
Abstract
The role of chemotherapeutic agents in tumor immunotherapy is still controversial. In this study, we test using a TC-1 tumor model whether gemcitabine plus E7 peptide vaccine regimens (E7 peptides+CpG-ODN+anti-4-1BB Abs) may result in tumor cure in mice with large established tumors, with a focus on their effects on Ag-specific cytotoxic T lymphocyte (CTL) and myeloid-derived suppressor cell levels. Gemcitabine inhibited tumor growth by its direct cytotoxicity to tumor cells in vivo. E7 peptide vaccine regimens enhanced Ag-specific CTL lytic and antitumor therapeutic activity. Initial combination therapy using gemcitabine and E7 peptide vaccine regimens resulted in tumor regression with tumor relapse in animals with large established tumors, which appeared to result from the suppression of Ag-specific CTL activity by gemcitabine treatment. However, optimization of gemcitabine therapy by reducing its dose and frequency led to complete tumor regression without any recurring tumors in all tested mice even after discontinuation of therapy, possibly due to Ag-specific CTL responses. Thus, this study shows that the optimal dose and therapy frequency of gemcitabine are critical for achieving tumor cure in tumor-bearing animals undergoing E7 peptide vaccine regimen therapy, mainly by preventing CTL suppression. These findings may have implications for designing peptide-based therapeutic vaccines in cancer patients undergoing chemotherapy.
Introduction
H
Gemcitabine is an FDA-approved cytotoxic nucleoside analog that has been used to treat patients with certain types of cancer. In addition to its cytotoxic effects, gemcitabine may increase antigen uptake from apoptotic tumor cells by local DCs for tumor antigen presentation to CD8+ T cells (Nowak et al., 2003). Gemcitabine treatment reduced the number of myeloid-derived suppressor cells (MDSCs) in the tumor microenvironment, possibly potentiating tumor-reactive CTL responses (Suzuki et al., 2005; Mundy-Bosse et al., 2011). Moreover, certain cytotoxic chemotherapeutic drugs induce endogenous immune responses by triggering apoptotic tumor cell death and subsequently eliciting tumor antigen-specific CTLs (Martins et al., 2009; Zitvogel et al., 2010). On the other hand, gemcitabine had an inhibitory effect on adaptive immune responses in a murine tumor model (Bauer et al., 2014). In that study, systemic delivery of gemcitabine plus DC vaccination increased antitumor therapeutic activity through increased sensitivity of drug-treated tumor cells to CTL-mediated killing. These reports have suggested a potential role of gemcitabine in modulating the tumor microenvironment by removing MDSCs. Recently, we reported that systemic delivery of gemcitabine had dramatic effects on tumor growth inhibition, but not MDSC inhibition in a CT26/HER2 tumor model (Danishmalik et al., 2017b). This prompted us to test using a TC-1 model whether systemic delivery of gemcitabine along with E7 peptide vaccination in the presence of CpG-ODN (for stimulation of antigen-presenting cells [APCs]) and anti-4-1BB (for activation of CD8+ T cells) might lead to tumor cure in animals with large established tumors, with a special focus on their effects on Ag-specific CTL and MDSC levels.
In this study, we observed that gemcitabine did not elicit endogenous CTL responses nor depleted MDSCs, while inhibiting Ag-specific CTL activity induced by E7 peptide vaccine regimens. Optimization of gemcitabine therapy to avoid Ag-specific CTL inhibition led to tumor cure in animals receiving E7 peptide vaccine regimen therapy. In particular, both increased sensitivity of gemcitabine-exposed tumor cells to CTL-mediated killing, and long-term memory antitumor immunity appeared to be associated with tumor cure in this model.
Materials and Methods
Mice and tumors
Female 6-week-old C57BL/6 mice were purchased from Daehan Biolink Co. (Chungbuk, Korea). TC-1 cells are epithelial cells of C57BL/6 origin and express HPV 16 E6 and E7, which have been used as an animal model of human cervical cancer. They were grown in cRPMI media (10% FBS, 1% L-glutamine, and 1% penicillin/streptomycin) supplemented with 400 μg/mL of G418. A murine colon adenocarcinoma cell line, MC38 cell line, was provided from J. Schlom (NCI, Bethesda) and grown in cDMEM media. In this study, the mouse strain (C57BL/6, H-2b) was tested as its major histocompatibility molecules match with those of TC-1 and MC38 cells. The mice were cared for under the guidelines of the Institutional Animal Care and Use Committee-approved protocols (KW-130419-1). The tumor cells were washed twice with phosphate-buffered saline (PBS) and injected into the mice.
Reagents and treatment of mice
For peptide injection, animals were immunized subcutaneously (s.c.) at 0, 1, and 2 weeks. The parameters of these immunizations were as follows: 20 μg of E7 CTL peptide (RAHYNIVTF) per mouse; with or without 20 μg of CpG-ODN; final volume of 100 μL in PBS; and 31-gauge needle (BD Biosciences, San Jose, CA). HPV 16 E7 peptides were purchased from Peptron (Taejon, Korea). The CpG-ODN (unmethylated form) designated as 1826 (5′-TCCATGACGTTCCTGACGTT-3′) was purchased from GenoTech (Taejon, Korea). It was synthesized with a nuclease-resistant phosphorothioate backbone, dissolved in water, and then confirmed to have an undetectable endotoxin level. For delivery of anti-4-1BB Abs, animals were injected intraperitoneally (i.p.) with 100 μg of anti-4-1BB Abs, generated from hybridoma cells (3H3), which were a kind gift of R. Mittler (Emory University, Atlanta, GA). Specifically, pristane-primed nude mice were injected with 3H3 cells for ascites fluid production, from which anti-4.1BB Abs were purified using a protein G column (Sigma-Aldrich, St. Louis, MO). Control rat IgG was purified from rat serum. Animals were also treated i.p. with gemcitabine (Gemzar®, Lilly) twice weekly (at 3-day intervals) at a dose of 75 μg/g body weight. This dose has been previously tested as a dose similar to the equivalent dose used in patients (Suzuki et al., 2005; Mundy-Bosse et al., 2011; Bauer et al., 2014, 2015). For combination therapy, the two drug doses, 75 and 37.5 μg/g body weight, were tested.
