
Abstract
The intronic microRNA, miR-125b, plays a vital role in promyelocytic and hematopoietic stem cells, and in the development and apoptosis of cancer cells. In this study, we showed that miR-125b regulates granulosa cell (GC) apoptosis in the yak ovary. Bioinformatic analyses and luciferase reporter assays demonstrated that bone morphogenetic protein receptor type 1B (BMPR1B) is an miR-125b target. miR-125b overexpression induced apoptosis in yak GC, and affected the mRNA and protein expression of BMPR1B and the ratio of Bcl2/Bax. Silencing of miR-125b decreased the rate of yak GC apoptosis and increased the ratio of Bcl2/Bax. In addition, the effects of an miR-125b inhibitor were overturned by cotransfection with siRNA-BMPR1B2 (siRNA-299) in yak GC. Together, these results demonstrated that miR-125b regulates GC apoptosis in the yak ovary by targeting BMPR1B.
Introduction
Y
In the 1960s, Urist determined that bone morphogenetic proteins (BMPs) induce bone formation (Urist, 1965). BMPs belong to the transforming growth factor-beta (TGF-β) superfamily (Heldin and Moustakas, 2012); members of this group are involved in the development of GCs and follicle maturation, such as BMP4, BMP7, BMP15, BMP1A, and BMPR1B (Vitt et al., 2000; Varani et al., 2002; Moore et al., 2003; Su et al., 2004). The BMPR1B gene is expressed in each stage of oocyte development based on in situ hybridization (Wilson et al., 2001). In addition, the expression of BMPR1B mRNA in large follicles is higher than that in small follicles (Chen et al., 2009; Lima et al., 2012). The BMPR1B has a serine/threonine protein kinase region and acts as a receptor for BMP ligands (Mulsant et al., 2001) during the regulation of bone differentiation and the development of ovarian follicles (Shimasaki et al., 2003). A recent study demonstrated that in Booroola Merino sheep carrying the BB genotype (FecB locus), follicle size was maintained and apoptosis was reduced (Regan et al., 2015; Souza and Baird, 2006).
microRNAs (miRNAs) are important regulatory factors for cell apoptosis, differentiation, and proliferation. Liu et al. (2014) reported that apoptosis of pig GC was promoted by overexpression of miR-26b (Liu et al., 2014). The let-7 g directly targets the TGFBR1 gene to promote GC apoptosis in pig ovary (Zhou et al., 2015). miR-125b is an important miRNA, and many studies have focused on its biological functions in cell proliferation and differentiation (Wu et al., 2012), and in cancer cells (He et al., 2013). Overexpression of miR-125b suppresses proliferation of ovarian, bladder, breast, and liver cancer cells (Hofmann et al., 2009; Huang et al., 2011; Zhang et al., 2011). miR-125b directly targets Bc13 and placental growth factor (Pogue et al., 2010). miR-125b also regulates the expression of proapoptotic proteins, such as p53, Bak1, Puma, and Bmf (Shi et al., 2007; Le et al., 2009; Xia et al., 2009; Shi et al., 2011). In addition, Hu et al. (2016) reported that miR-125b expression was downregulated in highly prolific sheep (Hu et al., 2016). However, few studies have explored the roles of miR-125b in GC proliferation and apoptosis in animals. In this study, we chose miR-125b, an miRNA that significantly upregulated during yak follicular atresia, and a candidate regulatory miRNA of BMPR1B, which is sought to explore the molecular mechanism in yak ovarian GCs.
Materials and Methods
Ethics statement
All experiments were conducted in accordance with the guidelines of the regional Animal Ethics Committee and were approved by the Institutional Animal Care and Use Committee of XiZang Agricultural and Animal Husbandry College. The institutional certification number is 12540000MB0P013721.
Animals
A total of 10 female yaks weighing 250–300 kg and of 6–7 years of age were selected from the yak farm belonging to the National Research Centre on Yak, situated 2750 m above sea level in Linzhi, Tibet. The animals were slaughtered and healthy yak ovaries were obtained. The animals were apparently healthy and free from any anatomical reproductive disorders, and completed 2 years of postpartum period. The yak ovaries were harvested and stored in physiological saline at 38°C before experimental analyses.
