
Abstract
Metastasis of ovarian cancer is regulated by microRNAs. This study focused on the effects of miR-30a-5p on ovarian cancer migration and invasion. Our results showed that the miR-30a-5p and mucin type O-glycan biosynthesis are closely related to ovarian cancer, and that miR-30a-5p was downregulated in ovarian cancer cells. miR-30a-5p overexpression reduced cell viability and inhibited migration and invasion in HO-8910 and HO-8910PM cells. S phase kinase-associated protein 2 (SKP2), B cell lymphoma 9 (BCL9), and NOTHC1 are direct target genes of miR-30a-5p. MTDH, SKP2, BCL9, and NOTCH1 genes were overexpressed in ovarian cancer cells, and they are direct target genes of miR-30a-5p. miR-30a-5p overexpression inhibited epithelial–mesenchymal transition (EMT) process, while upregulation of SKP2, BCL9, and NOTCH1 gene expression levels reduced the inhibition of EMT process by miR-30a-5p. miR-30a-5p was lowly expressed in ovarian cancer, and such a phenomenon is related to ovarian cancer metastasis. miR-30a-5p might inhibit the migration and invasion of ovarian cancer cells by downregulating the expression of SKP2, BCL9, and NOTCH1 genes.
Introduction
Ovarian cancer is one of the most common malignant tumors, and it has the highest mortality among all malignant tumors in women (Barnett, 2016). Five-year survival rate of patients with ovarian cancer is <50% (Ledermann et al., 2012). Ovarian cancer has a strong metastatic ability, which leads to distant metastasis by the time the disease is discovered and increases the challenges of treatment (Gabra, 2014).
Epithelial–mesenchymal transition (EMT) refers to the conversion of an epithelial cell to a mesenchymal cell. The loss of epithelial function and the acquisition of mesenchymal function during EMT are accompanied by an increase in migration and invasion properties, which are the two most important causes of ovarian cancer metastasis (Kalluri and Weinberg, 2009; Gonzalez and Medici, 2014). EMT of ovarian cancer cells accepts a variety of signaling pathways and gene regulation (Huang et al., 2012; Grassi et al., 2017; Weingarten et al., 2018). Studying the mechanism of regulating ovarian cancer metastasis is significant to the treatment of ovarian cancer.
MicroRNAs (miRNAs) are a category of endogenous, non-coding RNA molecules of length ∼18–24 nucleotides. miRNAs can regulate protein expression by inhibiting or inducing the degradation of messenger RNAs (mRNAs) through specifically binding to 3′-untranslated region (UTR) of the mRNAs (Cullen, 2004; Nilsen, 2007). A study showed that ∼30% of genes can be regulated by miRNA (Deng et al., 2008). Recent studies demonstrated that the occurrence and development of ovarian cancer is regulated by various miRNAs (Kobayashi et al., 2014; Xiang et al., 2014; Yu et al., 2014; Xia et al., 2015), and that some miRNAs could inhibit the metastasis of ovarian cancer by regulating EMT (Sun et al., 2016; Tan et al., 2016).
Therefore, this study summarized miRNAs closely related to ovarian cancer and used miRPath v3 to analyze the pathways associated with these miRNAs. Then we analyzed the miRNAs associated with EMT of ovarian cancer. The aim of this study was to investigate the mechanism of ovarian cancer metastasis and to provide evidence and strategies for its diagnosis and treatment.
Materials and Methods
Bioinformatics analysis
Pathways associated with miRNAs were analyzed using miRPath v3 as previously reported (Vlachos et al., 2015).
Cell culture and transfection
The human normal ovarian epithelial cells IOSE80 and ovarian cancer cell lines OVCAR-3, HO-8910PM, HO-8910, Caov-3, SKOV-3, A2780, COC1, and OV-90 were purchased from ATCC. The cells were cultured in RPMI 1640 medium containing 10% FBS at 37°C in an incubator with 5% CO2. Culture-related reagents were purchased from GIBCO Invitrogen.
The miR-30a-5p mimic was amplified with the sequence: 5′-UGUAAACAUCCUCGACUGGAAG-3′. The miR-30a-5p mimic were transfected into ovarian cancer cells with Lipofectamine 2000 (Invitrogen, Carlsbad, CA) following the manufacturer's protocol. Empty vector was used as negative control.