In vivo depletion of CD8+ T and natural killer cells
Anti-CD8 IgGs (100 μg) or anti-asialo GM1 Abs (200 μL) were administered i.p. on the indicated days. A hybridoma cell line (clones 2.43) was purchased from the American Type Culture Collection (Manassas, VA), and anti-CD8 IgG was obtained as previously described (Sin et al., 2013). Control rat IgG was purchased from Sigma-Aldrich. Anti-CD8 IgG treatment resulted in more than 98% depletion of CD8+ T cells at 7 days following Ab injection. Anti-asialo GM1 rabbit Abs were purchased from Wako Pure Chemical (Osaka, Japan). Anti-asialo GM1 rabbit Abs were diluted in accordance with the manufacturer's protocol and injected in 200 μL of PBS (containing the diluted anti-asialo GM1 Abs) per mouse. This Ab treatment resulted in 80–90% depletion of natural killer (NK) cells (expressing a CD3−/CD49b+ phenotype) at 5 days following Ab injection.
In vivo CTL lytic activity assay
Spleen cells from naive mice were treated with red blood cell lysis buffer (Sigma-Aldrich). One fraction of the splenocytes was then pulsed with 5 μg of E7 peptides in cRPMI for 60 min at 37°C, while the other fraction was left unpulsed. To generate peptide-pulsed cells with high carboxyfluorescein diacetate succinimidyl ester (CFSE), the peptide-pulsed splenocytes were incubated with 20 μM CFSE in RPMI (2.5% FBS) for 15 min. The unpulsed cells were instead incubated with 2.5 μM CFSE in RPMI (2.5% FBS) for 15 min to generate nonpeptide-pulsed cells with low CFSE. The cells were then washed thrice with PBS to remove unbound CFSE. Finally, an equal number of pulsed and unpulsed cells (a total of 2 × 107 cells/0.4 mL/mouse) was injected intravenously into the tested mice. After 18 h, the mice were sacrificed and the spleens were collected. After lysing the red blood cells, the splenocytes were analyzed directly for the two cell populations with CFSE staining (CFSE low vs. CFSE high) using a flow cytometer (BD Biosciences). The percentage of lysed cells (% lysis) was calculated as 100 × [1−(r unprimed/r primed)]. The ratio (r) was calculated as %CFSElow/%CFSEhigh.
Flow cytometry
To test the levels of CD11b+Gr-1+ MDSCs in the tumor tissues, CD45+ cells were gated from the cell suspensions from the tumor tissues. In detail, mice were sacrificed to obtain tumors. The tumors were put into 3 mL of RMPI and cut into small pieces, which were subsequently broken using needle tips and then treated with collagenase (1.5 mg/mL) and DNase I (0.5 μg/mL) at 37°C for 3 h. The tumor cell suspensions were applied to a 70 mm cell strainer. The collected cells were pretreated for 10 min with anti-CD16/32 (Fc blocker) and then stained with allophycocyanin-labeled CD45, phycoerythrin-labeled anti-CD11b, and fluorescein isothiocyanate-labeled anti-Gr-1. The tested Abs were purchased from BioLegend (San Diego, CA). In each step, the cells were washed thrice with the washing buffer (PBS +1% FBS). Finally, the cells were analyzed using a flow cytometer (BD Biosciences).
Tumor cell challenge
For the antitumor therapeutic studies, 2 × 105 TC-1 cells were injected s.c. into the right flank of C57BL/6 mice. When the mean tumor size was ∼2 or 5 mm, the animals were injected s.c. with 20 μg of E7 peptides and 20 μg of CpG-ODN per mouse in a final volume of 100 μL PBS. The animals were also injected i.p. with 100 μg of anti-4-1BB Abs. In the case of tumor cell rechallenge studies, 5 × 105 TC-1 and MC38 cells were injected s.c. into each flank of C57BL/6 mice. The mice were monitored twice per week for tumor growth. The tumor size was measured in mm using a caliper, and was recorded as the mean diameter [longest surface length (a) and width (b), (a + b)/2]. Animals with complete tumor regression were denoted as those showing no detectable tumors by hand manipulation techniques. The mice were euthanized when the mean diameter of the tumor exceeded 20 mm.
Statistical analysis
Statistical analysis was performed by the independent t-test or one-way ANOVA using the SPSS 17.0 software program. The values of the experimental groups were compared with the values of the control group. Any p-values <0.05 were considered to be significant.
