
Abstract
Emerging studies indicated that both long noncoding RNAs and micro-RNAs play crucial roles in the mediation of adipogenesis, which is closely linked to obesity-related diseases. However, the mechanisms of lncRNA-miRNAs coregulating in adipogenesis are still largely unknown. In this study, we determined that lncRNA growth arrest-specific 5 (GAS5) presented an opposite expression pattern with miR-21a-5p in 3T3-L1 adipocytes development. To explore the role of GAS5 in adipogenesis, pcDNA3.1-GAS5 expression vectors and GAS5-siRNAs were used to perform GAS5 overexpression and knockdown, respectively. Ectopic expression of GAS5 dramatically reduced miR-21a-5p level and suppressed the proliferation of 3T3-L1 preadipocytes, while silencing GAS5 slightly increased miR-21a-5p expression but had no significant influence on the cell viability. In addition, overexpression of GAS5 remarkably decreased the mRNA and protein levels of adipogenic marker genes, and resulted in a notable reduction of lipid accumulation. In contrast, overexpressing miR-21a-5p significantly facilitated differentiation of 3T3-L1 cells. By target gene prediction and luciferase reporter assay, we suggested that GAS5 might indirectly improve the expression of phosphatase and tensin homolog (PTEN) by repressing miR-21a-5p in a miRNA-based regulatory mechanism. Together, GAS5 plays a suppressive role in 3T3-L1 cells adipogenesis, which further highlights the importance of lncRNAs in adipogenesis.
Introduction
L
Adipogenesis is an extremely complicated and accurate physiological process, in which MAPK (Rosen et al., 2000), WNT/β-catenin (Kang et al., 2007), PI3K/Akt (Yu et al., 2008), and other crucial signal pathway and several central transcription factors such as peroxisome proliferator-activated receptor γ (PPARγ) (Brun and Spiegelman, 1997; Rosen et al., 1999; Tamori et al., 2002) and CCAAT/enhancer-binding protein α (C/EBPα) (Linhart et al., 2001) form a regulatory network. Abnormal adipogenesis and excessive adipocytes accumulation might trigger multiple types of obesity-related diseases, including hyperlipidemia, atherosclerosis, and chronic inflammation (Kahn et al., 2006; Sun and Karin, 2012; Shulman, 2014), which are prevalent health challenges and have attracted increasing attention of people all around the world. Mouse 3T3-L1 preadipocytes have been a typical model for adipogenesis study, which is mediated by an elaborate network for establishing the mature fat-cell phenotype. And adipogenesis of 3T3-L1 cells mainly proceeds into two stages called preadipocytes proliferation and adipogenic differentiation (Rosen and Spiegelman, 2000).
Recently, emerging evidence indicates that lncRNAs are gradually considered as important regulatory elements and play a critical role in adipogenesis (Sun et al., 2013; Xiao et al., 2015; Li et al., 2016). GAS5 is a growth arrest- and starvation-associated repressor of glucocorticoid receptor (Tomoshige et al., 2010), and was originally isolated from mouse NIH 3T3 cell that is one of the most widely used preadipocyte cell lines in mice as well as 3T3-L1 cells, and applied for studying adipocytes proliferation, differentiation, and regulation of relative gene expression during adipogenesis. Although GAS5 has been regarded as a suppressor of malignancy, the underlying effect of GAS5 on adipogenesis is still unknown. In this study, we found that GAS5 might indirectly promote phosphatase and tensin homolog (PTEN) expression through sponging miR-21a-5p, thereby repressing 3T3-L1 cells proliferation and adipogenic differentiation. Together, our results further implied a crucial role of lncRNA GAS5 in adipogenesis, which expanded our knowledge about molecular mechanism of regulating adipogenesis.
Materials and Methods
Cell culture and adipogenic differentiation
Mouse 3T3-L1 preadipocytes and HeLa cells were purchased from Stem Cell Bank, Chinese academy of sciences. The cells were incubated in 5% CO2 at 37°C and were cultured in high-glucose Dulbecco's Modified Eagle Medium (DMEM) (Gibco, Carlsbad, CA) with 10% fetal bovine serum (FBS) (Gibco) before differentiation. When cultures reached ∼80% confluence, cells were digested by incubation with 0.25% trypsin for 3 min at room temperature, and then seeded in 96-/12-well plates. For adipogenic differentiation, postconfluent 3T3-L1 cells were induced by differentiation medium (DM) containing 10% FBS, 0.5 mM 3-isobutyl-1-methylxanthine, 1 μM dexamethasone, and 1.7 μM insulin for 2 days. The medium was then replaced with DMEM containing 10% FBS and 1.7 μM insulin for additional 2 days.
Plasmid construction and luciferase reporter assay
The transcript covering the most part of mouse lncRNA GAS5 sequence (NR_002840.2 and ENSMUST00000161229.7) was amplified by high-fidelity PCR using primers GAS5-C-F and GAS5-C-R (Table 1), and then cloned into the expression vector pcDNA3.1 (Promega, Madison, WI). This recombinant vector was named pcDNA3.1-GAS5. To construct luciferase reporter plasmids, the miR-21a-5p targeting sites within the mouse lncRNA GAS5 and PTEN 3′ UTR sequence were established using RNAhybrid 2.2. The lncRNA GAS5 and PTEN-3′ UTR fragments, containing a putative miR-21a-5p targeting site, were amplified by PCR and cloned into the SacI-XbaI of the pmirGLO vector multiple cloning sites (MCSs) (Promega). And the two pairs of primers for PCR were GAS5-miR21-BS-F/GAS5-miR21-BS-R and PTEN-3′ UTR-F/PTEN-3′ UTR-R (Table 1). All PCR products were verified by DNA sequencing, and the recombinant reporter vectors were named pmirGLO-GAS5-BS and pmirGLO-PTEN-3′ UTR, respectively. For luciferase reporter assays, HeLa cells were seeded into 96-well plates in triplicate. After cell density reached 70%, pmirGLO-GAS5-BS or pmirGLO-PTEN-3′ UTR was cotransfected with the synthetic miR-21a-5p mimics (50 nM; catalog no.: miR10000530-1-5) into HeLa cells. PmirGLO or mimics negative control (NC) (50 nM; catalog no.: miR01201-1-5) was cotransfected as an internal control. At 48 h post-transfection, firefly luciferase (Luc2) activity was measured and normalized to the Renilla luciferase (hRluc-neo) activity according to the Dual-Luciferase® Reporter Assay System (Promega).