Cell culture
Yak ovaries were transported to the laboratory at 38°C. They were washed using saline and alcohol. Follicles were assessed according to appearance and form (McBride et al., 2012; Moor et al., 1978). The appearance of a healthy follicle is clear, with a cumulus oocyte complex on the follicular wall, and a complete granular cell layer. Atretic follicles are cloudy, and it is difficult to distinguish cumulus oocyte complexes and the GC layer folds. After follicle assessment, follicles were broken down using a homemade collection tube: the bottom of the tube has a gap in the middle for follicular fluid to pass through while the follicular tissue is filtered out. Then the collection tube was inserted into a 1.5 mL centrifuge tube and centrifuged for 5 min at 2000 g. The GCs and follicular fluid fell into the bottom of the centrifuge tube, and the follicle wall remained in the collecting tube. The GCs were then washed three times with PBS and obtained after centrifugation for 5 min (2000 g). GCs were cultured in modified Eagle's medium (MEM; Sigma) containing 10% fetal bovine serum (FBS; Atlanta Biologicals), 1% penicillin/streptomycin/L-glutamine (Mediatech), and 1% glutamine. The cells were cultured in an incubator containing 5% CO2 at 37°C.
In vitro cultured yak GCs showed typical epithelial-like cell growth characteristics. After inoculation for 24 h, the GCs moved closer to each other, and the cells contacted each other through extended pseudopods. After 72 h, the cell junctions were tight and microscopic intracellular small particles were observed.
HEK293 cells were grown in Dulbecco's modified Eagle's medium (DMEM; Sigma) containing 10% FBS and 1% penicillin/streptomycin/L-glutamine. HEK293 cells were also cultured in 5% CO2 at 37°C.
Hematoxylin and eosin staining
Yak ovaries were washed and fixed in 4% paraformaldehyde (in DEPC water) for 2–12 h. Then, they were immersed in wax and embedded, and the paraffin was sliced using a sheeter (OPJ-1B; Tianjin Tianli Aviation Electromechanical Co., Ltd.). The sections were stained in hematoxylin for 3–5 min. After hematoxylin staining, the slices were dehydrated with alcohol and stained in eosin for 5 min. The nucleus was stained in blue and the cytoplasm in red by hematoxylin and eosin, respectively.
Cell morphology observation
Pictures of follicles were obtained using a camera (Leica D-LUX typ109). The cultured cells were photographed with a BX51-DP71 (Olympus, Japan) upright microscope. Photoshop CS3 software was used to process pictures.
Bioinformatic analysis
miRNAs targeting BMPR1B were predicted by TargetScan a , miRDB b , and Microcosm c tools. miRNAs that are broadly conserved among vertebrates were selected in the TargetScan prediction results.
Cell transfection
Cells were grown to 75–80% confluence in 12-well or 6-well plates, and then, mimics, plasmids, or controls were transfected using Lipofectamine 2000 reagent (Invitrogen) according to the manufacturer's instructions. The synthetic miR-125b mimics, miR-125b inhibitor, negative control, siRNA-BMPR1B1 (siRNA-520), siRNA-BMPR1B2 (siRNA-299), and siRNA-control were obtained from GenePharma (Shanghai, China). Primer sequences are listed in Table 1.
Plasmids
The yak BMPR1B coding sequence was amplified by PCR using forward and reverse primers containing BamH I and Xho I sites, respectively. The PCR products were inserted into a pc-DNA3.1 vector (Invitrogen). The inserted sequence was verified by sequencing. To verify whether miR-125b targets BMPR1B in vitro, fragments of miR-125b target sites in BMPR1B 3′-UTR were cloned between the Sac I and Xho I sites. The PCR products were inserted into pmirGLO Dual-Luciferase Vector that was digested with Sac I and Xho I. The binding sites were mutated from CAGGGA to AGAAAG using a site-directed mutagenesis kit (Takara) according to the manufacturer's instructions. The vectors were named 3′-UTR-BMPR1B-WT and 3′-UTR-BMPR1B-MT. Primer sequences are listed in Table 2.
Digestion sites are underlined.
Dual-luciferase assay
HEK293 cells were grown to 75–80% confluence in 12-well plates, and then, 3′-UTR-BMPR1B-WT, 3′-UTR-BMPR1B-MT, pmirGLO Dual-Luciferase Vector (control), or miR-125b mimics were cotransfected using Lipofectamine 2000 reagent (Invitrogen) according to the manufacturer's instructions. After 24 h, cells were harvested, and double luciferase activity was examined with the dual-luciferase assay system (Promega).
Apoptosis assay
Yak GC apoptosis was examined using the Annexin 155 V-FITC/propidium iodide apoptosis kit (KeyGen, Nanjing, China) according to the manufacturer's protocol, and performed as previously described (Liu et al., 2014).
