
Abstract
Nd1-S is the nuclear-localizing short variant form of Nd1 (Ivns1abp) encoding a Kelch family transcription factor. While the function of Nd1 has been investigated in the context of metastasis and doxorubicin-induced cardiotoxicity, little is known about its role in hematopoiesis. In this study, we investigated the function of Nd1-S in hematopoiesis by transplanting the Nd1-S-overexpressing murine hematopoietic stem and progenitor cells (HSPCs) into recipient mice (Nd1-S mice). Enforced expression of Nd1-S led to erythroid and megakaryocyte dysplasia, demonstrated by dramatically decreased red blood cells and platelets, and megakaryocytes in the peripheral blood and bone marrow of the Nd1-S mice. Moreover, phenotypic megakaryocyte–erythroid progenitors (MEPs) accumulated in these Nd1-S mice with aberrant morphology and defective colony-forming capability. Furthermore, these phenotypic MEPs showed impaired pathways regulating erythroid differentiation and megakaryocyte development. Therefore, our study provides de novo evidence that overexpression of Nd1-S in HSPCs leads to erythroid and megakaryocyte dysplasia in vivo by targeting MEPs.
Introduction
N
To investigate the role of Nd1-S in hematopoiesis, we adopted a classic retroviral approach to overexpress Nd1-S in hematopoietic stem and progenitor cells (HSPCs), and studied its effects in hematopoietic repopulation in vivo by transplantation setting. We demonstrated that enforced Nd1-S expression led to erythroid and megakaryocyte dysplasia in peripheral blood and bone marrow of Nd1-S transplanted mice. Phenotypic megakaryocyte–erythroid progenitors (MEPs) accumulated in these mice with disrupted morphology and impaired colony-forming capability. RNA-Seq and GSEA analysis revealed that these MEPs overexpressing Nd1-S showed deregulated pathways for erythroid differentiation, megakaryocyte, and platelet development. Therefore, our study provides de novo evidence that overexpression of Nd1-S in HSPCs leads to erythroid and megakaryocyte dysplasia in vivo by impairing the differentiation of MEPs.
Materials and Methods
Mice
Eight- to ten-week-old female mice used in this study were housed in the SPF-grade animal facility of Guangzhou Institutes of Biomedicine and Health (GIBH). CD45.1 strain was originally bought from Jackson Laboratories. Donor fetal liver lineage negative cells were isolated from C57BL/6-CD45.2 embryos (E14.5) (Kim et al., 2006). All animal experiments were carried out in accordance with the Guide for the Care and Use of Laboratory Animals instructed by the Institutional Animal Care and Use Committee of Guangzhou Institutes of Biomedicine and Health, (IACUC-GIBH) and the committee approved all experimental protocols (2012013).
Retroviral transduction
Nd1-S cDNA was inserted into pMYs-IRES-EGFP to generate the pMYs-Nd1-S-IRES-EGFP vector (RTV-021; Cell Biolabs, Inc.). pMYs-IRES-EGFP or pMYs-Nd1-S-IRES-EGFP were transfected into Plat-E cells to produce high-titer, replication-incompetent retroviruses. The retrovirus supernatant was collected 48–72 h later and used freshly after collection. Lineage negative fetal liver (Lin− FL) cells were enriched based on lineage depletion method by AutoMACS (Miltenyi Biotec). Enriched Lin− FL cells were suspended in viral supernatants (1 × 105 cells/mL) with 8 μg/mL Polybrene (Sigma-Aldrich) and centrifuged at 800 g for 120 min, followed by 24-h culture in StemSpan SFEM (09600; Stem Cell Technologies) in the presence of 10 μg/mL heparin, 50 ng/mL SCF (300-07; PeproTech), and 20 ng/mL TPO (300-18; PeproTech). Cells underwent two rounds of spin infection before transplantation as previous research did (Zhang et al., 2018).
Quantitative real-time PCR
Total RNA was extracted with TRIzol (TR 118; Molecular Research Center, Inc., Cincinnati, OH) according to the manufacturer's protocol. Up to 1 μg total RNA was reverse transcribed to cDNA using the ReverTra Ace qPCR RT Master Mix with the gDNA Remover Kit (FSQ-301; TOYOBO, Osaka, Japan). Quantitative real-time PCR was performed using SsoAdvanced SYBR Green Supermix (1725274; BIO-RAD, Hercules, CA) on CFX96 (BIO-RAD). Gapdh was used for qPCR normalization of the samples. The primers were used as follows: Nd1-total (forward: GCATACACGGCTCAGTTGAAAGCTG, reverse: GAAATTCCTCCTCTTCTGAAATTTG); Nd1-L (forward: CAAATTTCAGAAGAGGAGGAATTTC, reverse: GCCACTTATCTGCTTATGGCCATTC); and Gapdh (forward: CCTGGAGAAACCTGCCAAGTATG, reverse: CCTGGAGAAACCTGCCAAGTATG); All reactions were performed in triplicate.