The underlined nucleotides represent enzyme cutting sites.
F, forward; qRT-PCR, quantitative real-time PCR; R, reverse.
Transfection of plasmids, siRNAs, and miRNA mimics
When 3T3-L1 preadipocytes reached ∼70% confluence, GAS5 overexpression plasmids (pcDNA3.1-GAS5) were transfected into the cells using Lipofectamine 3000 (Invitrogen), while siRNA-GAS5-1 (100 nM; catalog no.: siB1356203228), siRNA-GAS5-2 (100 nM; catalog no.: siB1356203300), siRNA-GAS5-3 (100 nM; catalog no.: siB1356203319), siRNA-NC (100 nM; catalog no.: siN05815122147), miR-21a-5p mimics (50 nM), or mimics NC (50 nM) (all purchased from RiboBio, Guangzhou, China) were transfected into cells using ribo Fect™ CP Transfection Kit (RiboBio) according to the manufacturer's instructions. After 2 days, postconfluent cells were induced to differentiate with DM.
Cell proliferation assays and EdU staining
Proliferation of cells was monitored by colorimetric assay using Cell Counting Kit-8 (CCK-8; Beyotime, Shanghai, China), which is based on the cleavage of the tetrazolium salt WST-8 by mitochondrial dehydrogenase in viable cells. In brief, 100 μL of 3T3-L1 cell suspension from each group (2 × 103 cells/well) was seeded into 96-well plates and preincubated for 12 h. Then, 10 μL of CCK-8 solution was added to each well incubated for 1 h, and the absorbance at 450 nm was measured every 24 h for 5 days using a microplate reader. Each experiment sets six individual technical repetitions, and results represent the mean of all measurements. For EdU staining, EdU solution (RiboBio) was diluted 1:1000 in cell medium. 3T3-L1 cells were treated with 50 μM EdU medium 48 h after transfection and incubated for further 2 h. EdU staining was done according to the manufacturer' protocol. Images were captured using a Nikon TE2000 microscope (Nikon, Tokyo, Japan).
Total RNA isolation and quantitative real-time PCR
Total RNA was extracted from tissues or 3T3-L1 cells using RNAiso Plus reagent (TaKaRa, Japan) according to the manufacturer's protocol. RNA quality and concentration were determined by 1.5% gel electrophoresis and NanoDrop 2000 UV-Vis Spectrophotometer (Thermo, Waltham, MA). And then, cDNA was reverse transcribed using the PrimeScript RT reagent kit (TaKaRa). The Bulge-loop™ miRNA qRT-PCR Primer Sets (one RT primer and a pair of quantitative PCR primers for each set, MQP-0101 and MQP-0201) specific for miR-21a-5p and U6 small nuclear RNA were designed by RiboBio (Guangzhou, China). Quantitative real-time PCR (qRT-PCR) was performed to analyze mRNA and miR-21a-5p expression as described previously (Li et al., 2017) with normalization to β-actin for mRNAs and to U6 for miR-21a-5p.
Western blotting and antibodies
Cells were harvested, washed with 1× phosphate-buffered saline (PBS), and lysed in radio immunoprecipitation assay (RIPA) buffer (Beyotime) with protease inhibitors (Sigma). The protein content of different treatment groups was quantified by the bicinchoninic acid (BCA) method. After centrifugation, the supernatant was boiled in sodium dodecyl sulfate (SDS) loading buffer (Beyotime) for ∼5–10 min, 10 μg total protein was electrophoresed in 12% SDS-polyacrylamide gels and transferred onto polyvinylidene difluoride (PVDF) membranes (Millipore, Bedford, MA). And then, blocking and antibodies incubation were performed as described previously (Ning et al., 2017). The membranes were incubated with chemiluminescence reagents (Beyotime), and signal images were captured by a GelDoc XR+ gel imaging system (Bio-Rad). The antibodies used were anti-PPARγ (dilution 1:1000; Abcam, Cambridge, United Kingdom), anti-FABP4 (dilution 1:1000; Cell Signaling Technology, MA), anti-PTEN (dilution 1:800; Sangon, Shanghai, China), anti-β-actin (dilution 1:1000; Sangon), and horseradish peroxidase-conjugated goat anti-rabbit IgG (dilution 1:5000; Sangon).
Oil Red O staining and triglyceride assay
Adipocytes were washed twice with PBS, and then fixed with 4% paraformaldehyde for 30 min. Oil Red O (ORO) working solution was prepared and freshly filtered before staining. After fixation, the cells were washed thrice again with PBS and stained with 1% ORO solution for 30 min at room temperature. Then, the stained intracellular lipid droplets were washed twice with PBS and visualized using a phase contrast microscope. For triglyceride assay, ORO was eluted from the stained cells with isopropanol and quantified by measuring the optical density (OD) values at 510 nm.
Statistical analysis
Data were analyzed by GraphPad Prism 6, and the results are presented as mean ± standard error of the mean based on at least three separate experiments. Student's t-test was used for two parametric groups. One-way ANOVA followed by Duncan's test and Bonferroni correction was performed for multiple comparisons. p < 0.05 was considered statistically significant.
Results
GAS5 expression levels of mouse tissues and 3T3-L1 cells
To investigate the GAS5 expression levels of mouse different tissues, we extracted the total RNA from six tissues of 4-week-old mice, and the qRT-PCR result showed that the relative GAS5 expression level was higher in heart, muscle, and fat than in lung and liver (Fig. 1A). We subsequently examined the GAS5 expression levels of 3T3-L1 cells under different cultivation conditions. As shown in Figure 1B and C, the GAS5 expression levels were significantly upregulated in 3T3-L1 cells at a high cell density (full confluence) and induced by serum starvation for 12 h.

GAS5 expression levels of mouse tissues and 3T3-L1 cells.
