
Abstract
The AMP deaminase 1 (AMPD1) gene plays an important role in purine nucleotide interconversion and energy metabolism. In this study, two novel single nucleotide polymorphisms (SNPs) (g.-626 G > A and g.-566 A>G) were detected in the proximal promoter region of the AMPD1 gene. The Chinese indigenous pig breeds (Meishan and Tongcheng) had higher frequencies of the G and A alleles than Western meat-type breeds (Landrace and Large White) at the g.-626 G > A and g.-566 A>G loci. The transcriptional activity of the AMPD1 promoter carrying the haplotype H1 (A−626G−566) was significantly (p < 0.05) higher than that of the haplotype H2 (G−626A−566). In addition, pigs with the haplotype combination H1H1 had significantly (p < 0.05) higher mRNA expression levels of the AMPD1 gene than those with haplotype combinations H1H2 and H2H2 in two different skeletal muscles. Association analyses revealed that the pigs with the haplotype combination H1H1 had significantly higher lean meat percentage values but lower average backfat thickness (ABT, cm), buttock fat thickness (cm), and thorax-waist fat thickness (cm) values than the pigs with the haplotype combinations H1H2 and H2H2. These findings demonstrate that the two novel SNPs in the promoter region of the AMPD1 gene have significant associations with pig fat deposition traits.
Introduction
T
A C/T transition (c.34 C > T) is found in the coding region at nucleotide 34 (codon 12 in exon 2). The C/T transition is a nonsense mutation and produced a severely truncated AMPD peptide in the skeletal muscle and heart of patients (Morisaki et al., 1992; Yacoub et al., 2004; Fedotovskaya et al., 2013). A single nucleotide polymorphyism (SNP) (c.426 T > C) in the pig AMPD1 gene is significantly associated with the meat quality traits in 400 commercial cross-bred pigs (Zhang et al., 2014). The pig AMPD1 gene has been mapped to SSC4q1.6-q2.3 (Stratil et al., 2000), where many significant quantitative trait loci affecting carcass traits have been identified (Silva et al., 2011; Walling et al., 2000). Reducing AMP deaminase activity in vitro system extends anaerobic glycolysis and pH decline in pig longissimus muscle samples. The muscles of RN (-) genotype pigs contain lower AMP deaminase activity than wild-type muscle samples, and they may contribute to the low ultimate pH in RN-pork (England et al., 2015).
In our previous study, the pig AMPD1 gene was cloned and characterized. The AMPD1 gene is expressed predominantly in skeletal muscle and is upregulated during skeletal muscle development (Wang et al., 2008), suggesting that it may have a relationship with skeletal muscle development. In addition, proteomics analyses were performed in the longissimus dorsi muscle of both Large White and Meishan pigs. The AMPD1 protein exhibits considerable differences between the different skeletal muscle development stages of the two breeds (Xu et al., 2009, 2012). Therefore, we focused on the pig AMPD1 gene that could be a candidate gene relating to pig carcass traits. In this study, we investigated the polymorphisms in the promoter region of the AMPD1 gene and performed an association analysis in our Large White × Meishan pig resource family with carcass traits.
Materials and Methods
Ethics statement
All research involving animals was conducted according to the Regulations for the Administration of Affairs Concerning Experimental Animals (Ministry of Science and Technology, China, revised in June 2004) and approved by the Institutional Animal Care and Use Committee at the College of Animal Science and Technology, Sichuan Agricultural University, Sichuan, China under permit No. DKY-B20110807.
Animals and traits
The two F2 resource populations used in the association analysis were derived from the intercross of three Large White boars and seven Meishan sows. One population consisted of 140 F2 animals derived by mating 5 males to 16 females of the F1 generation, and the other population consisted of 194 F2 animals derived by mating 5 males to 21 females of the F1 generation (Wang et al., 2014a). They were fed twice daily with diets formulated according to age based on a standardized feeding regimen and free access to water. The average live weight at slaughter was 87.0 ± 7.07 kg. The F2 pigs were slaughtered in 2000 and 2003 following a common protocol (Zhang et al., 2007).
The estimated values of important economic traits involving fat deposition and carcass traits were obtained as follows: fat meat percentage, lean meat percentage (LMP), ratio of lean to fat, shoulder fat thickness (cm), six to seventh rib fat thickness (cm), thorax-waist fat thickness (TFT, cm), buttock fat thickness (BFT, cm), average backfat thickness (ABT, cm), leaf fat weight (kg), caul fat weight (kg), loin eye height (cm), and loin eye area (cm2).