Antibodies
Mac1 (M1/70), CD90.2 (53-2.1), B220 (6B2), CD2 (RM2-5), CD3 (145-2C11), CD4 (RM4-5), CD8 (53-6.7), Gr1 (RB6-8C5), TER119 (TER-119), CD127 (B12-1), c-kit (2B8), Sca-1 (E13-161.7), CD34 (RAM34), IgM (II/41), and CD16/CD32 (93) antibodies conjugated with biotin or specific dyes as required were purchased from eBioscience. CD150 (TC15-12F12.2) antibodies was purchased from BioLegend.
Cell cycle analysis
Cell cycle analysis was performed as previously described (Wang et al., 2013). Briefly, for cell cycle assay of MEPs, antibodies recognizing lineage markers (CD2, CD3, CD4, CD8, Mac1, Gr1, B220, TER119, and IgM) and CD127 were added first. After wash, CD16/32-PE, APC-eFluor 780-conjugated streptavidin, PE-CF594-c-kit, PE-Cy7-Sca-1, and PerCP-Cy5.5-CD34 antibodies were added. Stained cells were then fixed with 4% PFA and permeabilized with 0.1% saponin in PBS. Cells were subsequently stained by PE-KI67 (eBioscience) and DAPI (Invitrogen) for 45 min before examination. The stained cells were analyzed on Fortessa SORP (BD Bioscience), and data were analyzed by FlowJo software (FlowJo).
Transplantation
Adult C57BL/6 recipient mice (CD45.1+, 8–10 weeks old) were irradiated with two doses of 550 rads (RS 2000; Rad Source), 4 h apart. Two hundred sorted GFP+ LSK cells (CD45.2+) with 0.25 million Sca-1 depleted helpers were injected into the retro-orbital venous sinus of irradiated CD45.1-recipients. The transplanted mice were maintained on trimethoprim–sulfamethoxazole-treated water for 2 weeks. After transplantation, peripheral blood was obtained from the retro-orbital venous sinus monthly for flow cytometric analysis. All experiments that require termination of recipients' survival were carried out 16 weeks posttransplantation, including HE section of bone marrow, analysis of myeloid progenitors and RNA-Seq of phenotypic Nd1-S-MEPs, and control.
RNA-seq
One thousand cells of hematopoietic stem cells (Lin−CD48−c-kit+Sca-1+CD150+) and multipotent progenitors (Lin−CD48−c-kit+Sca-1+CD150−) compartment were sorted, respectively, into collection buffer and pellet down. Subsequent to lysis, reverse transcription and amplification were optimized based on single-cell RNA sequencing protocol (Tang et al., 2010). Sequencing libraries were then generated with the Nextera XT Sample Preparation Kit (Illumina). More than 50,000 cells of myeloid progenitors (Lin−CD48−Sca-1−c-kit+), T (CD90.2+), B (B220+), Myeloid (Mac1+), and MEP (Lin−CD127−Sca-1−c-kit+CD16/32−CD34−) were sorted into RLT lysis buffer and sequencing libraries were generated with the TruSeq RNA Sample Preparation Kits (Illumina) as previously described (Yang et al., 2015). Samples were sequenced on the Illumina Nextseq500 sequencer. Sequence reads were processed and analyzed with the help of the GIBH bioinformatics core.
Statistical analysis
Data are presented as mean ± SDs. Student's t-tests were performed using statistic software GraphPad Prism 6 and p-values were indicated in figure legends.