GAS5 might function as a ceRNA for miR-21a-5p
It had demonstrated that GAS5 and miR-21a-5p could reciprocally inhibit each other in human breast cancer cells (Zhang et al., 2013). Thus, we identified the complementary sequence between GAS5 and miR-21a-5p in mouse using RNAhybrid to examine whether the regulation exists in mouse as well. As shown in Figure 2A, the mouse GAS5 gene contains the target site for miR-21a-5p in its exon 1. To manifest the relationship between GAS5 and miR-21a-5p, the fragment of GAS5 containing the miR-21a-5p binding site was cloned into the MCS of the pmirGLO vector (Fig. 2B). The dual-luciferase reporter assay showed that the relative Luc2 activity was remarkably reduced when cotransfection of pmirGLO-GAS5-BS and miR-21a-5p mimics but remained unchanged in other combinations (Fig. 2C). Based on this verification, we also detected the expression of miR-21a-5p under those two stresses. In contrast to GAS5 (Fig. 1B, C), Bulge-loop qRT-PCR showed that cell–cell contact inhibition and serum starvation significantly decreased the miR-21a-5p expression compared with the corresponding NC groups (Fig. 2D, E).
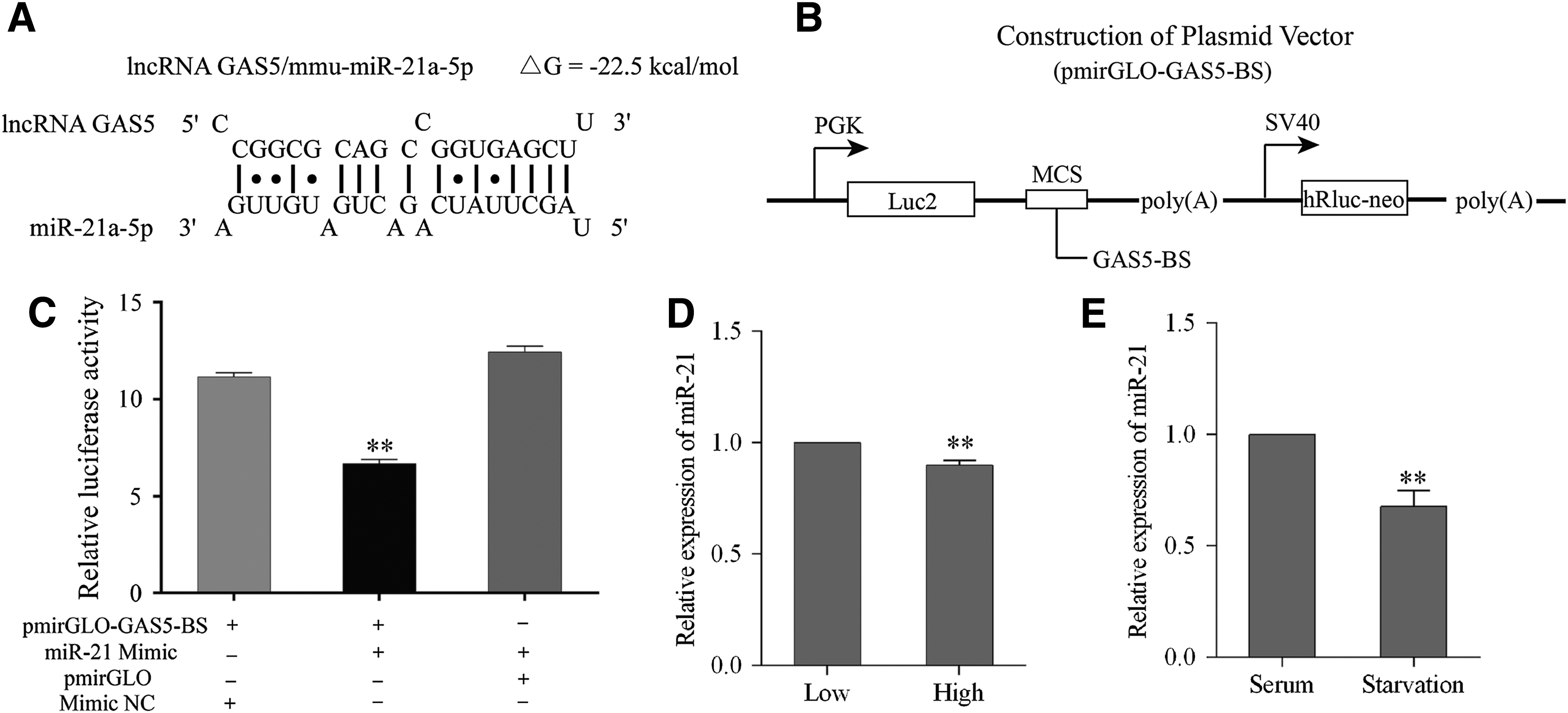
GAS5 might function as a ceRNA for miR-21a-5p.
Evaluation of pcDNA3.1-GAS5 and si-GAS5-1/2/3 transfection efficiency and effects on miR-21a-5p expression
To evaluate the transfection efficiency, we transfected 3T3-L1 cells with pcDNA3.1-GAS5 expression vector and synthetic siRNAs of GAS5 (si-GAS5-1/2/3), respectively. As shown in Figure 3A, GAS5 was significantly overexpressed by almost sevenfold after pcDNA3.1-GAS5 transfection. In contrast, GAS5 was effectively inhibited when si-GAS5-1/2/3 were transfected into 3T3-L1 cells compared with si-NC (Fig. 3B). Next, we detected the expression of miR-21a-5p in different groups using stem-loop qRT-PCR. As shown in Figure 3C, miR-21a-5p expression in 3T3-L1 cells transfected with pcDNA3.1-GAS5 was ∼50% of the empty vector group. However, transfection with si-GAS5-1/2/3 slightly enhanced the miR-21a-5p level, and only the difference between si-GAS5-1 and si-NC was significant (Fig. 3D).