Isolation and sequencing of pig AMPD1 promoter regions
Pig AMPD1 promoter regions were obtained from genomic DNA by PCR of genomic DNA. Two PCR primers (AMPD1-PF and AMPD1-PR; Table 1) were designed from the published nucleotide sequence of the pig AMPD1 promoter (GenBank accession no. HM487108) (Wang et al., 2008). PCR amplifications were carried out from four pig breeds (three animals per breed), including Large White, Landrace, Meishan, and Tongcheng. The purified products were subcloned into the pMD-19T vector (TaKaRa, Dalian, China) and sequenced by Invitrogen Life Technology Co., Ltd. (Shanghai, China). The nucleotide sequences of each animal were compared to determine polymorphisms in the pig AMPD1 promoter. Analyses were performed using the Lasergene software program Seqman (DNAstar, Inc., Madison, WI).
AMPD1, AMP deaminase 1.
PCR-RFLP genotyping
To genotype the polymorphic site in the pig AMPD1 gene promoter, PCR-RFLP was performed. Two primers (SNP-F and SNP-R; Table 1) were used to amplify the fragment from genomic DNA. It yielded a 401-bp PCR product including a polymorphic locus for g.-626 G > A and g.-566 A>G. PCR was carried out in a 25 μL volume containing 2.5 μL 10 × PCR buffer (with NH4 +), 1.5 μL 25 mM MgCl2, 1.5 μL 2 mM of each dNTP, 0.5 μL of each primer (10 μM), 0.5 U Taq DNA polymerase (Biostar International, Toronto, Canada), and 50 ng DNA. The PCR cycling program remained at 94°C for 4 min, followed by 35 cycles at 94°C for 30 s, staying at 60°C for 30 s and 72°C for 40 s, and a final extension at 72°C for 8 min. PCR products (401 bp) were digested (10 μL reaction solution containing 3 U restriction enzyme) with BglII and MaeI restriction enzymes (TaKaRa) followed by electrophoresis on 2% agarose gels stained with ethidium bromide.
Quantitative real-time analysis
For AMPD1 gene expression analysis between different genotypes, longissimus dorsi and biceps femoris muscle tissues were sampled from the F2 of Large White and Meishan pigs. Total RNA was isolated using the TRIzol reagent (Invitrogen, CA). The quantitative PCR (qPCR) was used to detect the expressions level of AMPD1 in muscle tissues. Each qPCR (total volume 10 μL) included 5 μL of SYBR Green Real-Time PCR Master Mix (TaKaRa), 0.8 μL of cDNA template, and 0.4 μL of primer pairs (RT-PCRF and RT-PCRR; Table 1). The cycling conditions consisted of an initial, single cycle for 30 s at 95°C followed by 40 cycles consisting of 5 s at 95°C, 30 s at 58°C, and 10 s at 72°C, followed by a final extension for 7 min. The specificity of the PCR products was confirmed by melting curve analysis. An optimized comparative Ct (2−▵▵Ct) value method was used to quantify the gene expression levels relative to the expression of beta-actin. All PCR amplifications were performed in triplicate for each cDNA sample. One-way analysis of variance was employed using SPSS version 13.0 to compare the differences in gene expression in the muscles of different haplotypes and Duncan's new multiple range test was used to analyze their statistical significance.
Construction, transfection, and activity assay of luciferase plasmids
To evaluate the effects of the two SNPs (g.-626 G > A and g.-566 A>G) on AMPD1 promoter activity, the promoter region of the pig AMPD1 gene was amplified using PCR with primers (PGL3-F and PGL3-R; Table 1) with individuals of the haplotype H1 (A−626G−566) and the haplotype H2 (G−626A−566) as a template, respectively. The product was ligated into the pGEM-T Easy vector (Promega, Madison, WI) and then subcloned into the KpnI/SmaI site of the pGL3-Basic vector (Promega). Then, two reporter plasmids (haplotype H1 and haplotype H2) were constructed, and further identified by double digestion and sequencing.
C2C12 cells were seeded into 24-well plates (0.6 × 105 cells/well) 24 h before transfection. Transfections were then performed using 800 ng of luciferase reporter construct DNA and 80 ng of an internal control plasmid pRT-TK (Promega). After 6 h, the transfection medium was removed and the cells were incubated in differentiation medium (DMEM with 2% horse serum) for 48 h. Firefly and Renilla luciferase activities (Dual Luciferase Reporter Assay System; Promega) were measured according to the manufacturer's instructions. The firefly luciferase activity was normalized to that of Renilla luciferase.