Results
CpG-ODN and anti-4-1BB, as an E7 peptide adjuvant, are effective at enhancing Ag-specific CTL lytic and antitumor therapeutic activity
First, we examined whether immunization with E7 peptides plus CpG-ODN enhances Ag-specific CTL and antitumor therapeutic activity significantly compared to the therapeutic activity of the E7 peptide alone in vivo. As shown in Figure 1A, B, animals receiving immunization with E7 peptides alone induced Ag-specific CTL lytic activity significantly greater than that in the negative controls. Notably, effects of the H-2b-restricted immuno-dominant E7 CTL peptides have been previously reported (Feltkamp et al., 1993). Furthermore, animals receiving immunization with E7 peptides plus CpG-ODN induced Ag-specific CTL lytic activity significantly greater than that in animals receiving the E7 peptides alone. However, no such effects were observed when animals were vaccinated with CpG-ODN alone. Thus, these data suggest that TLR9 stimulation by CpG-ODN enhances Ag-specific CTL lytic activity induced by E7 peptides in the TC-1 model. Figure 1C shows the antitumor therapeutic activity of treatment using E7 peptides and CpG-ODN. In this study, TC-1 tumor (5 mm in mean tumor size)-bearing animals showed a significant level of tumor growth inhibition over the time points measured following treatment with E7 peptides+CpG-ODN, compared to that in animals treated with either E7 peptides or CpG-ODN alone. However, despite this antitumor effect, none of the mice displayed complete tumor regression. Moreover, it should be noted that although E7 peptides alone induced a degree of Ag-specific CTL lytic activity, tumor-bearing mice receiving E7 peptides failed to display antitumor therapeutic activity, suggesting that tumor control appears to require a level of CTL activity greater than the level induced by E7 peptides alone. We next tested the possibility that additional stimulation of 4-1BB with anti-4-1BB Abs may augment Ag-specific CD8+ CTL responses induced by E7 peptides plus CpG-ODN, thereby leading to increased antitumor therapeutic activity against large established tumors; 4-1BB signaling has been known to activate CD8+ T cells and expand their life span (Cooper et al., 2002; Lee et al., 2002). For this testing, we immunized mice s.c. with E7 peptides+CpG-ODN, and i.p. with anti-4-1BB Abs simultaneously. As shown in Figure 1D, E, animals immunized with E7 peptides+CpG-ODN+anti-4-1BB Abs displayed Ag-specific CTL lytic activity significantly greater than that in animals immunized with E7 peptides+CpG-ODN. Similar to previous results, we also observed that animals immunized with E7 peptides+CpG-ODN induced Ag-specific CTL lytic activity significantly greater than that in untreated control mice. Furthermore, animals injected with anti-4-1BB Abs alone showed no Ag-specific CTL lytic activity in a manner similar to untreated control mice. We next treated the large tumor (5 mm)-bearing animals s.c. with E7 peptides+CpG-ODN, and i.p. with anti-4-1BB Abs simultaneously (Fig. 1F). Tumor-bearing mice treated with E7 peptides+CpG-ODN+anti-4-1BB Abs displayed tumor growth inhibition significantly more than that in animals treated with E7 peptides+CpG-ODN. However, animals treated with anti-4-1BB Abs failed to show therapeutic antitumor activity as did untreated control mice. In this study, it is notable that the tumor-bearing animals treated with E7 peptides+CpG-ODN+anti-4-1BB Abs were unable to eradicate tumors. Thus, these data collectively suggest that a combination of E7 peptides+CpG-ODN+anti-4-1BB Abs (henceforth called E7 peptide vaccine regimens) is an effective regimen to augment Ag-specific CTL lytic activity for tumor control, and that stimulation with CpG-ODN and anti-4-1BB Abs contributes to the augmentation of both Ag-specific CTL and antitumor therapeutic activity in the E7 peptide vaccine model.

Ag-specific CTL lytic and antitumor therapeutic activity in animals injected with E7 peptides plus CpG-ODN
Gemcitabine directly inhibited tumor growth in mice without any involvement of immune effector cells, NK and CD8+ T cells
It was reported that gemcitabine reduced the number of MDSCs, leading to increased activity of tumor-reactive CTL and NK cells (Suzuki et al., 2005). Mundy-Bosse et al. (2011) also reported that gemcitabine depleted MDSCs in a C26 tumor model. Moreover, gemcitabine may induce endogenous immune responses, thus eliciting Ag-specific CTLs. To test this possibility, we depleted the tumor-bearing animals receiving gemcitabine therapy of CD8+ T cells and NK cells, and then measured tumor growth patterns. Initially, we evaluated whether tumor (tumor sizes, 2 vs. 5 mm)-bearing animals receiving gemcitabine therapy may experience complete tumor regression. When tumor (2 mm)-bearing mice were treated with gemcitabine, they showed a degree of tumor regression (Fig. 2A). When tumor (5 mm)-bearing mice were treated with gemcitabine, they showed tumor growth inhibition without any tumor regression over the time points (Fig. 2B). This result demonstrates that small tumors, but not large tumors, may be removed by the systemic delivery of gemcitabine and that there may be a limit to the tumor sizes that can be removed by chemotherapy. We next examined whether gemcitabine treatment influenced tumor control through modulation of immune effector functions. As shown in Figure 2C, there was no significant difference in antitumor therapeutic effectiveness between gemcitabine-treated animals with and without NK and CD8+ T cells, as determined by immune cell subset depletion. Therefore, these data highlight the notion that gemcitabine inhibits tumor growth primarily by its direct cytotoxicity to tumor cells.