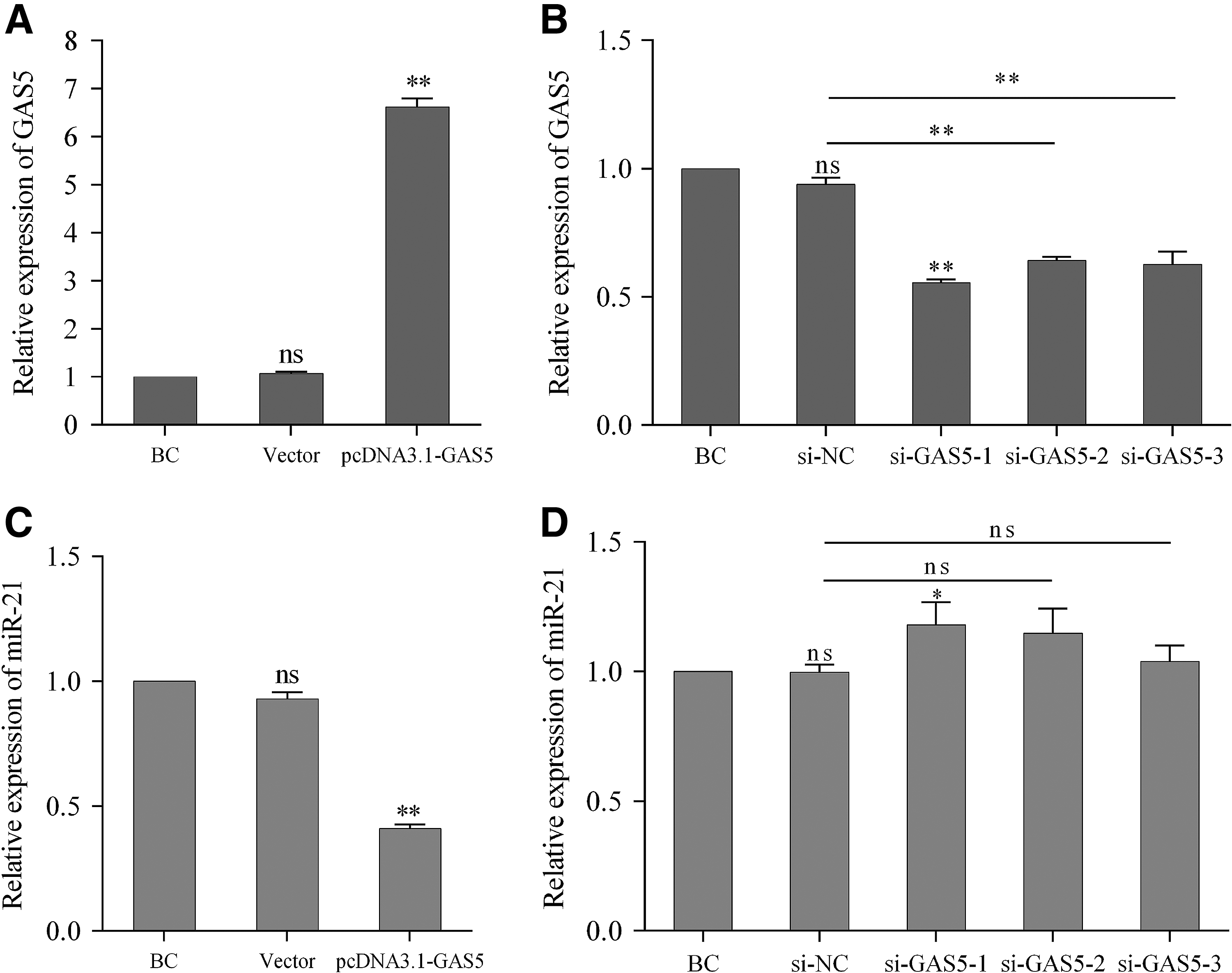
Evaluation of pcDNA3.1-GAS5 and si-GAS5-1/2/3 transfection efficiency and effects on miR-21a-5p expression. 3T3-L1 cells were transfected with pcDNA3.1-GAS5, pcDNA3.1, synthesized si-GAS5-1/2/3, or si-NC. Expression of GAS5
Overexpressing GAS5 suppresses 3T3-L1 cells proliferation
We then investigated the effects of GAS5 on 3T3-L1 cells proliferation through CCK-8 assay and EdU staining. Cell proliferation assay by CCK-8 revealed that overexpressing GAS5 remarkably decreased the OD value of 3T3-L1 cells on days 3, 4, and 5 after seeding, while GAS5 knockdown did not increase the cell viability compared with si-NC (Fig. 4A). Similarly, overexpressing GAS5 strongly decreased the cell number and the ratio of EdU positive cells on 2 days post-transfection (Fig. 4B).
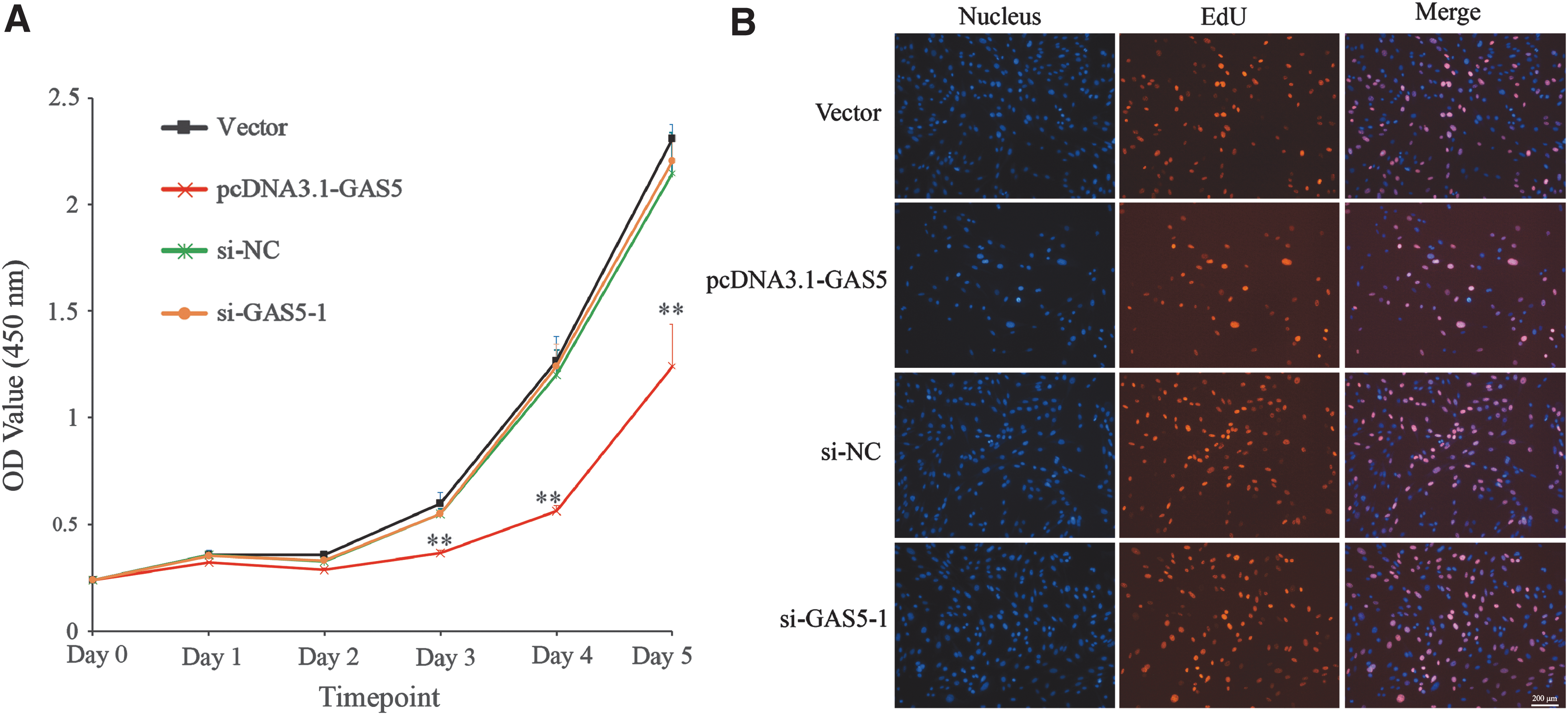
Overexpressing GAS5 suppresses 3T3-L1 cells proliferation. Cell proliferation assays of different groups were measured by CCK-8
Expression of GAS5 and miR-21a-5p during 3T3-L1 cells adipogenic differentiation