Quantitative RT-PCR
Total RNA was extracted from yak GCs using the TRIzol reagent (Invitrogen) according to the manufacturer's instructions and then reverse transcribed to cDNA using PrimeScript™-RT Master Mix (TaKaRa, Dalian, China). RT-PCR was conducted using the SYBR Green Master Mix (Vazyme Biotech, Nanjing, China) and the results were analyzed using the 2−ΔΔCt method (Du et al., 2016). Each experiment was repeated three times and the data are shown as mean ± SEM. Primer sequences are listed in Table 1.
Western blot
Yak GCs were collected at 48 h post-transfection, and whole-cell lysates were prepared in RIPA buffer (Sigma) (Liu et al., 2014). The total protein concentration was quantified using the BCA protein assay kit (Invitrogen), and each sample was separated in a 4–12% SDS-PAGE gel (Invitrogen) and then transferred onto a nitrocellulose membrane. The membrane was blocked by incubating for 2 h at room temperature with 2% bovine serum albumin (BSA) in 20 mM Tris-HCl, 150 mM NaCl, and 0.05% Tween 20. The membrane was then incubated at 4°C overnight with a diluted (1:1500) monoclonal anti-GAPDH antibody as an internal control, followed by a secondary antibody (1:2000) for 2 h at room temperature: anti-BMPR1B (ab155058; Abcam, United Kingdom) and anti-GAPDH (ab9482; Abcam), all of these antibodies were diluted with 0.25% BSA. After washing, the membrane was incubated with the appropriate secondary antibody and bands were visualized with an enhanced chemiluminescence system.
Statistical analysis
SPSS version 20.0 (SPSS, Inc., Chicago, IL) was used for statistical analysis. t-Test and ANOVA were used to assess statistical significance. p < 0.05 was considered statistically significant. All data are expressed as mean ± SEM.
Results
BMPR1B inhibits apoptosis in yak GCs
A recent study reported that Booroola Merino sheep carried the BB genotype (FecB locus), which meant that follicle size was maintained and apoptosis was reduced (Regan et al., 2015). Based on this study, we investigated BMPR1B regulation in yak GC apoptosis using siRNA to knock down BMPR1B expression. qPCR and Western blot analysis showed that the BMPR1B mRNA and protein (p < 0.01) levels significantly decreased on siRNA-BMPR1B2 (siRNA-299) transfection compared with control (Fig. 1A, 