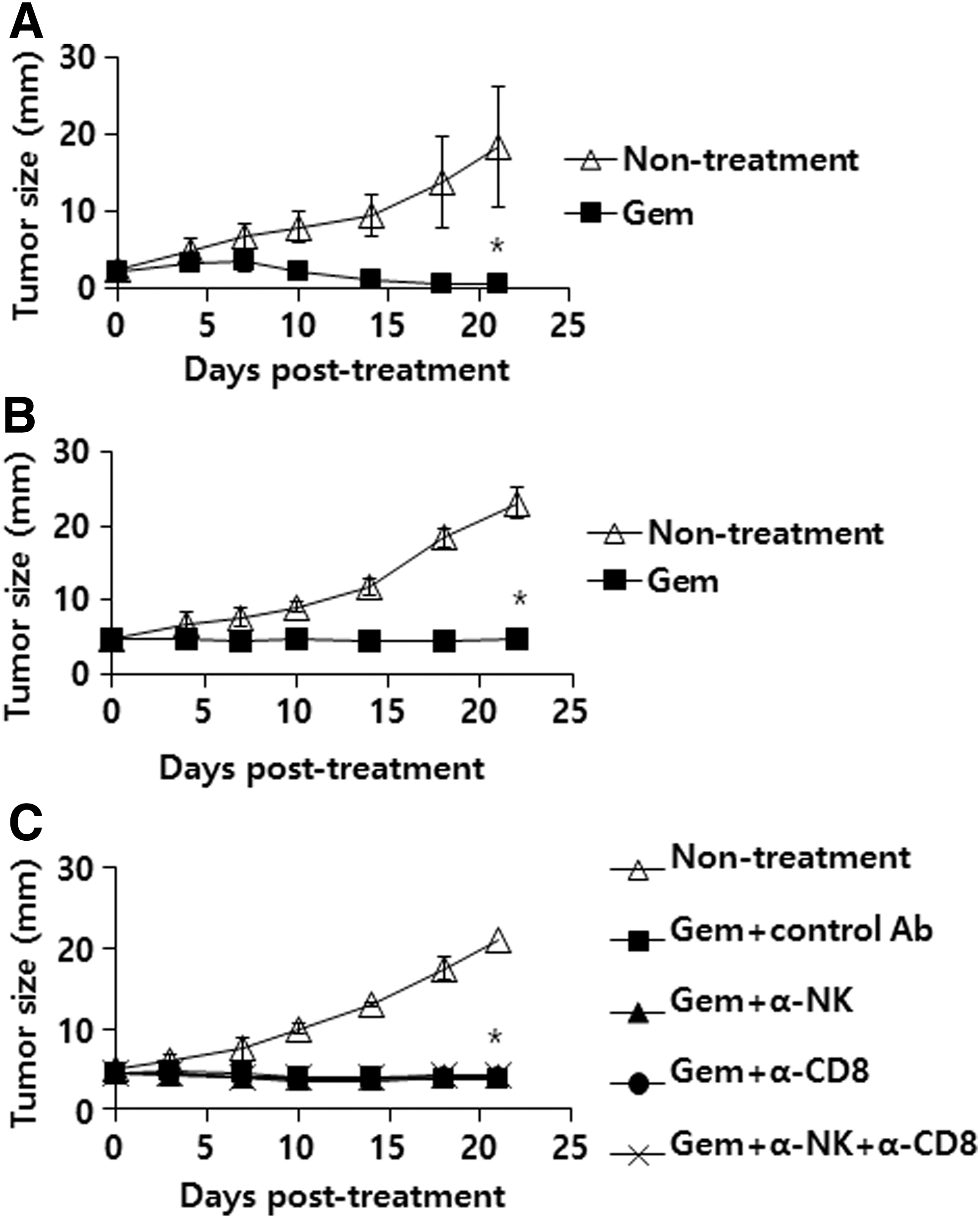
Therapeutic antitumor effects of gemcitabine in animals with tumor sizes (mean tumor diameters; 2 mm vs. 5 mm)
Combined therapy using gemcitabine plus E7 peptide vaccine regimens resulted in complete tumor regression with recurring tumors in animals with large established tumors
We hypothesized that combination therapy using gemcitabine and E7 peptide vaccine regimens might possibly induce complete tumor regression in mice with large established tumors (tumor sizes, 5 mm). To test this hypothesis, we first treated tumor-bearing mice with gemcitabine (75 μg/g body weight) and E7 peptide vaccine regimens, and measured tumor growth patterns over indicated time points. However, there was an insignificant degree of antitumor combination effect when the two therapy protocols were combined (data not included). Moreover, none of Ag-specific CTL lytic activity was detected in the tumor-bearing mice under treatment with gemcitabine (75 μg/g body weight) plus E7 peptide vaccine regimens (data not included). Thus, we reduced the drug dose from 75 μg/g body weight to 37.5 μg/g body weight. This treatment protocol is illustrated in Figure 3A. As shown in Figure 3B, animals receiving treatment with gemcitabine plus E7 peptide vaccine regimens experienced tumor regression dramatically greater than that observed in animals receiving single agent therapy. Overall, 5 of 10 animals displayed complete tumor regression at 30 days following the first treatment, whereas the remaining 5 animals failed to show complete removal of tumors. In addition, animals receiving treatment with either gemcitabine or E7 peptide vaccine regimens alone failed to show complete tumor removal. However, they exhibited a more significant degree of tumor growth control compared to that in the nontreatment group. Complete tumor regression and tumor recurrence rates are shown in Figure 3C. Among 10 tumor-bearing animals receiving combination therapy, 5 experienced complete tumor regression (50% tumor regression rates) at 30 days of the first treatment. However, tumors recurred at 60 days following the first treatment (100% tumor recurrence rates) in the five mice displaying complete tumor regression. Overall, all the tested animals failed to eradicate tumors at 60 days posttreatment by this combination therapy schedule. Thus, these results demonstrate that this combination therapy scheme synergistically inhibits tumor growth, leading to complete tumor regression in some animals, in which tumors recurred after discontinuation of therapy.