Results
Enforced expression of Nd1-S in HSPCs induces pancytopenia in the peripheral blood of recipient mice
To investigate the expression pattern of Nd1 in hematopoietic components, we sorted hematopoietic stem cells (HSCs), multipotent progenitors (MPPs), myeloid progenitors (MPs), and other mature blood lineages, and performed RNA sequencing (RNA-Seq). RNA-Seq data indicated that Nd1 was highly expressed in myeloid progenitors, which implies Nd1 as a regulator of hematopoiesis, especially in myeloid progenitor context (Supplementary Fig. S1a; Supplementary Data are available online at
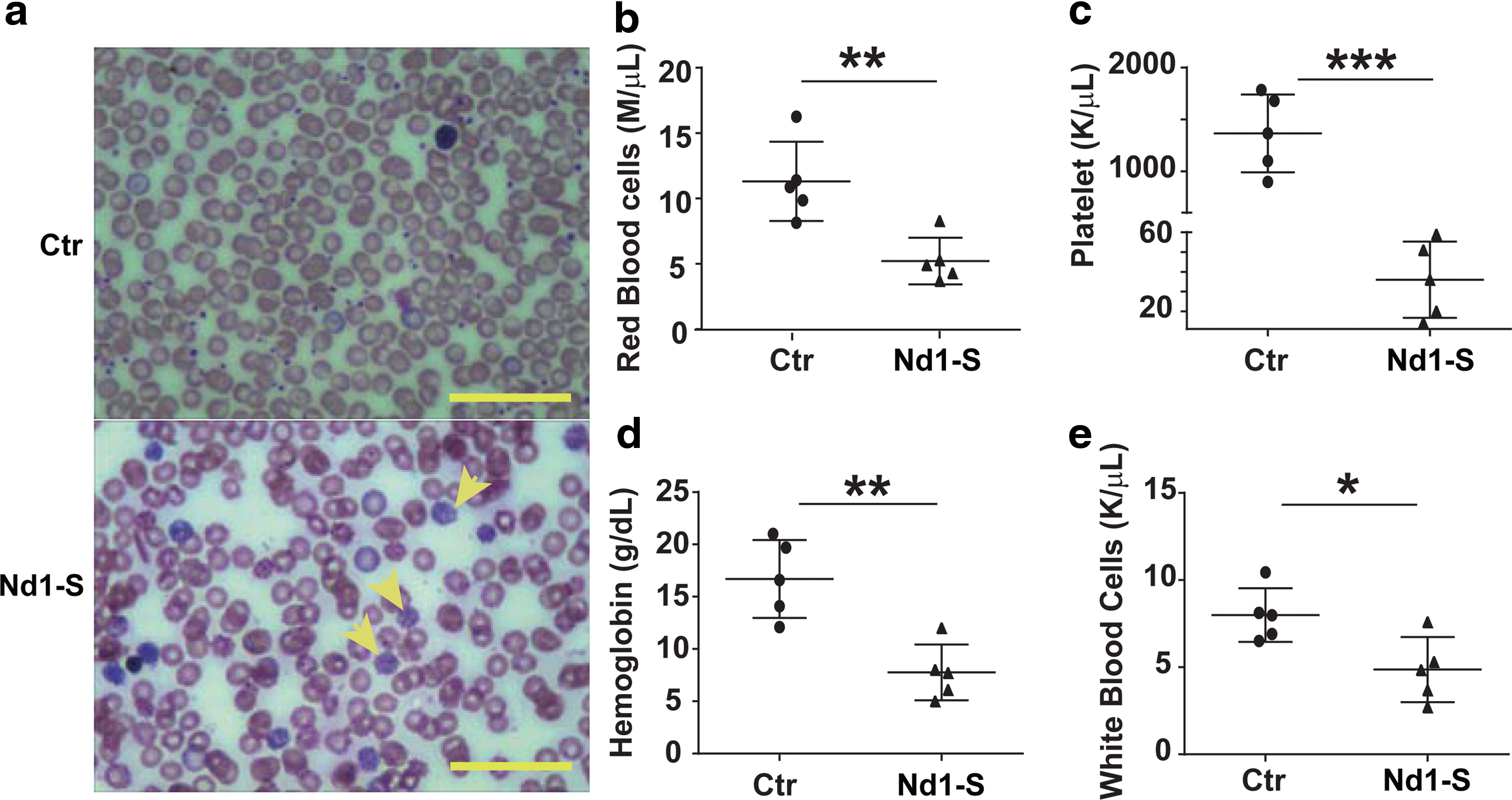
Ectopic expression of Nd1-S induces pancytopenia in peripheral blood.
Overexpression of Nd1-S induces severe cytopenia in the bone marrow of recipient mice
To determine whether the depletion of erythrocytes and megakaryocytes is due to erythroid and megakaryocyte dysplasia in bone marrow, we sacrificed Nd1-S mice at 16 weeks posttransplantation, and performed Hematoxylin and Eosin staining of their bone marrow cells. We indeed observed reduced number of red blood cells and diminished megakaryocytes in the bone marrow of these mice. In addition, the bone marrow from the Nd1-S HSPC recipients was pale and lacked lobulated-nucleus megakaryocytes (Fig. 2a), which was consistent with the phenotype of pancytopenia in peripheral blood. In addition, H&E staining of spleen sections from Nd1-S HSPC recipients indicated splenic parenchyma lacking erythrocytes (Fig. 2b). Altogether, these data demonstrated that overexpression of Nd1-S in HSPCs impaired erythropoiesis and megakaryopoiesis in vivo.

Overexpression of Nd1-S in HSPCs leads to erythroid and megakaryocyte dysplasia.