1B). Furthermore, flow cytometry analysis showed that the apoptosis rate of yak GC was significantly increased (p < 0.01) in the siRNA-BMPR1B group compared with the siRNA-control group (Fig. 1C), and the ratio of Bcl2/Bax was downregulated (p < 0.01) (Fig. 1D). To further demonstrate the role of BMPR1B in yak GC apoptosis, we constructed the yak BMPR1B overexpression vector, pcDNA3.1-BMPR1B. Transfection of pcDNA3.1-BMPR1B into yak GCs increased BMPR1B mRNA and protein expression (p < 0.01) (Fig. 1E, 1F); the yak GC apoptosis rate was suppressed (p < 0.01) (Fig. 1G), and the ratio of Bcl2/Bax was increased (p < 0.01) (Fig. 1H). These results suggested that BMPR1B inhibits yak GC apoptosis.
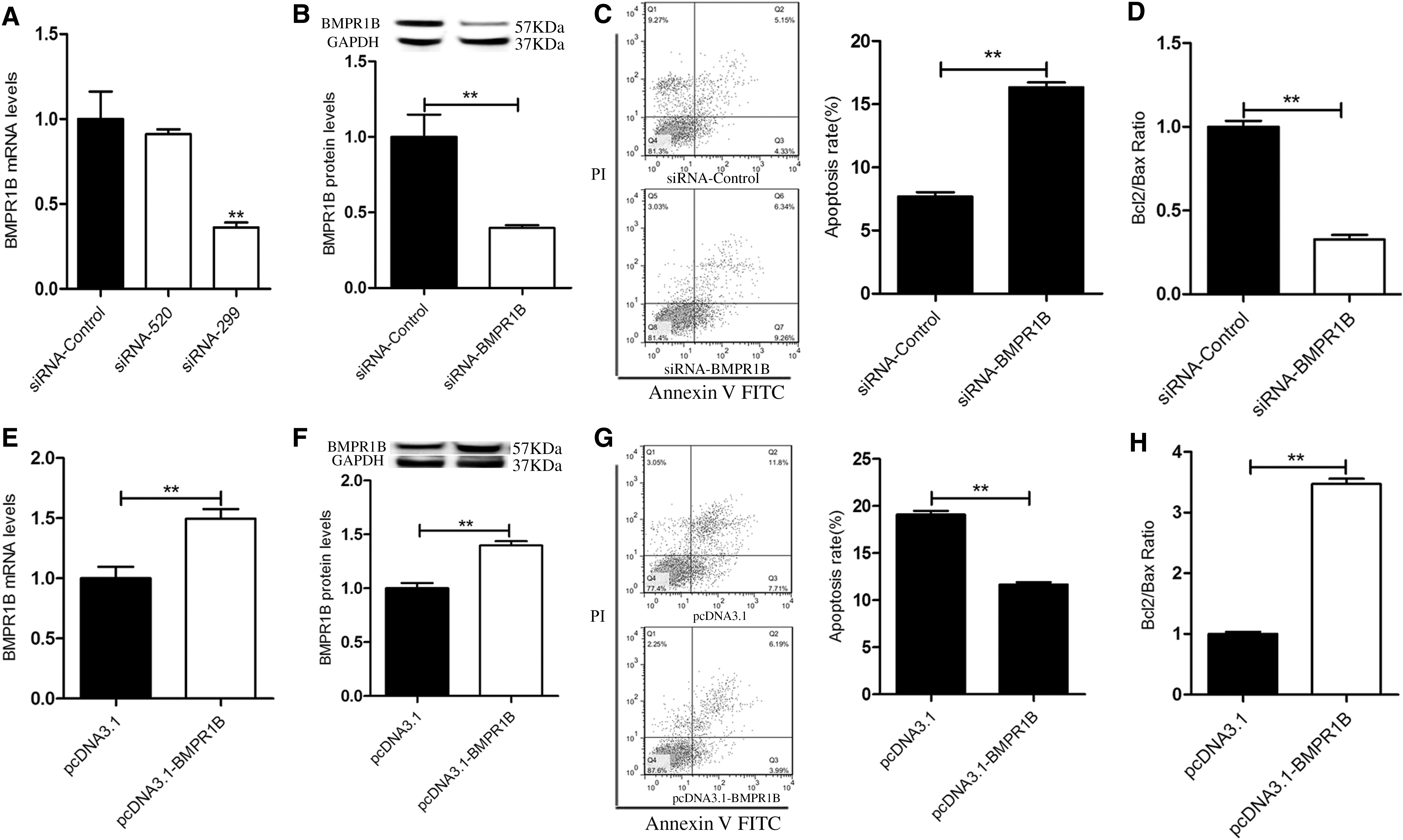
BMPR1B depresses yak GC apoptosis.
BMPR1B is a predicted target of miR-125b
miRNAs usually function by targeting multiple genes, even in the same biological process. To determine which miRNAs regulate BMPR1B, a total of 365 candidate miRNAs targeting BMPR1B were predicted, of which miR-125b and miR-182 were predicted by three algorithms (Fig. 2A). In addition, after transfection with pcDNA3.1-BMPR1B, the expression of miR-125b was downregulated in yak GCs, whereas the levels of miR-182 were unchanged (Fig. 2B). Therefore, we focused on miR-125b in our subsequent analyses. Seed sequences of miR-125b are highly conserved in vertebrates (Fig. 2C). Sequence alignment revealed that vertebrate miR-125b genes share a common seed sequence (Fig. 2D). To verify whether miR-125b targets BMPR1B in vitro, luciferase reporter vectors were constructed, containing 3′-UTR-BMPR1B-WT or 3′-UTR-BMPR1B-MT (Fig. 2E). The luciferase activity of the 3′-UTR-BMPR1B-WT vector was significantly reduced (p < 0.01), but there was no activity in the 3′-UTR-BMPR1B-MT vector on cotransfection with miR-125b mimics (Fig. 2F), indicating that BMPR1B is a true target of miR-125b.

BMPR1B is a target of miRNA-125b.
miR-125b promotes GC apoptosis and affects follicle atresia
In mammals, GC apoptosis is a key mechanism regulating follicle atresia (Krysko et al., 2008). To evaluate the role of miR-125b in yak GCs and follicles, first, as shown in Figure 3A–D, we separated healthy follicles from atretic follicles. Then we cultured the GCs, as shown in Figure 3E and F.