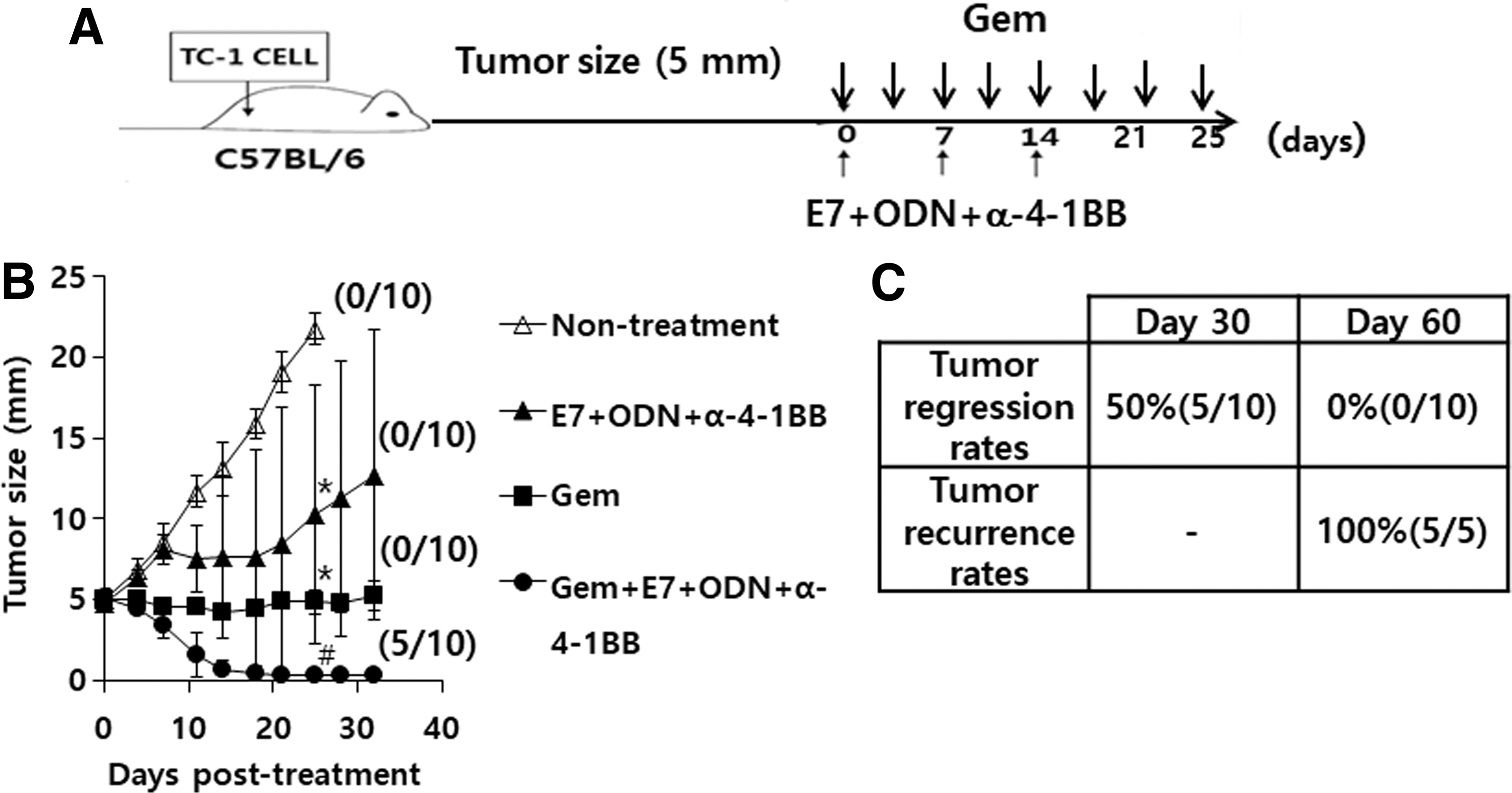
Tumor control and recurrence rates of tumor-bearing mice by combined therapy with gemcitabine plus E7 peptide vaccine regimens (E7 peptides+ODN+anti-4-1BB Ab).
Tumor-bearing mice receiving treatment with gemcitabine plus E7 peptide vaccines regimens showed Ag-specific CTL lytic activity dramatically lower than that in animals receiving E7 peptide vaccine regimens alone without any impact on MDSCs in the tumor tissues
We investigated why the combination therapy scheme failed to maintain the ability to control tumors over time in tumor-regressed animals. We speculated that gemcitabine might suppress Ag-specific CTL responses induced by E7 peptide vaccine regimens during combination therapy. To test this hypothesis, we measured and compared the levels of Ag-specific CTL lytic activity between tumor-bearing mice treated with E7 peptide vaccine regimens and those treated with gemcitabine (37.5 μg/g body weight) plus E7 peptide vaccine regimens. As shown in Figure 4A, the tumor-bearing mice receiving E7 peptide vaccine regimen therapy displayed a significant degree of Ag-specific CTL lytic activity. In contrast, the tumor-bearing mice receiving treatment with gemcitabine plus E7 peptide vaccines regimens showed Ag-specific CTL lytic activity dramatically lower than that in animals receiving E7 peptide vaccine regimens alone. However, untreated animals and animals receiving gemcitabine treatment alone showed a background level of Ag-specific CTL lytic activity. This result underscores the notion that gemcitabine alone does not induce endogenous CTL responses; however, it may inhibit E7 vaccine-induced CTL responses. We next evaluated the levels of MDSCs in the tumor tissues. As shown in Figure 4B, the percentages of MDSCs (expressing a CD11b+/Gr-1+ phenotype) among the CD45+ cell populations in the animals receiving gemcitabine therapy were similar to those in the untreated animals. A similar finding was observed between the animals receiving therapy with gemcitabine plus E7 peptide vaccine regimens and the animals receiving therapy with E7 peptide vaccine regimens. These data suggest that gemcitabine may not have any inhibitory effect on MDSCs in the tumor tissues. Taken together, these results demonstrate that gemcitabine treatment may lead to the inhibition of Ag-specific CD8+ CTL responses resulting from E7 peptide vaccine regimen therapy, while sustaining their minimal ability to kill tumor cells. Moreover, gemcitabine treatment is unlikely to result in reducing MDSCs in the tumor microenvironment.