Statistical analysis
HAPLOVIEW 4.2 software was used to analyze the pairwise linkage disequilibrium (LD; r2) between two SNPs in the pig AMPD1 promoter region (Barrett et al., 2005). Haplotypes of F2 individuals were obtained by using the PHASE v2.1 program (Stephens et al., 2001). The association between genotypes and traits recorded was performed with the general linear model procedure (SAS Institute, Inc. Cary, NC). The statistical model was assumed to be: Tijkl = μ + Si + Yj + Gk + F l + bijklXijkl + eijkl , where Tijkl is the observed values of traits, μ is the overall mean, Si is the effect of sex (i = 1 for male or 2 for female), Yj is the effect of year (j = 1 for 2000 or 2 for 2003), Gk is the effect of haplotype combinations (K = H1H1, H1H2 and H2H2), Fl is the random effect of family (l = 1 to 37), bijkl is the random regression coefficient of the slaughter weight in variant families, Xijkl is the slaughter weight, and eijkl is the random residual.
Results
Genetic variation in the pig AMPD1 gene promoter regions
To find the SNPs in the AMPD1 gene promoter regions, one primer pair (AMPD1-PF and AMPD1-PR;Table 1) was used to amplify the AMPD1 gene proximal promoter sequences from 12 individuals of four breeds (two Western meat-type breed Landrace and Large White and two Chinese indigenous breed, Meishan and Tongcheng). As given in Figure 1A, two nucleotide substitutions g.-626 G > A and g.-566 A>G, which are related to the position of the transcription start site (−626 and −566), were detected in the AMPD1 promoter sequences (GenBank accession no. HM487108).
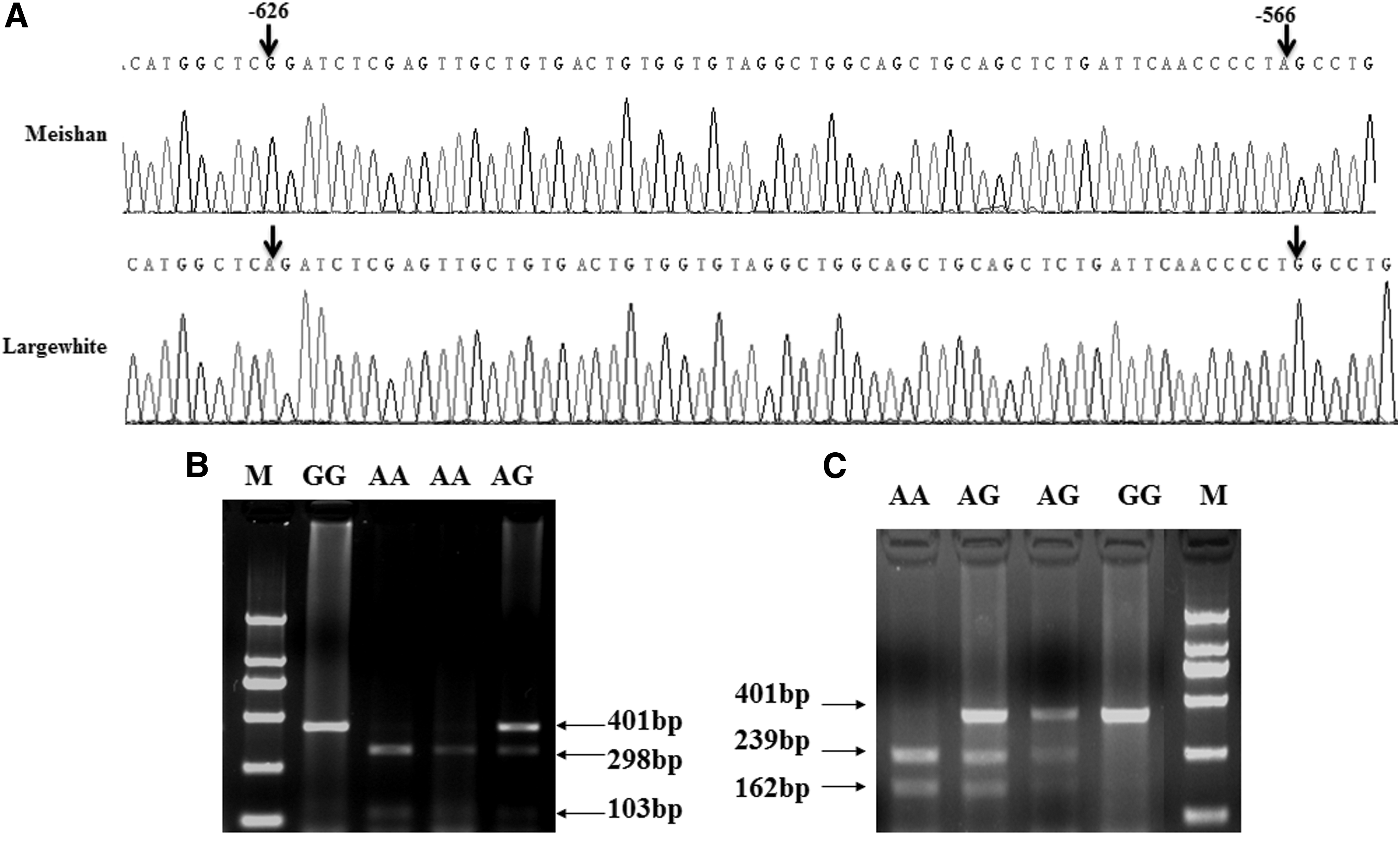
Two SNPs identified in the promoter region of the pig AMPD1 gene.
To genotype the SNPs using the PCR-RFLP assay, a new primer pair (SNP-F and SNP-R; Table 1) was used to amplify a 401-bp fragment containing two SNPs and then digested with a differential endonuclease. As shown in Figure 1B, the g.-626 G > A mutation could be detected by digestion with BglII, resulting in allele G (401 bp) and allele A (298 and 103 bp). The SNP g.-566 A>G mutation was genotyped using the restriction enzyme MaeI, with allele G (401 bp) and allele A (239 and 162 bp) (Fig. 1C).
Allele frequencies of the pig AMPD1 gene in different breeds