Nd1-S overexpression in HSPCs reduces BFU-E colony formation in vitro
To investigate whether Nd1-S disrupted differentiation potential of hematopoietic progenitor cells, GFP+ bone marrow cells from recipients were sorted and plated for CFU assay. Notably, overexpression of Nd1-S completely abolished the formation of BFU-E and CFU-GEMM, whereas other myeloid lineages remained unaffected (Fig. 3a, b). Giemsa staining of BFU-E and GEMM colonies confirmed the colonies' identity (Fig. 3c). Therefore, Nd1-S overexpression inhibited the differentiation of erythroid progenitor cells in vitro.
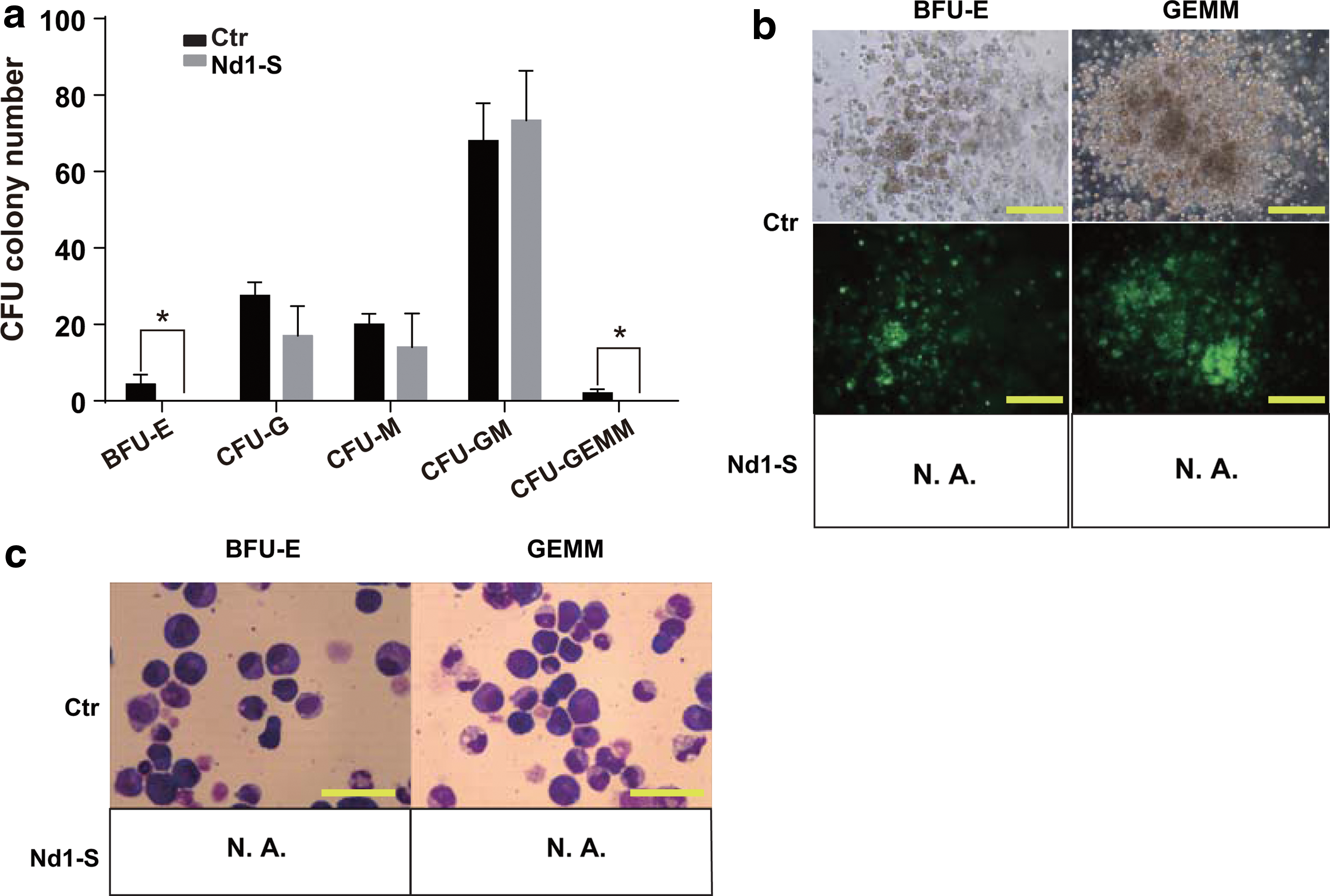
Nd1-S overexpression in HSPCs show rare BFU-E colonies in vitro. Recipients were sacrificed 16 weeks posttransplantation. The 2 × 104 GFP+ bone marrow cells from recipients were sorted and plated for each CFU plate.