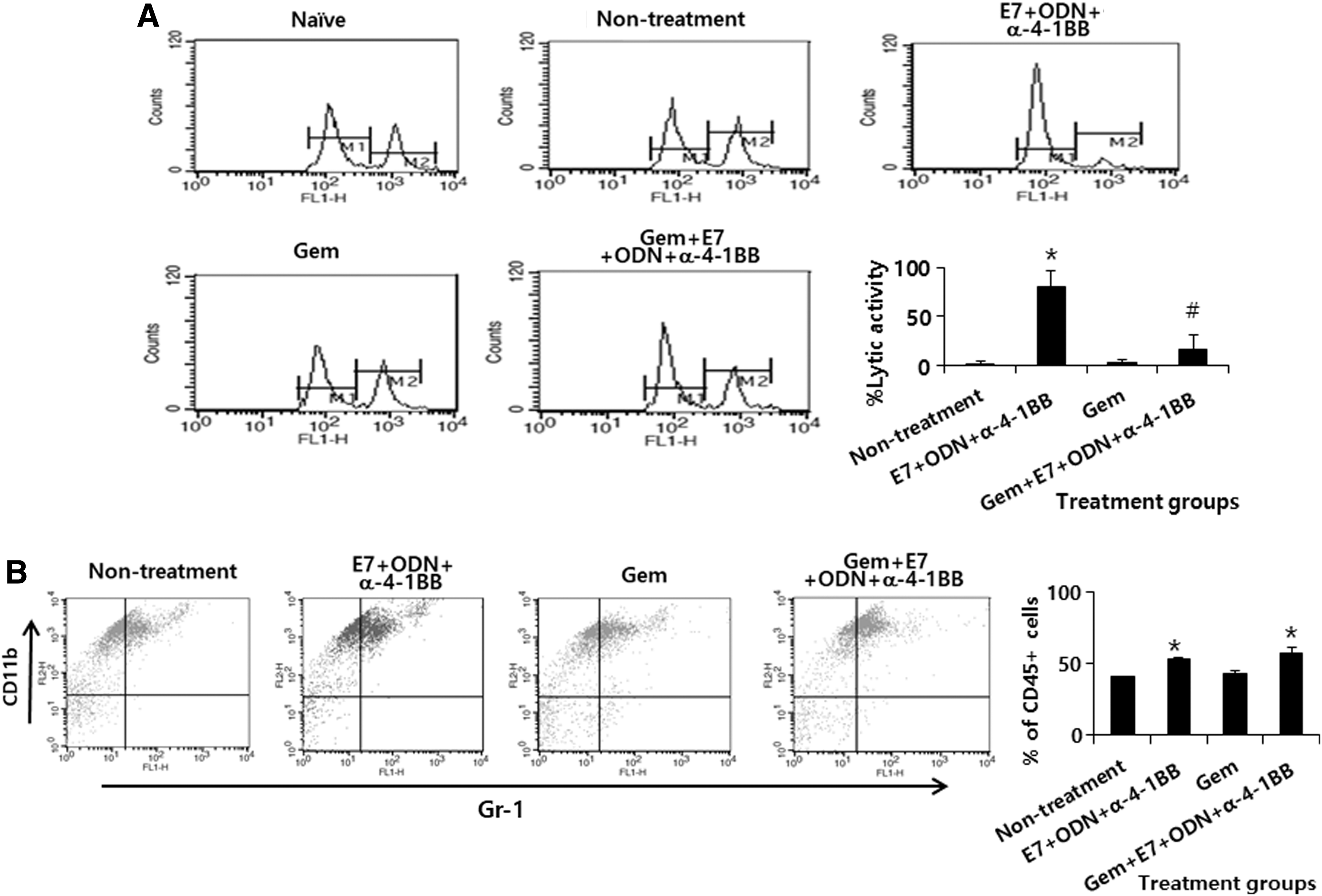
Ag-specific CTL lytic activity and MDSC populations in tumor-bearing animals treated with gemcitabine in combination with E7 peptide vaccine regimens (E7 peptides+CpG-ODN+anti-4-1BB Abs).
Optimized gemcitabine therapy in combination with E7 peptide vaccine regimens was critical for inducing complete tumor regression without any recurring tumors in mice with large established tumors
Next, we hypothesized that in tumor-regressed animals receiving combination therapy, a dramatic decrease in Ag-specific CTL lytic activity might be associated with tumor recurrence and that a reduction of gemcitabine therapy frequency might lead to complete tumor regression without tumor relapse. To test this hypothesis, gemcitabine therapy was discontinued when tumor sizes became less than 2 mm in mean diameter (Fig. 5A). As shown in Figure 5B, animals receiving treatment with gemcitabine plus E7 peptide vaccine regimens experienced tumor regression dramatically greater than that observed with single agent therapy, similar to previous observations (Fig. 3B). In addition, animals receiving single agent treatment failed to control tumor growth, as previously observed. However, they exhibited a more significant degree of tumor regression compared to that in the nontreatment group. Overall, all the tested animals showed complete tumor regression at both 30 and 60 days following the first treatment. However, animals receiving single agent therapy experienced continuous tumor growth, while having significantly lower tumor growth patterns compared to those in untreated animals. Complete tumor regression and recurrence rates are shown in Figure 5C. Among 10 tumor-bearing animals receiving combination therapy, all showed complete tumor regression (100% tumor regression rates) at both 30 and 60 days following the first treatment. Furthermore, all tumor-regressed animals displayed no tumor recurrence at 60 days after the first treatment (0% tumor recurrence rates). These effects were also observed at 90 days following the first treatment (data not included). These animals were denoted as tumor-cured mice. Taken together, these results demonstrate that in combination with E7 peptide vaccine regimens, optimal use of gemcitabine to avoid Ag-specific CTL inhibition is critical to achieve tumor cure. Thus, this result highlights the importance of the optimization of combination therapy using cytotoxic drugs and therapeutic vaccines for achieving complete tumor control after discontinuation of antitumor therapy.