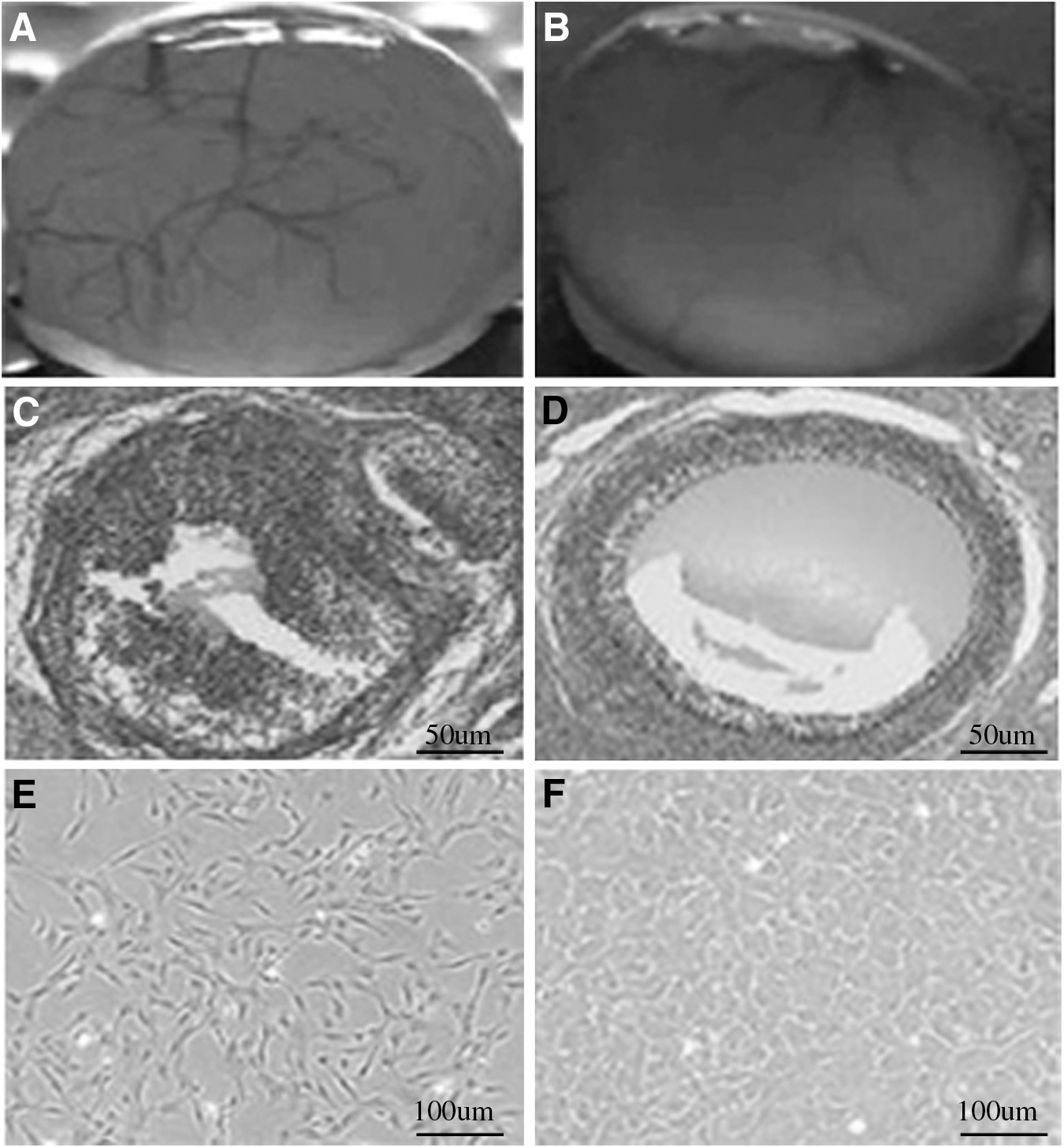
Identification of healthy and atresia follicles.
miR-125b promotes GC apoptosis and follicle atresia
The expression levels of miR-125b and BMPR1B were analyzed by qPCR in healthy and atretic follicles. The BMPR1B mRNA expression was higher in healthy follicles (p < 0.01) (Fig. 4A). miR-125b expression was higher in atretic follicles (p < 0.01) (Fig. 4B), and ratio of Bcl2/Bax was downregulated (p < 0.01) (Fig. 4C). The yak GC apoptosis was promoted by miR-125b (p < 0.01) (Fig. 4D). These results indicated that miR-125b promotes GC apoptosis, and there is a correlation between miR-125b and BMPR1B in yak GCs.