We analyzed the genotypes and allele frequencies of two SNPs in 159 unrelated pigs representing four Chinese indigenous and Western meat-type breeds. As shown in Table 2, at the g.-626 G > A loci, Chinese indigenous breeds (Meishan and Tongcheng) had higher frequencies of the G allele, with values of 90.2% and 90.4%, respectively. In contrast, at the g.-566 A>G loci, allele A was the dominant allele in Chinese pigs (Meishan and Tongcheng) with frequencies of 94.1% and 96.2%, respectively.
Association of haplotypes of pig AMPD1 gene with carcass traits
An LD test revealed D’ values of 0.94 between g.-626 G > A and g.-566 A>G, indicating that the two loci are tightly linked. Two major haplotypes of H1 (A−626G−566, 47.1%) and H2 (G−626A−566, 44.8%) were detected in the Large White × Meishan pig population. Because other haplotypes were very rare in the F2 population, associations with carcass traits were analyzed only with the haplotype combinations H1H1, H1H2, and H2H2. The results indicated that the pigs with the haplotype combination H1H1 had significantly higher LMP values (p < 0.01), but lower ABT (cm), BFT (cm), and TFT (cm) values than the pigs with the haplotype combination of H1H2 and H2H2 (p < 0.05) (Table 3).
Same superscript letters in one column indicate no significant differences between the different genotypes. An asterisk (*) indicates significant difference at p < 0.05 level; a double asterisk (**) indicates significant difference at p < 0.01 level. H1 haplotype of the g.-626 A and g.-566 G, H2 haplotype of the g.-626 G and g.-566 A.
ABT, average backfat thickness; BFT, buttock fat thickness; LMP, lean meat percentage; SFT, shoulder fat thickness; TFT, thorax-waist fat thickness.
The two SNPs in the promoter of the AMPD1 gene significantly affected its transcriptional activity
To further investigate whether the two haplotypes H1 (A−626G−566) and H2 (G−626A−566) could affect the promoter transcriptional activities of the pig AMPD1 gene, we designed two reporter plasmids carrying the two haplotypes, constructed using PCR with primers (Table 1), for the individuals of the A−626G−566 and G−626A−566 haplotype as a template. As given in Figure 2A, the AMPD1 promoter fragment containing haplotype H1 (A−626G−566) showed 2.3-fold greater activity in the C2C12 cells than the AMPD1 promoter construct for haplotype H2 (G−626A−566).

Two SNPs in the promoter region affected AMPD1 gene transcriptional activity.