We first quantified the expression of GAS5 and miR-21a-5p after induction of differentiation. As shown in Figure 5A, the relative expression of GAS5 was upregulated and peaked on day 1 after induction of adipogenic differentiation. And it decreased back to baseline from day 1 to day 3 and slightly rose on day 4. In contrast, the miR-21a-5p level showed an opposite tendency. Then, 3T3-L1 cells were transfected with pcDNA3.1-GAS5, miR-21a-5p mimics, and corresponding NC separately, and induced by differentiation at 48 h post-transfection. The expression of GAS5 was significantly upregulated in 3T3-L1 cells transfected with pcDNA3.1-GAS5 compared with empty vector and obviously downregulated in cells transfected with miR-21a-5p mimics compared with mimics NC during the differentiation (Fig. 5B). In addition, miR-21a-5p was significantly overexpressed under miR-21a-5p mimics transfection but weakened after pcDNA3.1-GAS5 transfection (Fig. 5C).
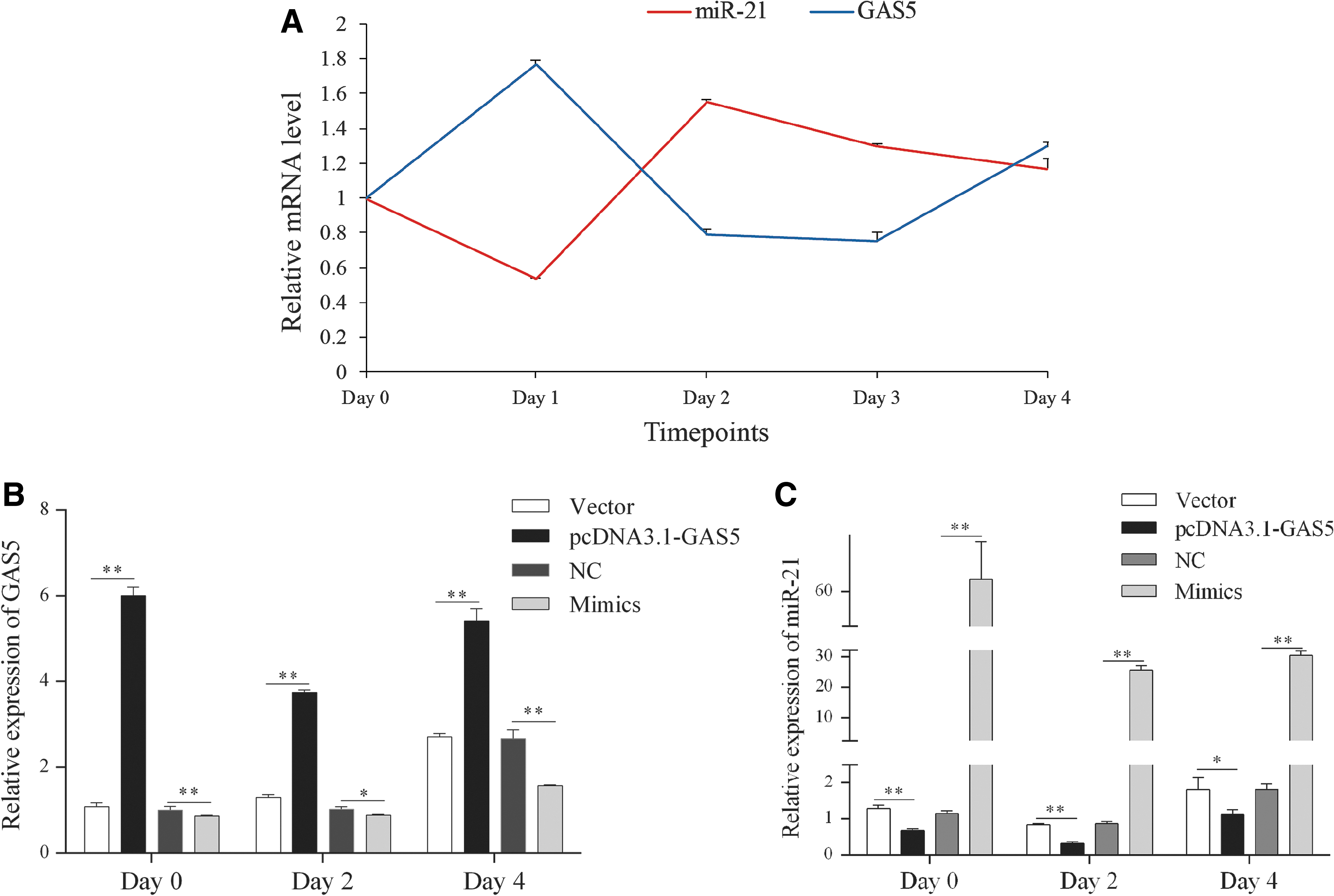
Expression of GAS5 and miR-21a-5p during 3T3-L1 cells adipogenic differentiation.