Nd1-S overexpression leads to accumulation of phenotypic MEPs in the bone marrow of Nd1-S mice
It is possible that the aberrant erythropoiesis and megakaryopoiesis is due to the inability to generate MEPs from Nd1-S-overexpressing HSPCs. To test this possibility, the myeloid progenitor compartment of Nd1-S recipients was analyzed by flow cytometry. Unexpectedly, instead of the reduction of MEPs, an accumulation of phenotypic MEPs in the bone marrow was observed in Nd1-S recipients. Surprisingly, MEPs comprised 94.8% of total MPs in Nd1-S group, in contrast to 46.6% in WT mice (Fig. 4a, b). Therefore, the aberrant erythropoiesis and megakaryopoiesis caused by Nd1-S overexpression was not a result of inefficient generation of MEPs from HSPCs. Moreover, the Nd1-S MEPs appeared to have distorted shape and uneven stained nuclei (Fig. 4c). These phenotypic MEPs failed to form BFU-E colonies in CFU assay (Fig. 4d). Thus, Nd1-S overexpression in HSPCs induced accumulation of phenotypic MEPs with abnormal function.
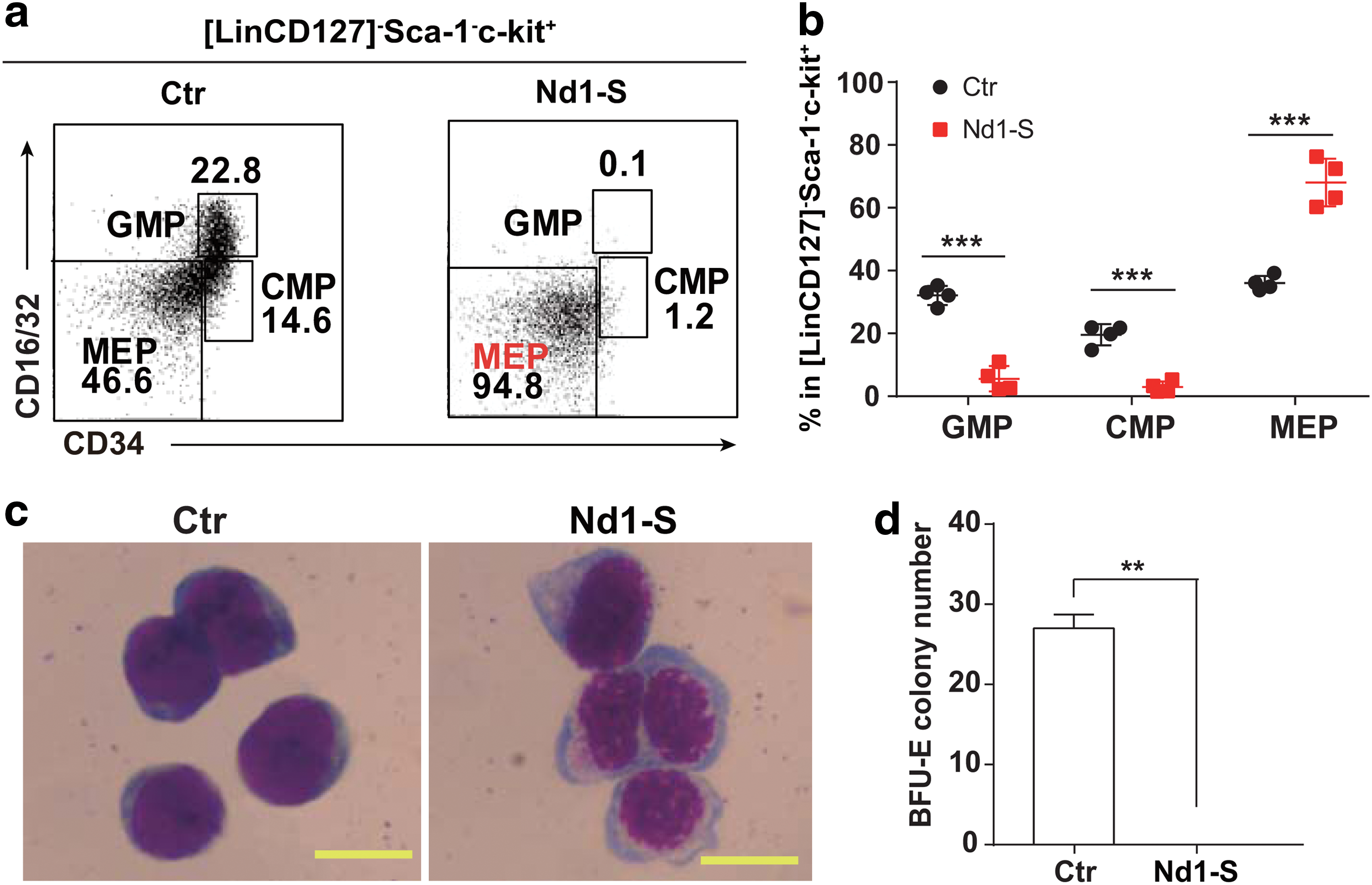
Enforced expression of Nd1-S leads to accumulation of phenotypic MEP compartment. All recipients were analyzed at week 16 posttransplantation.