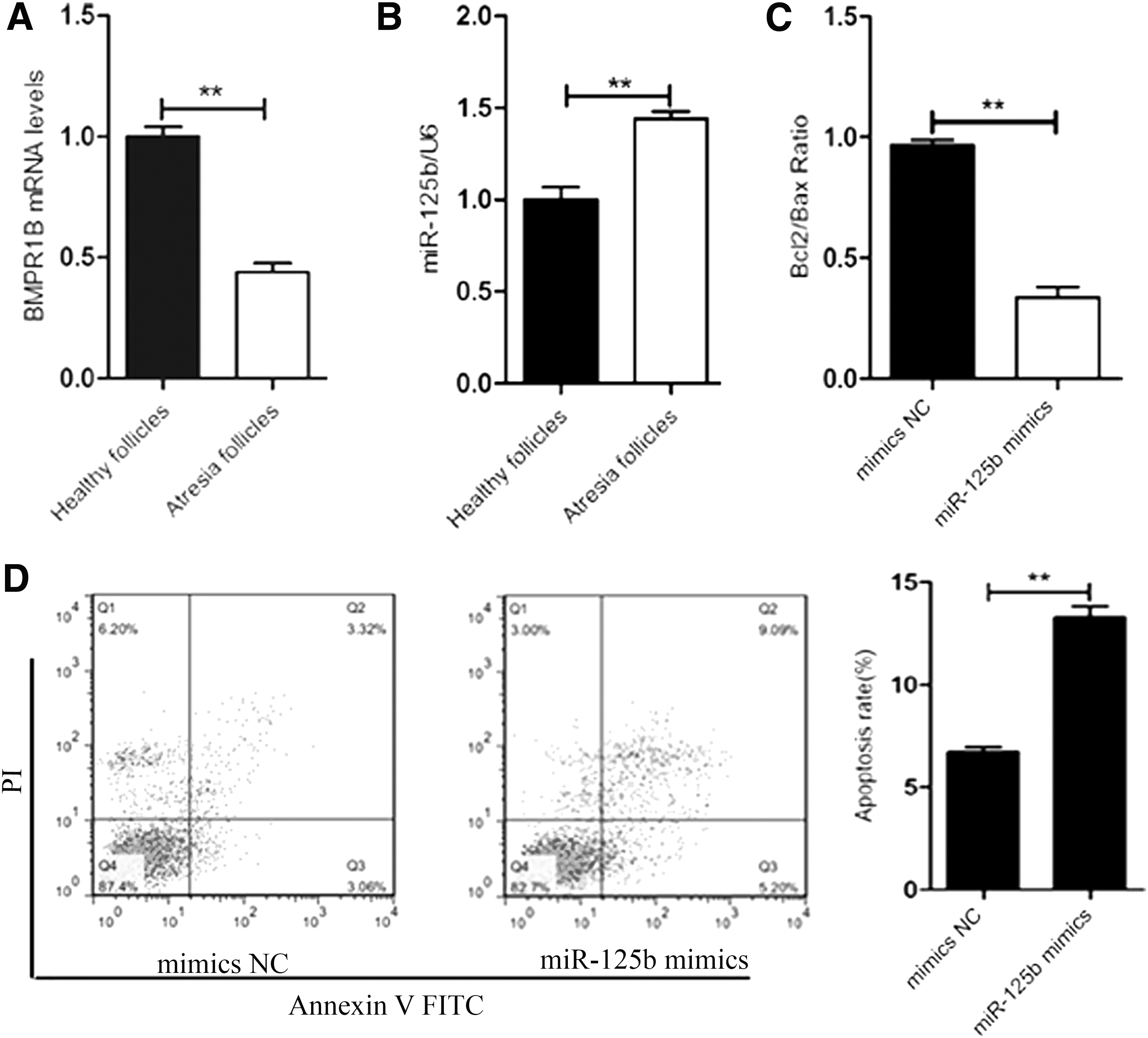
miR-125b promotes yak GC apoptosis.
miR-125b regulates BMPR1B expression in yak GCs
We next explored the correlation between miR-125b and BMPR1B in yak GC. The BMPR1B mRNA and protein levels were evaluated after transfection with miR-125b mimics, miR-125b inhibitor, or control. The results showed that miR-125b mimics downregulated BMPR1B mRNA and protein expression (p < 0.05) (Fig. 5A), while the miR-125b inhibitor obviously upregulated BMPR1B mRNA and protein expression in yak GC, compared with the control group (p < 0.01) (Fig. 5B). These findings indicated that miR-125b targets BMPR1B 3′-UTR regulating its expression.