Effects of GAS5 and miR-21a-5p on lipid accumulation
Next, we investigated the effects of GAS5 and miR-21a-5p on lipid accumulation using ORO staining and a triglyceride extraction assay. And we found that overexpressing GAS5 significantly inhibited lipid droplet formation and triglyceride synthesis (Fig. 6A, B). In contrast, photomicrograph and quantitative analysis of ORO on day 4 of differentiation showed a significant increase of lipid droplets accumulation in miR-21a-5p mimics group compared with NC group (Fig. 6A, B).
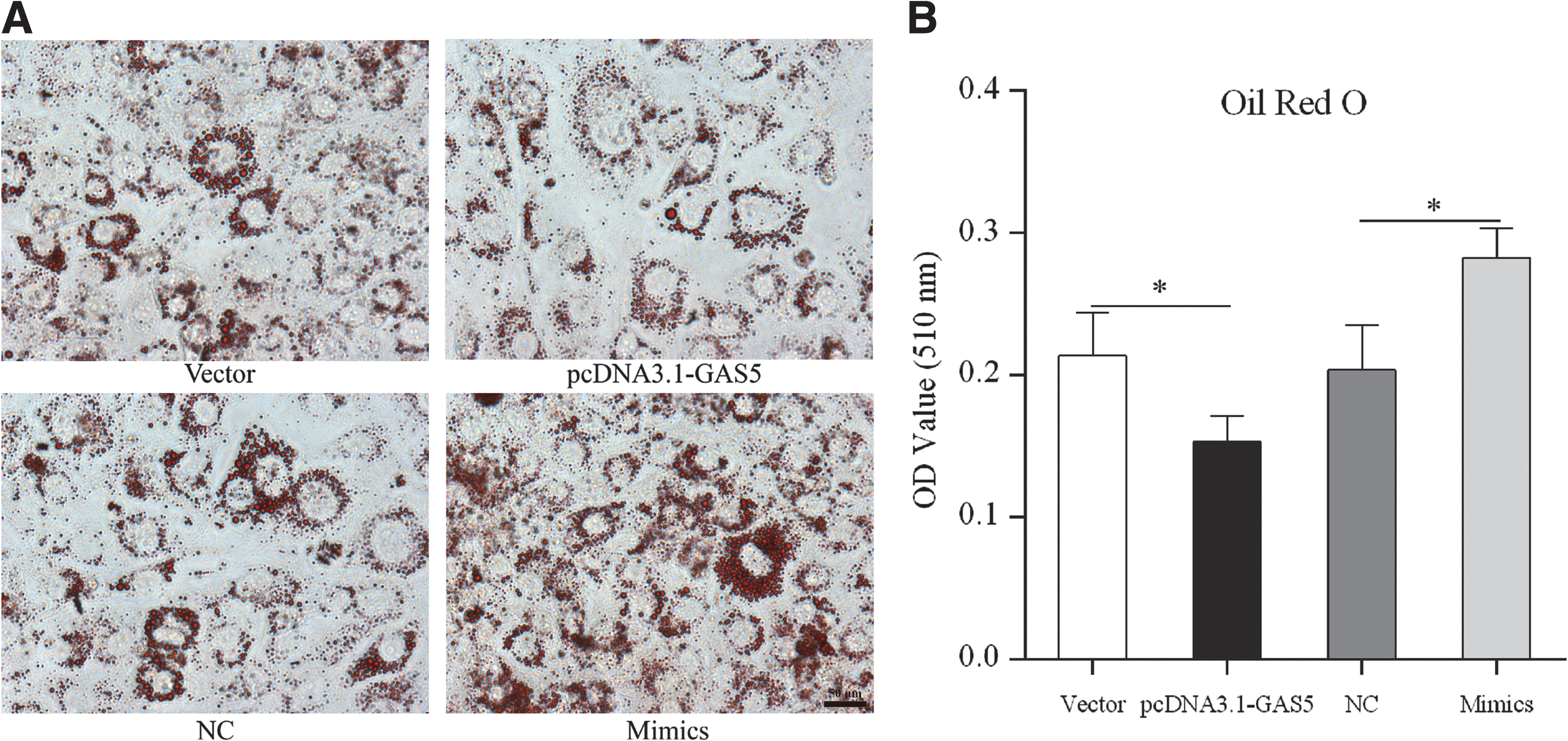
Effects of GAS5 and miR-21a-5p on lipid accumulation. Photomicrograph
Effects of GAS5 and miR-21a-5p on the expression of adipogenic genes
To further explore the influence of GAS5 on adipocytes differentiation, we detected the mRNA (on days 0, 2, and 4 during differentiation) and protein (on day 4 after differentiation) levels of crucial genes related to adipogenic differentiation, fatty acid binding protein 4 (FABP4), lipoprotein lipase (LPL), PPARγ, and fatty acid synthase (FAS). As shown in Figure 7A–D, the mRNA levels of FABP4, LPL, PPARγ, and FAS were remarkably downregulated in cells transfected with pcDNA3.1-GAS5 compared with NC. However, the mRNA levels of FABP4, LPL, and PPARγ markedly increased in miR-21a-5p mimics group compared with NC group. Although there was no significant change of FAS mRNA on day 0 between miR-21a-5p mimics group and NC group, a significant increase was detected on days 2 and 4 after differentiation (Fig. 7D). Consistent with the mRNA expression, the expression alteration of FABP4 and PPARγ on day 4 after differentiation was also presented by Western blotting analysis (Fig. 8A–D).