To further determine whether the two haplotypes could affect the pig AMPD1 gene expression, we selected F2 Large White and Meishan pigs with the haplotype combinations H1H1, H1H2, and H2H2 to compare the expression of the AMPD1 gene in muscle tissues. Pigs carrying the haplotype combination H1H1 had significantly (p < 0.01 or p < 0.05) higher expression levels of the AMPD1 gene than pigs with the haplotype combinations H1H2 and H2H2 in the longissimus dorsi (Fig. 2B) and biceps femoris (Fig. 2C) muscle tissues. No significant differences in expression were observed between the haplotype combinations H1H2 and H2H2.
Discussion
AMPD1 is a rate-limiting enzyme of the purine nucleotide cycle, and a deficiency of this enzyme can cause muscle dysfunction and cardiovascular diseases (Palmer et al., 2007; Feng et al., 2017; Rannou et al., 2017). In mice, the disruption of AMPD1 in skeletal muscle improves glucose tolerance and enhances insulin clearance in animals fed HFD (Cheng et al., 2014). An 18-bp deletion mutation in the AMPD1 gene was identified in cattle, which is significantly associated with many carcass traits in the Qinchuan cattle population (He et al., 2010). In chicken, three SNPs have been detected in exons 4, 6, and 8 of the AMPD1 gene in Fast Partridge and Lingshan chickens; these SNPs exhibited significant associations with inosine monophosphate (IMP) content (Hu et al., 2015). To date, there have been no studies of potentially functional genetic variants in the promoter region of the AMPD1 gene. In this study, two novel SNPs (g.-626 G > A and g.-566 A>G) in the proximal promoter region of the AMPD1 gene were identified.
The allele frequencies of g.-626 G > A and g.-566 A>G exhibited significant differences between Western meat-type breeds and Chinese indigenous breeds. Western breeds had higher frequencies of the A allele at the g.-626 G > A loci and the G allele at the g.-566 A>G loci. LD analysis indicated that the g.-626 G > A and g.-566 A>G were detected with high LD values (r 2 = 0.94). The results suggested that the two SNPs may have consistent effects on carcass traits. Further association analysis showed that the haplotype combination H1H1 was significantly associated with LMP and ABT, showing that these two loci were linked to each other in the F2 Large White × Meishan pig resource family. Pigs with the haplotype combination H1H1 had a higher LMP and lower ABT than other pigs. This result suggests that the H1H1 haplotype may be used as a molecular marker for the selection of pigs with higher LMP and lower ABT values. It could be speculated that improving the frequencies of allele G at the g.-566 A>G locus and allele A at the g.-626 G > A locus could help improve the genetic progress of carcass traits in the pig population. However, the number of pigs analyzed is limited. Further investigation of other pig populations is required to confirm the association between the H1H1 haplotype and carcass traits.
To investigate whether the g.-626 G > A and g.-566 A>G mutations in the promoter region alter transcriptional activity, we constructed recombinant constructs containing haplotypes H1 (A−626G−566) and H2 (G−626A−566). The results from transient transfection experiments showed that the transcriptional activity of haplotype H1 (A−626G−566) was significantly higher than that of H2 (G−626A−566) (p < 0.05). We also confirmed that pigs with H1 (A−626G−566) had significantly higher expression levels of the AMPD1 gene than pigs with H2 (G−626A−566) in muscle tissues. In this study, the g.-626 G > A and g.-566 A>G loci are located at the core promoter region of the AMPD1 gene, suggesting that the two SNPs are likely to regulate the pig AMPD1 gene expression.
To determine whether SNPs in the promoter region could affect the AMPD1 expression through the disruption of transcription factor binding sites, we searched for transcription factors using genomatrix (
Conclusions
In summary, two novel SNPs (g.-566 A>G and g.-626 G>A) in the proximal promoter of the AMPD1 gene were found to be associated with LMP and ABT and could be used as a molecular marker for carcass traits in pig breeding programs. The two SNPs significantly affected the transcriptional activity of the AMPD1 gene promoter. Further work is necessary to investigate the molecular mechanisms associating carcass traits with the two SNPs in the promoter of the pig AMPD1 gene.
Footnotes
Acknowledgments
This work was supported by the Scientific Research Starting Foundation for Returned Overseas Chinese Scholars of Sichuan Province and Hainan Provincial Natural Science Foundation of China (2017CXTD013).
Disclosure Statement
No competing financial interests exist.