Enforced expression of Nd1-S shows no impact on the cell cycle of MEPs
Since Nd1-S has been reported to regulate cell cycle progress in fibroblasts (Inoue et al., 2005), we further investigated whether Nd1-S overexpression altered the cell cycle status of MEPs. Unexpectedly, the cell cycle status of MEPs overexpressing Nd1-S was comparable with WT MEP controls (Fig. 5a, b). Collectively, our data demonstrated that the differentiation of phenotypic MEPs accumulated in the bone marrow of Nd1-S mice was impaired.
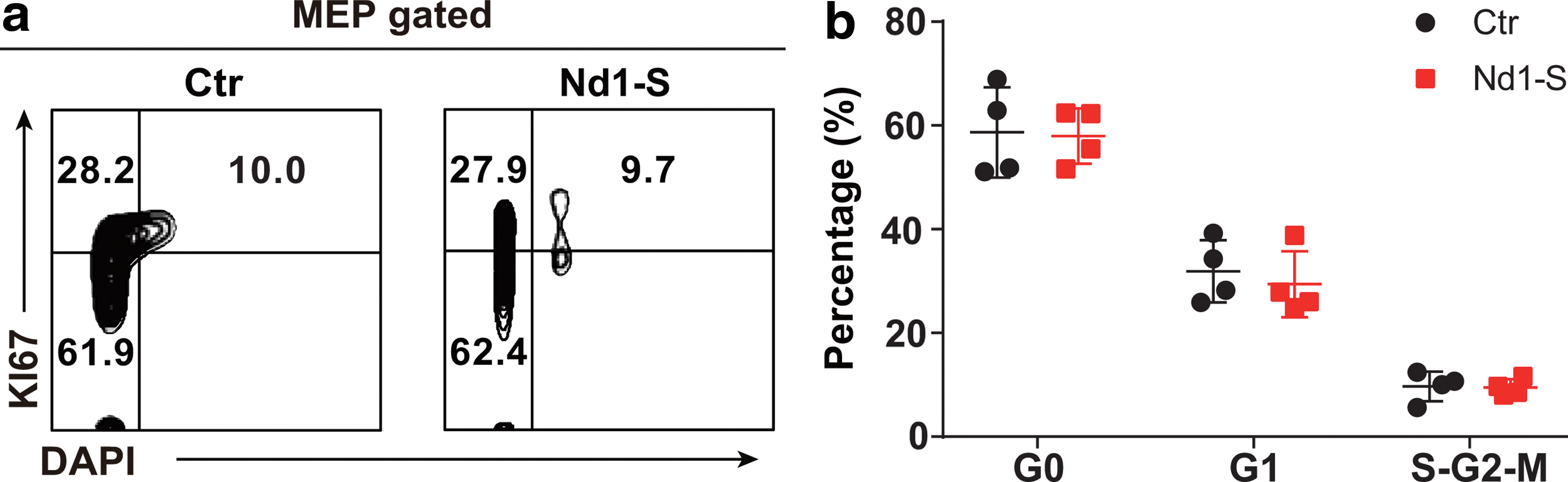
Enforced expression of Nd1-S shows no impact on the cell cycle of MEPs.
MEPs overexpressing Nd1-S demonstrated impaired pathways of erythroid differentiation and megakaryocyte development
To explore the molecular mechanism by which Nd1-S affects the function of MEPs, RNA-Seq was performed using the sorted MEPs from the Nd1-S mice and control mice. GSEA analysis of the RNA-Seq data showed that the signaling pathways for erythroid differentiation significantly decreased in Nd1-S-overexpressing cells (Fig. 6a, b, p < 0.05). Of note, Klf1 and Gata1, two well-recognized erythropoiesis regulators (Pevny et al., 1991, 1995; Nuez et al., 1995; Perkins et al., 1995; Gutierrez et al., 2008; Hopfer et al., 2012), were significantly downregulated in the Nd1-S-overexpressing MEPs. In addition, genes involved in megakaryocyte development and platelet production were also dramatically downregulated (Fig. 6c, d, p < 0.05). Notably, we found that overexpression of Nd1-S downregulated the expression level of Dock4 in the phenotypic MEP population, which was consistent with the observation that reduction of Dock4 induces erythroid dysplasia (Sundaravel et al., 2015). Besides the downregulated pathways mentioned above, the platelet homeostasis pathway was upregulated in the Nd1-S-overexpressing MEPs (Supplementary Fig. S2). Since Nd1 encodes an evolutionary conserved protein believed to serve as an E3 ubiquitin ligase adaptor protein, we observed that protein ubiquitination involved in ubiquitin-dependent protein pathway was downregulated (Supplementary Fig. S3). Collectively, we provide molecular evidence that overexpressing Nd1-S in MEPs impaired erythroid differentiation and megakaryocyte development.