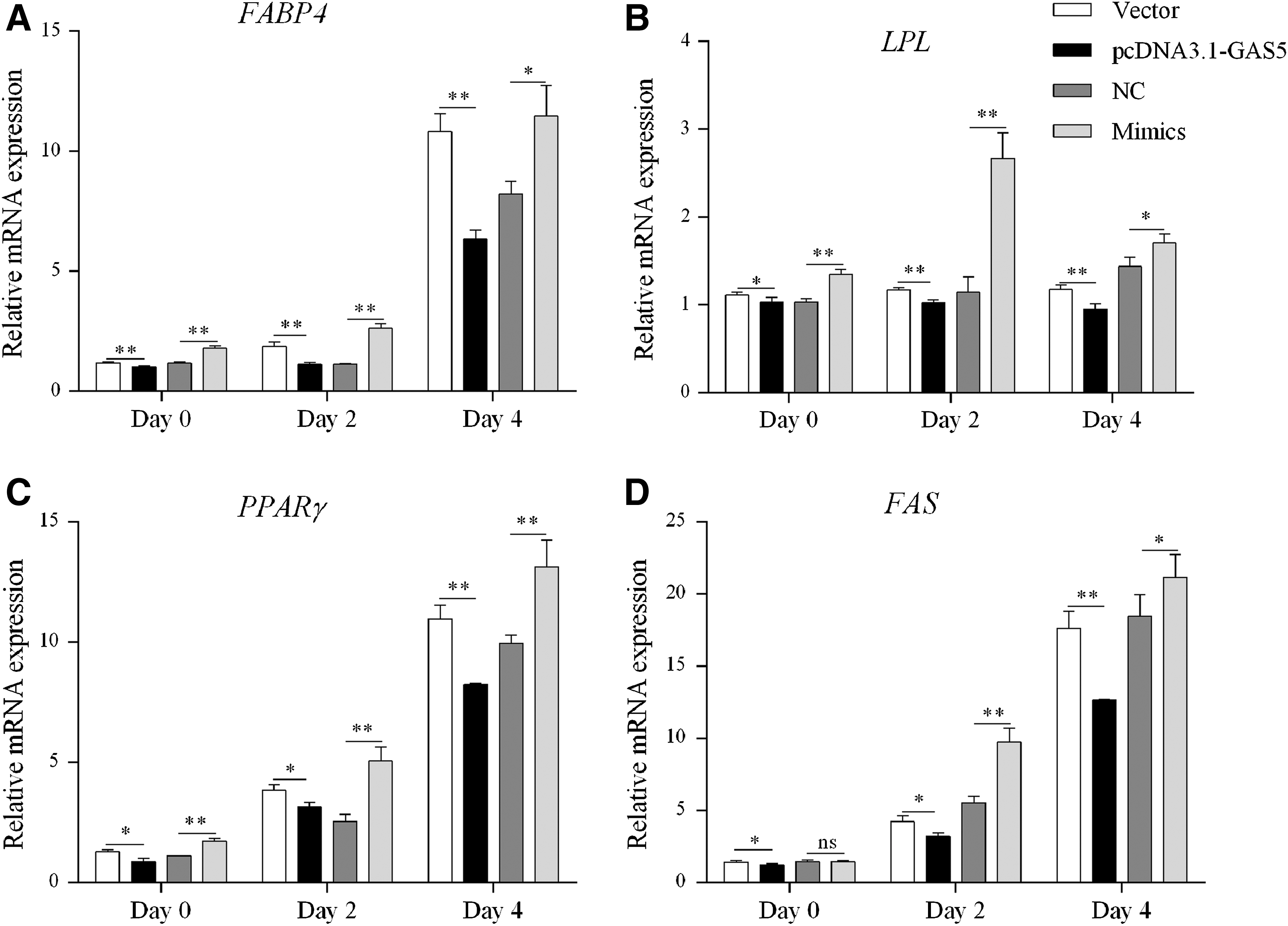
Effects of GAS5 and miR-21a-5p on the mRNA expression of adipogenic genes. The mRNA expression of FABP4

Effects of GAS5 and miR-21a-5p on the protein expression of adipogenic genes.
GAS5 might indirectly alleviate impaired PTEN expression by suppressing miR-21a-5p
To investigate the functional mechanism of GAS5, we further predicted target genes of miR-21a-5p by TargetScan 6.2 (
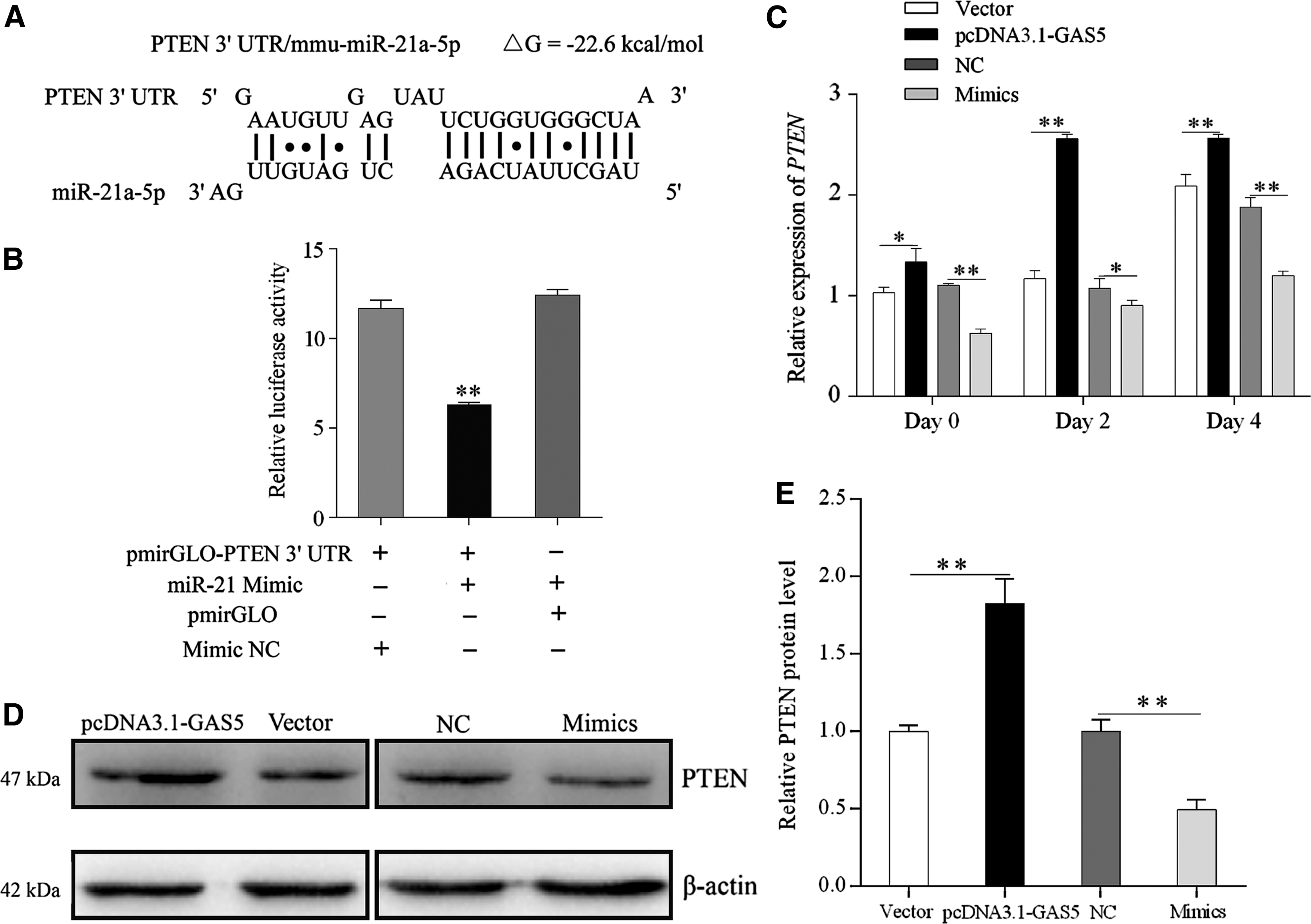
GAS5 might indirectly alleviate impaired PTEN expression by suppressing miR-21a-5p.