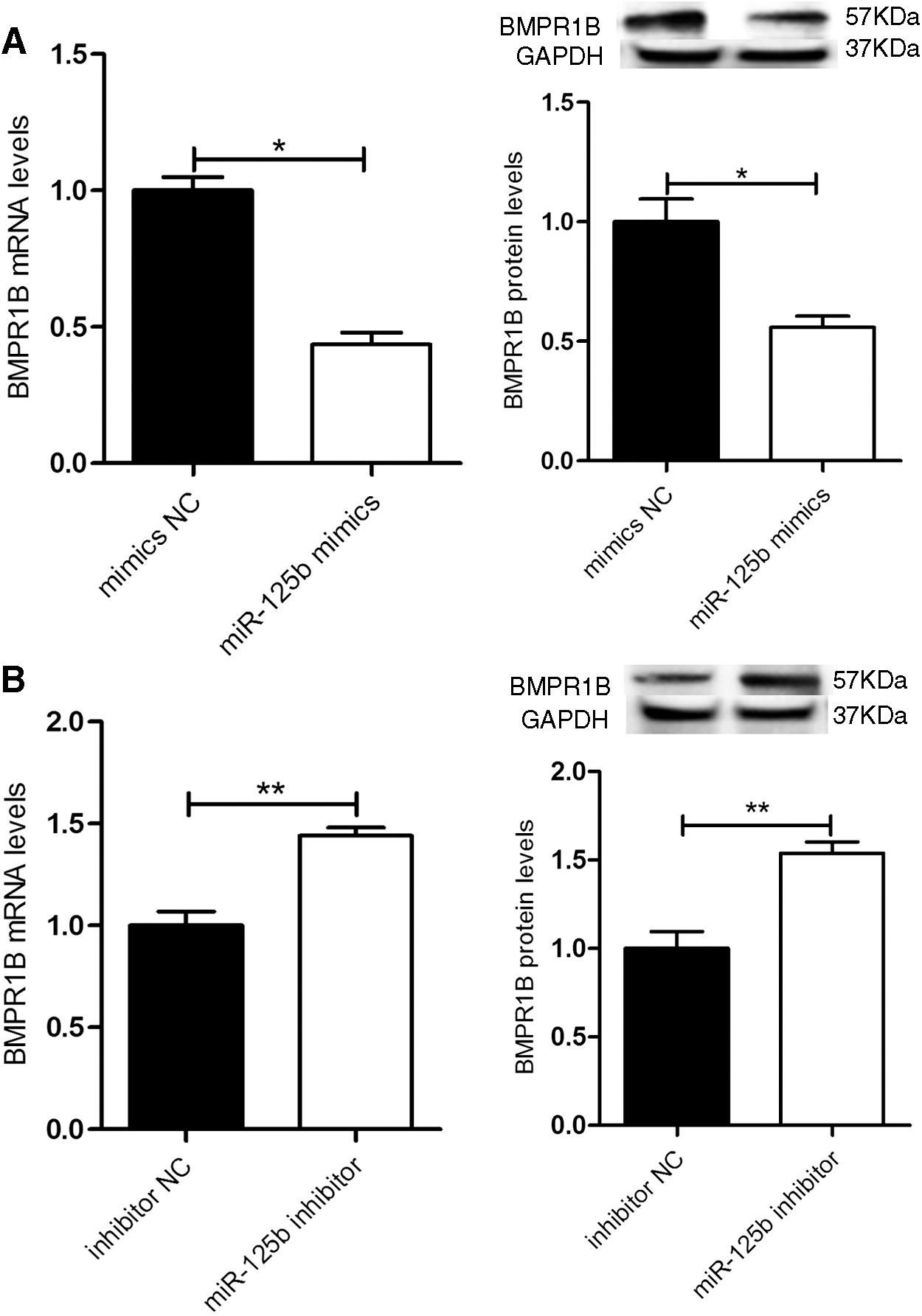
miR-125b decreases expression of BMPR1B in yak GC.
miR-125b regulates apoptosis in yak GC via targeting BMPR1B
We further investigated whether miR-125b regulates apoptosis via BMPR1B 3′UTR in yak GC. After knocking down, miR-125b decreased (p < 0.01) yak GC apoptosis and increased the ratio of Bcl2/Bax (Fig. 6A, B). Then, we knocked down both miR-125b and BMPR1B in yak GC by cotransfecting miR-125b inhibitor and siRNA-BMPR1B2. The results showed that effects of miR-125b inhibitor in yak GC were significantly reversed by knocking down BMPR1B (p < 0.01) (Fig. 6C, D), indicating that miR-125b regulates yak GC apoptosis by targeting BMPR1B.