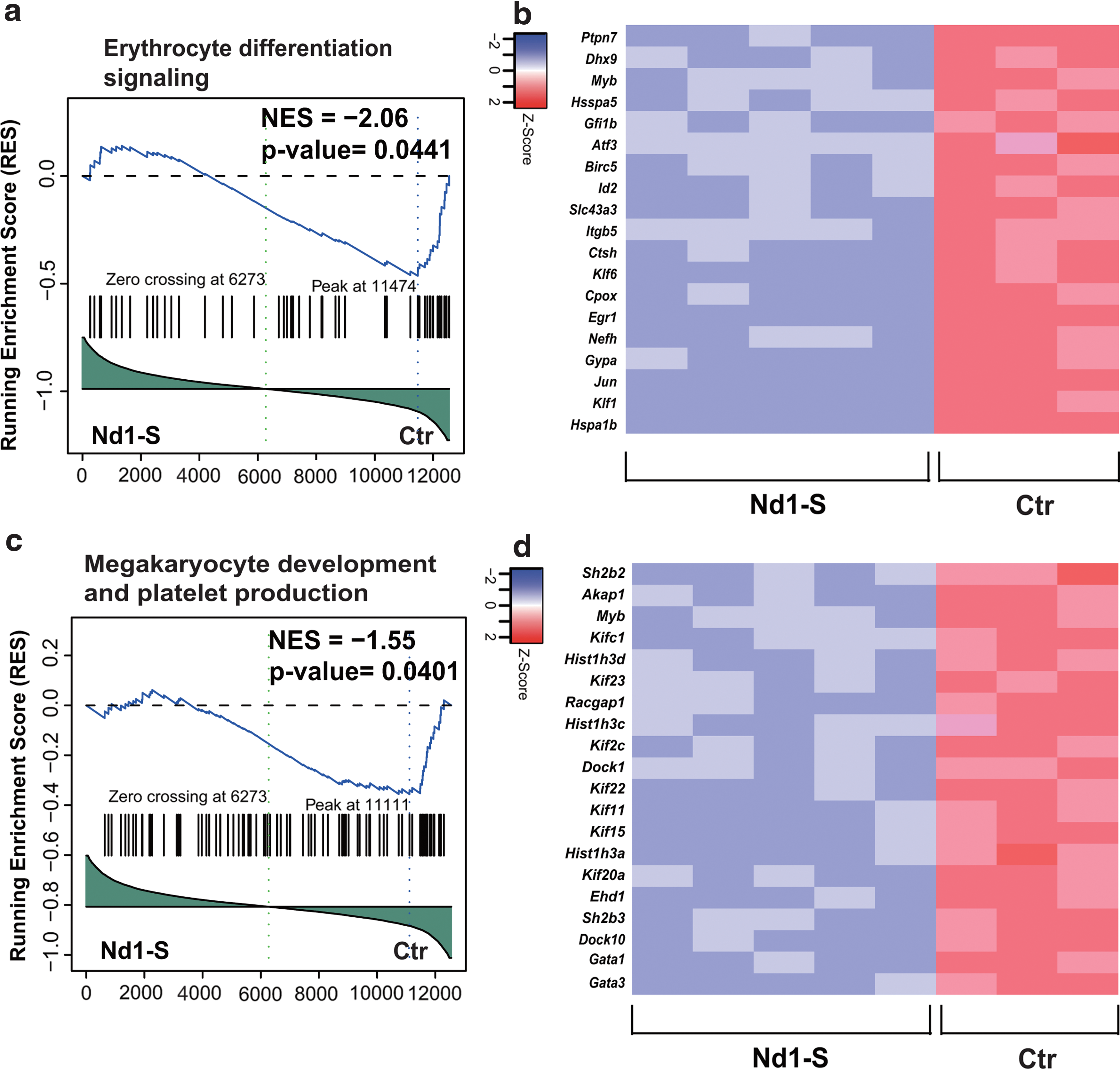
Transcriptome profile of MEPs from control and Nd1-S mice. MEPs were sorted from recipient mice 16 weeks posttransplantation. The 5 × 104 MEPs were used for each library preparation sample.
Discussion
In this study, we investigated the role of Nd1-S in hematopoiesis by overexpressing Nd1-S in murine HSPCs and followed transplantation assay. Enforced Nd1-S expression led to erythroid and megakaryocyte dysplasia in peripheral blood and bone marrow of Nd1-S mice, which are associated with the accumulation of abnormal phenotypic MEPs in Nd1-S mice. RNA-Seq of these phenotypic MEPs revealed deregulated pathways of erythroid differentiation and megakaryocyte development. We provide de novo evidence that Nd1-S plays a vital role in regulating differentiation of MEPs.