Discussion
In recent years, an increasing number of evidence have demonstrated that lncRNAs could participate in mediating adipogenesis through multiple ways. For instance, 10 candidate lncRNAs have been identified and validated to have biological function during adipogenesis by deep RNA-Seq and RNAi-mediated loss of function assays (Sun et al., 2013). Li et al. (2016) have reported that ADNCR was the most downregulated lncRNA during bovine preadipocytes differentiation and could augment SIRT1 expression by sponging miR-204, thereby inhibiting adipogenesis. GAS5 was originally identified and presented a relatively high expression level in mouse NIH 3T3 fibroblasts. We found that GAS5 could be induced by cell–cell contact inhibition and serum starvation in 3T3-L1 cells, which was consistent with a previous study (Schneider et al., 1988). In addition, 3T3-L1 cell line is one of the most widely applied mice adipocyte models as well as NIH 3T3 fibroblasts, and the relationship between GAS5 and mechanisms of 3T3-L1 adipogenesis remains unknown. In this study, gain and loss of function assays were performed to investigate the regulatory effect and potential mechanism of GAS5 during 3T3-L1 cells adipogenesis.
A previous study had demonstrated that GAS5 could function as a ceRNA for miR-21 and suppress proliferation of human breast cancer cells (Zhang et al., 2013). Although sequence conservation of lncRNAs was poor, certain partial sequences and function of lncRNA across different species presented relatively high conservation (Mitchell et al., 2009; Ulitsky et al., 2011). Our prediction and dual-luciferase reporter assay also showed that GAS5 might be a ceRNA for miR-21a-5p in murine 3T3-L1 cells. GAS5 was previously described to present a low expression level and played a strong tumor-suppressive role in multiple tumor tissues (Guo et al., 2015; Pickard and Williams, 2016; Wang et al., 2017). Meanwhile, miR-21a-5p, as one of the earliest identified miRNAs, had been considered as an oncogene (Meng et al., 2007; Mjd et al., 2017). Therefore, we explored the effect of GAS5 on miR-21a-5p and 3T3-L1 cells proliferation, and our results indicated that overexpressing GAS5 could significantly inhibit miR-21a-5p expression and strongly repress cells viability. Out of our expectation, knocking down GAS5 did not remarkably promote cells proliferation, which might be caused by the relatively low endogenous expression level of GAS5 compared with miR-21a-5p.
Recent studies also revealed that miR-21a-5p intimately involved in adipogenesis and accelerated adipogenic differentiation through several signal pathways such as TGF-β/Smad and ERK/MAPK (Kim et al., 2009; Mei et al., 2013; An et al., 2016). Therefore, we wanted to investigate whether GAS5 could indirectly influence 3T3-L1 cells differentiation by repressing miR-21a-5p. For this purpose, we first quantified the expression of GAS5 and miR-21a-5p during 3T3-L1 cells differentiation. As shown in Figure 5A, the expression pattern of GAS5 was opposite to that of miR-21a-5p. Our data further showed that overexpressing GAS5 significantly inhibited lipid droplet formation and marker gene expression levels during adipogenic differentiation, which is in contrast to overexpression of miR-21a-5p. Together, these results indicated that GAS5 might inhibit 3T3-L1 cells adipogenic differentiation by operating as a sponge for miR-21a-5p.
To investigate the concrete regulatory mechanism of GAS5, we further predicted target genes of miR-21a-5p, and found that PTEN contains the target site for miR-21a-5p in its 3′ UTR. PTEN was originally characterized as a tumor suppressor (Salmena et al., 2008), frequently mutating or deleting in human cancers, and was an important plasma membrane lipid phosphatase that attenuated the activity of phosphatidylinositol 3-kinase type 1 (PI3K) by dephosphorylating the 3′ position of phosphatidylinositol-3, 4, 5-trisphosphate (PIP3) (Maehama and Dixon, 1998; Chalhoub and Baker, 2009). Evidence indicated that ectopic expression of PTEN restrained the activity of insulin-stimulated PI3K/Akt signaling pathway, glucose uptake, and resulted in apparent insulin resistance (Nakashima et al., 2000), while silencing PTEN obviously enhanced Akt phosphorylation and glucose transport in 3T3-L1 adipocytes (Tang et al., 2005). Furthermore, our previous study had provided evidence for the negative regulation of PTEN on 3T3-L1 cells adipogenic differentiation (Li et al., 2017). Thus, in this study, we further determined the expression of PTEN on days 0, 2, and 4 during differentiation using qRT-PCR and Western blotting. Together, these results implied a miRNA-based function of GAS5, and indicated that lncRNA GAS5 might indirectly alleviate impaired PTEN expression by suppressing miR-21a-5p.
Conclusion
In summary, as a negative regulator of proliferation and adipogenic differentiation, lncRNA GAS5 could be induced by cell–cell contact inhibition and serum starvation in 3T3-L1 cells. In addition, lncRNA GAS5 might indirectly promote PTEN expression and finally suppress 3T3-L1 cells adipogenesis by sponging miR-21a-5p, further highlighting the importance of lncRNA in adipogenesis.
Footnotes
Acknowledgments
This research was supported by grants from the Science and Technology Cooperation Project of Sichuan Province and Chinese Academy of Science (2017JZ0025), the National Natural Science Foundation of China (31772576, 31601919, and 31522055), the Program for Innovative Research Team of Sichuan Province (2015TD0012), the Science and Technology Support Program of Sichuan (2016NYZ0042), and the Science and Technology Major Projects of Sichuan (2017NZDZX0002).
Disclosure Statement
No competing financial interests exist.