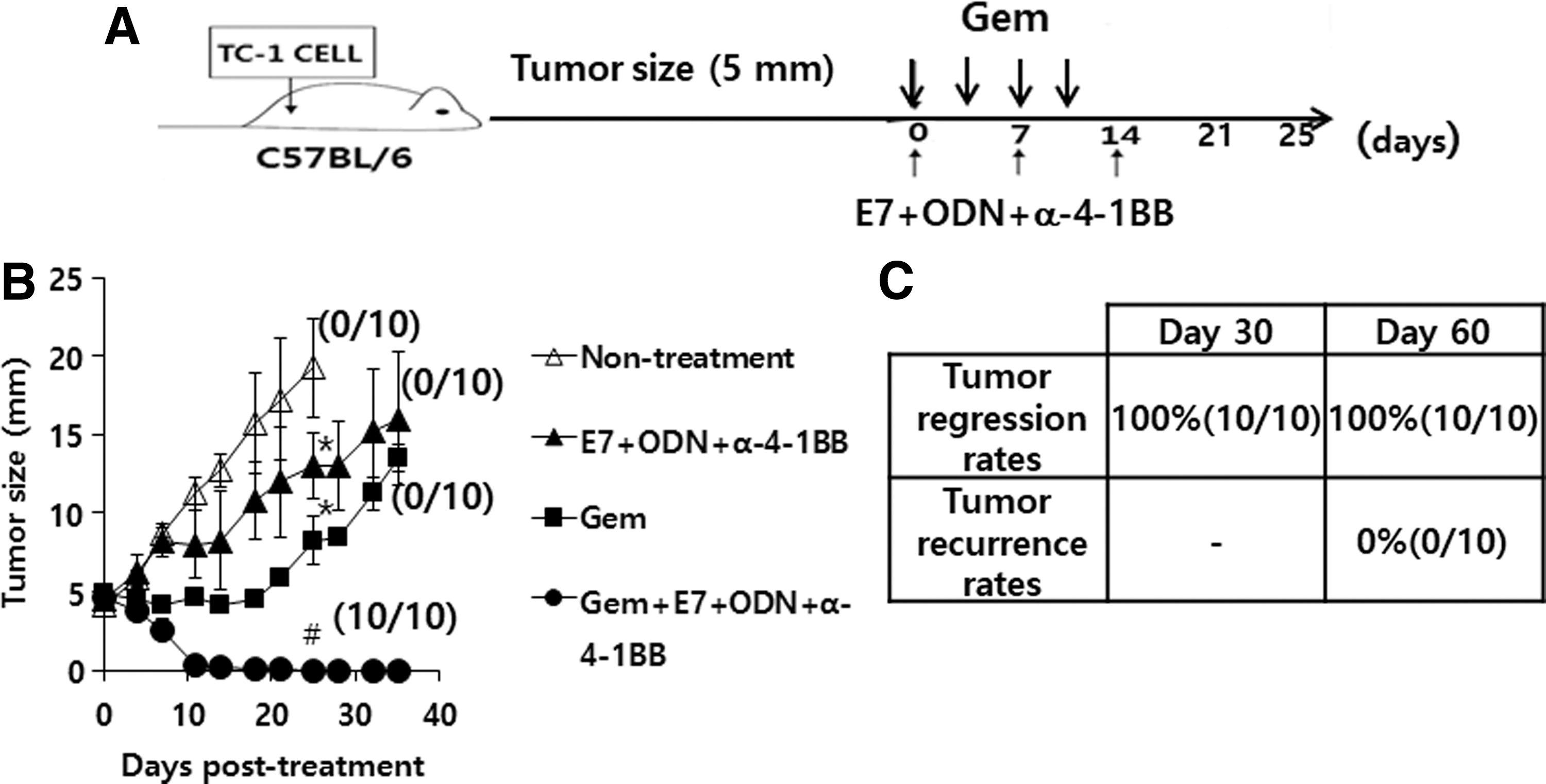
Tumor control and recurrence rates of tumor-bearing mice by combined therapy with gemcitabine plus E7 peptide vaccine regimens (E7 peptides+ODN+anti-4-1BB Ab).
Long-term antitumor memory responses were responsible for complete tumor regression without any recurring tumors
Next, we investigated the mechanism(s) underlying the prevention of tumor recurrence by treatment with gemcitabine plus E7 peptide vaccine regimens in tumor-cured animals. For this test, tumor-cured animals were re-challenged with parental TC-1 tumor cells on the right flank and irrelevant MC38 tumor cells on the left flank, in parallel with age-matched naive control mice. Tumor sizes were measured over indicated time points. As shown in Figure 6A, both types of tumor cells (TC-1 on the right flank and MC38 on the left flank) formed tumors and continued to grow in all five naive control mice. In the tumor-cured animals, however, TC-1 tumor cells formed tumors on the right flank, which regressed thereafter in four of the five tumor-cured animals, while MC38 tumor cells formed tumors on the left flank and continued to grow in all five mice (Fig. 6B). Thus, these data suggest that optimized gemcitabine therapy in combination with E7 peptide vaccine regimen therapy results in tumor eradication in animals through long-term tumor-specific memory responses, which appears to be responsible for prevention of tumor recurrence in these animal groups.

Long-term antitumor memory responses in TC-1 tumor-cured animals. Tumor-cured animals
Discussion
In this study, we observed that the H-2b-restricted CTL epitopes, E7 peptides, were immunogenic in producing Ag-specific CD8+ CTL lytic activity when injected into animals. They were also more immunogenic when co-injected with the TLR9 ligand, CpG-ODN. This is in agreement with previous findings in B16 and TC-1 models (Kim et al., 2002; Jérôme et al., 2006). It has been known that CpG-ODN activates APCs for the production of IL-12, which induces enhancement of CD8+ CTLs (Warren et al., 2000; Kim et al., 2004). We also observed that the addition of anti-4-1BB Abs to E7 peptides plus CpG-ODN regimens resulted in the enhancement of both Ag-specific CTL lytic and antitumor therapeutic activity. This result is in alignment with our previous findings in a B16 melanoma model (Sin et al., 2013). These findings are also supported by those of previous studies. For instance, 4-1BB signaling activates CD8+ T cells and increases their life span by eliciting the expression of antiapoptotic molecules (Bcl-Xl and Bfl-1) (Cooper et al., 2002; Lee et al., 2002). Moreover, augmented antitumor CTL activity may be ascribed to the blocking effects of anti-4-1BB on reverse signaling through 4-1BB ligand, as has been recently reported (Kang et al., 2017). On the other hand, a recent failure in treating pancreatic cancer patients with telomerase peptide vaccines underscores the importance of optimizing their immune responses (Staff et al., 2014). Taken together, these results suggest that CpG-ODN activates APCs for increasing Ag-specific CD8+ CTLs, which may be subsequently stimulated by 4-1BB signaling in this peptide vaccine model.