In this study, enforced Nd1-S expression in HSPCs leads to erythroid and megakaryocyte dysplasia in vivo. Intriguingly, 4.52% (10 of 221 samples tested) of hematopoietic and lymphoid cancer samples overexpressed IVNS1ABP (COSMIC database), which indicates that Nd1-S may contribute to disease development and progression of hematopoietic malignancies. We also observed that phenotypic MEP population accumulated in the Nd1-S mice, which failed to form B/CFU-Es in vitro. The impaired function of MEPs is likely due to the deregulated pathways of erythroid differentiation, megakaryocyte development, and platelet production.
In the previous report, overexpression of Nd1-S perturbs cell cycle progression of fibroblasts and arrests these cells at G1/S phase (Inoue et al., 2005) However, we found no evidence of cell cycle entry or delay of phenotypic MEPs in Nd1-S mice compared with wild-type counterparts. Nonetheless, we observed irregular shapes and distorted nuclei of these Nd1-S MEPs, which failed to form CFU-E in colony-forming assay. The dysfunction of phenotypic MEP differentiation in this study is more likely a consequence of deregulated pathways for normal erythroid differentiation and megakaryocyte development. For example, in our Nd1-S MEPs, the expression level of Flk1 is significantly reduced compared with control (Fig. 4b and Supplementary Table S1). Klf1 is essential for the definitive erythropoiesis, and its deficiency leads to embryonic lethality with fetal anemia, with β globin deficiency (Nuez et al., 1995; Perkins et al., 1995). Further study of the Klf1 null mice found that Klf1 regulates genes involved in hemoglobin metabolism and membrane stability, and Klf1−/− primitive cells show wrinkled morphology (Drissen et al., 2005). Similar to these report, we observed the morphologically abnormal erythrocytes in our Nd1-S mice (Fig. 1d), which suggests that Nd1-S may directly downregulate genes involving in differentiation pathways (Klf1, for example), which may cause the dysplasia in Nd1-S-overexpressing HSPC recipients. It would be interesting to investigate whether Nd1-S directly binds the regulatory elements of erythroid and megakaryocyte lineage regulators and tunes their expression levels. In addition, in our sequencing results, some Dock family members, such as Dock8, Dock5, Dock2, were upregulated while other members, such as Dock6, Dock4, Dock1, Dock10, were substantially attenuated in the Nd1-S MEPs. Dock4 has been reported to be actin filament stabilizer, and its reduction leads to erythroid dysplasia (Sundaravel et al., 2015). Since dock family proteins are guanine nucleotide exchange factors, which participate in intracellular signaling network, it deserves further investigation as to whether and how the dock family forms an interactive network to regulate erythroid and megakaryocyte differentiation (Cote and Vuori, 2002). What's more, the protein ubiquitination, involved in ubiquitin-dependent protein pathway was downregulated in Nd1-S-overexpressing MEPs, indicated that abnormal expression of Nd1-S also affects the protein ubiquitination process, since Nd1 encodes an evolutionary conserved protein believed to serve as an E3 ubiquitin ligase adaptor protein. Besides the downregulated pathways mentioned above, the platelet homeostasis pathway was upregulated in the Nd1-S-overexpressing MEPs. Platelet homeostasis reflects a balance between the production of platelets through cytoplasmic fragmentation of megakaryocytes (Patel et al., 2005). Therefore, the ectopic expression of Nd1-S also deregulated the platelet homeostasis, and indicated that Nd1-S played an important role in the development of erythrocytes, megakaryocytes, and platelets.
Authorship
Y.S.L., F.X.H., X.L.C., and J.D. designed the study, conducted the experiments, and analyzed the data. Y.X.L., X.D.H., Y.D., and D.Y. conducted the animal experiments and transcriptome analysis. J.D., T.J.W., and J.Y.W. supervised the study and wrote the article. X.Z.Z. corrected and approved the final version of the manuscript. All authors read and approved the final article.
Ethics Approval
All animal experiments were carried out in accordance with the Guide for the Care and Use of Laboratory Animals instructed by the Institutional Animal Care and Use Committee of Guangzhou Institutes of Biomedicine and Health (IACUC-GIBH) and the committee approved all experimental protocols (2012013).
Footnotes
Acknowledgments
This work was supported by the Natural Science Foundation of Guangdong Province, China (2014A030313138), Cooperation Program from Guangdong Natural Science Foundation (2014A030312012), Basic Research Program from the Guangdong Natural Science Foundation (S2013010016559, 2014A030313138, 2015A030313885), the Youth Innovation Promotion Association of the Chinese Academy of Sciences (2014327), the General Program from Guangzhou Scientific and Technological Project (201707010157), and the grants from the National Natural Science Foundation of China (Grant No. 31471117, 81470281, 31500948, and 31600948).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.