miR-125b mediates yak GC apoptosis via BMPR1B. Inhibition of miR-125b increases yak GC apoptosis
Discussion
GCs play a vital regulatory role in follicular growth and atresia through the secretion of estrogen and other regulatory factors (Maeda et al., 2007; Moley and Schreiber, 1995). The synthesis and secretion of mucopolysaccharides by GCs (Rabinovici et al., 1992; Xiaguoliang, 2002; Jimenezkrassel et al., 2003) can regulate cell/cell nutrition, exchange of small molecules and transmission of information, and play an important role in the growth, development, and maturation of oocytes (Orisaka et al., 2009; Alpini et al., 1980; Bleil and Wassarman, 1980). However, the miRNA mechanisms that regulate apoptosis by targeting the TGF-β family in the yak ovary have not been fully investigated.
miRNAs can directly target homologous mRNA sequences, leading to transcript degradation and inhibition of protein biosynthesis (Ambros, 2004). A recent study in mice reported that miR-125b is highly expressed perinatally and is specifically located in the ovarian somatic cells, which inhibited somatic cell proliferation in neonatal mouse ovary by impairing the activin/Smad2 signaling by directly targeting Acvr2a, a type II receptor in the TGF-β superfamily (Han et al., 2013). In our study, using bioinformatic analysis, we also found that miR-125b can directly target BMPR1B 3′UTR and downregulate luciferase activity, suggesting that miR-125b has a biological function. MiR-125b is also involved in apoptosis of various cell types, including normal cells and cancer cells. Liang et al. (2010) reported that miR-125b controls cell apoptosis by targeting Mcl-1, Bcl-W, and ALAT1 in bladder cancer cells. In blood monocytes, miR-125b regulates cell apoptosis by targeting BIK and MTP18 (Duroux-Richard et al., 2016), and p53 in lens epithelial cells (Qin et al., 2014). miR-125b was detected at all four stages of follicular and luteal development (McBride et al., 2012). In addition, miR-125b was involved in GC apoptosis and follicular development of mammals, including humans (Sirotkin et al., 2010). Notably, miR-125b was also downregulated in hepatocellular carcinoma, and its expression level was positively correlated with the apoptosis rate (Alpini et al., 2011; Tsang et al., 2014; Hubble et al., 2014). Taken together, these data suggested that miR-125b promotes apoptosis. In the present study, qRT-PCR results revealed that miR-125b expression was lower in healthy follicles than in atretic follicles, and apoptosis was promoted by miR-125b. Moreover, overexpression of miR-125b downregulated the ratio of Bcl2/Bax in yak GC. These results indicated that miR-125b can promote apoptosis of GC in yak ovaries.
BMPR1B is a necessary growth factor that initiates follicular development in ovaries and regulates the ovulation rate (Moore et al., 2003; Juengel et al., 2004; Paradis et al., 2008). BMPR1B also plays an essential role in osteoblast differentiation and cell apoptosis (Liu et al., 2012), which are required for maintaining healthy female reproductive function (Yi et al., 2001). In this study, we demonstrated that BMPR1B expression was high in healthy follicles, but lower in atretic follicles. In addition, it was reported that BMPR1B null mice exhibited defects in cumulus expansion and the increase in BMPR1B mRNA and protein abundance during the periovulatory period in GC, which suggested that BMPR1B gene can regulate the development of follicles by regulating GCs (Liu et al., 2012). In our study, knock down of BMPR1B really increased the apoptosis rate of yak GC, and the ratio of Bcl2/Bax was downregulated. These results indicated that BMPR1B suppresses yak GC apoptosis. We also found that mRNA and protein levels of BMPR1B were downregulated by miR-125b. In addition, yak GC apoptosis was decreased by the transfection miR-125b inhibitor, and the ratio of Bcl2/Bax was increased. However, when we knock down both miR-125b and BMPR1B in yak GC, the effects of miR-125b inhibitor in yak GC were significantly reversed. These results indicated that miR-125b can regulate apoptosis of yak GC by targeting BMPR1B 3′UTR.
In conclusion, our data suggest that miR-125b enhances yak GC apoptosis. miR-125b targets BMPR1B to downregulate its mRNA and protein expression. Thus, we propose a novel pathway through which miR-125b targets BMPR1B to enhance yak GC apoptosis.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (31460604), the comprehensive strength of the Midwest colleges and universities to enhance the plan, Tibet Institute of Animal Husbandry Animal Genetics and Breeding Discipline Construction Project (502000105), and the National Beef Yak Industrial System Project (CARS-37).
Disclosure Statement
We declare that we have no financial and personal relationships with other people or organizations that could inappropriately influence our work.