It has been known that CD8+ CTLs are primarily associated with the control of HPV 16-associated diseases in humans (Nakagawa et al., 1997, 2000). The importance of CTLs has also been demonstrated in numerous animal model systems. For example, we previously demonstrated using a TC-1 model that Ag-specific CD8+ CTLs act as the major effector cell subsets responsible for protection from E6-/E7-expressing tumor growth (Kim et al., 2002; Bae et al., 2007; Ye et al., 2007). In this sense, the induction and enhancement of tumor antigen-specific CD8+ CTLs have been a mainstay in tumor immune therapy. However, in this study, using large established TC-1 tumors, we failed to see complete tumor eradication by CTL lytic activity driven by E7 peptides, CpG-ODN, and anti-4-1BB Abs in combination. Collectively, these data imply that there may be a limit to the tumor sizes that may be controlled by immune therapy.
We also observed that systemic delivery of gemcitabine led to the removal of small established TC-1 tumors (mean tumor size, 2 mm). This is in line with the observation in the CT26/HER2 tumor model (Danishmalik et al., 2017b). On the other hand, when animals with large established TC-1 tumors (mean tumor size, 5 mm) were treated systemically with gemcitabine, they displayed a degree of tumor growth inhibition without any tumor regression, suggesting that there may be a limit to the tumor sizes, which may be removed by systemic delivery of the cytotoxic drug, gemcitabine. This hypothesis is supported by our finding that gemcitabine failed to eradicate established tumors at both doses (75 and 37.5 μg/g body weight). Moreover, the antitumor effects of gemcitabine appeared to be mediated by its direct cytotoxicity to tumor cells, as determined by the CD8+ and NK cell depletion assay. Moreover, none of the Ag-specific CTL responses were detectable in tumor-bearing animals receiving gemcitabine therapy, underscoring the notion that gemcitabine may be unable to elicit endogenous CTL responses. Contrary to previous reports, we were unable to see any decrease in the proportions of MDSCs among the CD45+ cell populations in the tumor tissues of animals receiving gemcitabine therapy. This result suggests that gemcitabine-mediated MDSC depletion is not associated with tumor control in this model. This is in agreement with our previous finding (Danishmalik et al., 2017b). In that study, anti-Gr-1 Ab treatment, but not gemcitabine treatment, resulted in a decrease in MDSC populations in both lymph nodes and tumor tissues, suggesting that gemcitabine is indeed unable to remove MDSCs. However, this is not in line with the findings of other studies (Suzuki et al., 2005; Mundy-Bosse et al., 2011). The reasons for this discrepancy are presently unclear; however, the differences in the doses of gemcitabine tested may be a factor involved. On the other hand, we observed that gemcitabine treatment led to the inhibition of the lytic function of Ag-specific CTLs driven by E7 peptide vaccine regimens, likely dampening their antitumor effectiveness. In this context, gemcitabine likely inhibits metabolically active immune cells, including Ag-specific CD8+ CTLs, systemically and in the tumor tissues, thus leading to both dramatically reduced CTL activity and tumor recurrence in animals receiving combination therapy. Furthermore, none of the tested animals displayed complete tumor regression through systemic delivery of gemcitabine (75 μg/g body weight) plus immunization with E7 peptide vaccine regimens. In contrast, complete tumor regression was observed when tumor-bearing mice were treated with E7 peptide vaccine regimens plus a twofold decreased dose of gemcitabine (37.5 μg/g body weight). In these tumor-regressed mice, however, tumors recurred after discontinuation of therapy, suggesting that the suppression of Ag-specific CTL functions by gemcitabine is likely associated with tumor recurrence. This result highlights the notion that Ag-specific CTL responses may be negatively affected by gemcitabine treatment, resulting in tumor recurrence in animals with complete tumor regression. This hypothesis is further supported by our subsequent findings, wherein reduced frequency of gemcitabine therapy resulted in complete tumor regression without any recurring tumors in animals receiving E7 peptide vaccine regimens. In addition to this antitumor therapeutic efficacy, a degree of long-term antitumor memory against parental TC-1 cells was observed in the tumor-cured mice, which appeared to be responsible for a lack of tumor recurrence in this model. Previously, others, including our group, reported that therapeutic synergy between chemotherapy and immunotherapy is mediated primarily by increased sensitivity of drug-exposed tumor cells to CTL-mediated tumor cell killing (Bae et al., 2007; Tseng et al., 2008; Ramakrishnan et al., 2010; Bauer et al., 2014; Danishmalik and Sin, 2017a). In this study, we also observed that gemcitabine-treated TC-1 tumor cells became more susceptible to CTL-mediated killing in vitro (data not included), which appeared to be the major mechanism underlying the therapeutic synergy between these two therapeutic modalities. Collectively, these studies demonstrate that gemcitabine is indeed inhibitory to Ag-specific CTL responses. Thus, the optimal use of gemcitabine to avoid Ag-specific CTL inhibition may be critical for achieving tumor cure when combined with the E7 peptide vaccine regimens.
In summary, our data demonstrated that as E7 peptide vaccine adjuvants, both CpG-ODN and anti-4-1BB functioned to enhance Ag-specific CTL lytic and antitumor therapeutic activity. Systemic delivery of gemcitabine resulted in tumor growth inhibition by its direct cytotoxicity. As gemcitabine did not induce endogenous CTL responses and influence MDSCs, but inhibited Ag-specific CTL responses during combination therapy, optimal use of gemcitabine to avoid Ag-specific CTL inhibition appeared to be critical for achieving complete tumor regression without any recurring tumors even after discontinuation of therapy, possibly mediated by long-term tumor-specific memory responses. These findings may have clinical implications for designing peptide-based therapeutic vaccines for patients with cancer undergoing chemotherapy.
Footnotes
Acknowledgments
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF), funded by the Ministry of Education, Science and Technology (NRF-2015R1D1A1A01058704). This study was supported by 2017 Research Grant from Kangwon National University (No. 520170425).
Disclosure Statement
No competing financial interests exist.