Birbrair et al. (2014a, 2014b) described two different mice pericyte subpopulations with different biological potentials. Assoni et al. (2017) also observed that MSCs obtained from different donors possess a different secretory profile. The evaluation of other lineages of CD146+ cells obtained from different individuals will be helpful to better characterize such differences. Different protocols may improve the clinical outcome, for example, extending cell administration during longer periods.
Conclusion
In short, in this study, we report for the first time that human adipose-derived CD146+ cells can extend survival of a muscular dystrophy mouse model more efficiently than MSCs. Our experiments suggest that cells were acting through the immunomodulation of the existing inflammation in the host muscle and possibly enhancing blood vessel formation, which would be beneficial to muscle cells delaying their degeneration.
Footnotes
Acknowledgments
We would like to thank Eder Zucconi, Tatiana Jazedje, Natale Cavaçana, Naila Lourenço, Michel Naslavsky, Melinda Becari, Miriam Frangini, and Aline Lopes for their helpful suggestions throughout the project; Professor Stefano Hagen, for his cooperation on X-ray animal examinations; Constância Gotto, Vanessa Sato, and Wagner Falciano for administrative support; and Neide Mascarenhas and co-workers from IPEN for the collaboration regarding animal care. This work was supported by CEPID/FAPESP, INCT, and CNPq and the Processo FAPESP 2013/08028-1 grant.
Disclosure Statement
All authors declare no potential conflict of interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.