The targets of S phase kinase-associated protein 2 (SKP2), B cell lymphoma 9 (BCL9), and NOTHC1 coding sequences were subcloned into pcDNA3.1 (Sangon Biotech, China) to construct pcDNA expression vectors. SKP2, BCL9, and NOTHC1 transfections were performed using Lipofectamine 2000 according to the instructions. The empty plasmid was used as control.
Cell counting kit-8 assay
The cell counting kit-8 (CCK-8) assay was performed to test cell viability, and the kit was purchased from Tongren (Japan). Cells were preincubated at 37°C in a 5% CO2 atmosphere, and then CCK-8 reagent was added into the wells and cultured at 37°C with 5% CO2 for 4 h. The optical density (OD) of each well at 450 nm was measured using a microplate reader (ELX 800; Bio-Teck).
Scratch assay
A black marker pen was used to draw a horizontal line across the hole behind the six-well plate. 5 × 105 cells were added to each well and incubated overnight. The pipette tip was used for vertical scratching, and the cells were incubated in serum-free medium (Oringeng) for 24 h. The image was processed using Image J software (Rawak Software, Inc., Germany) and was calculated for the proportion of relative wound closure.
Transwell assay
Transwell chamber and related reagents were purchased from Corning. 500 pL of whole culture was added in the lower chamber, and 5 × 104 cells were then added and cultured for 4 h. The upper chamber was placed in the lower chamber. Finally, cell at a density of 5 × 104 cells/mL was seeded into the upper chamber. The cells were cultured at 37°C in an atmosphere with 5% CO2 for 6–8 h. After staining with Giemsa stain (Shanghai Gefan Biotechnology Co., Ltd., China), five high-power fields were randomly selected under a microscope and counted for their relative migration rates.
Dual-luciferase reporter assay
miR-30a-5p target gene associated with mucin type O-glycan biosynthesis pathway was predicted on miRTarBase site. Target gene of miR-30a-5p was predicted using Targetscan and miRWalk sites, and a Venn diagram was then mapped. Ovarian cancer cells were co-transfected with the luciferase reporters in combination with miR-30a-5p mimics using Lipofectamine 2000. At 48 h after transfection, luciferase experiment was performed using a dual-luciferase reporter assay kit (Promega, Beijing, China) according to the instructions.
Real-time quantitative PCR analysis
Real-time quantitative PCR (RT-qPCR) was carried out to detect miR-30a-5p, MTDH, SKP2, BCL9, NOTCH1, E-cadherin, N-cadherin, Vimenin, and matrix metalloproteinase 9 (MMP-9) mRNA expression levels. The cells were triturated and lysed, and then the RNA was extracted using an RNA extraction kit (Promega). Reverse transcription kit (TaKaRa, Japan) was used to synthesize cDNA. Reverse transcription reaction condition was set at 37°C for 15 min, and reverse transcriptase inactivation condition was set at 85°C for 15 s. RT-qPCR was performed with the RT-qPCR kit (TaKaRa). PCR was performed by activating the DNA polymerase at 95°C for 5 min, followed by 40 cycles of two-step PCR (at 95°C for 10 s and at 60°C for 30 s) and a final extension at 75°C for 10 min and held at 4°C. RNase-free water was used as the template of negative control. All primers were obtained from Genewiz (Suzhou, Jiangsu, China) and listed in Table 1. The formula 2 −ΔΔCT was adopted to analyze the mRNA expression levels.
The Sequences of Primers
Western blot
Western blot was performed to test protein expression levels. Cells were lysed and the supernatant was collected after centrifugation at 12,000 rpm at 4°C for 15 min. Bicinchoninic acid assay (BCA) was used to determine the protein concentration. SDS-PAGE gel was applied to electrophoresis. A polyvinylidene fluoride membrane (Bio-Rad) was transferred using a Trans-Blot Transfer Slot (Bio-Rad) and blocked with 5% non-fat milk at room temperature for 2 h. Primary antibodies (anti-MTDH, ABclonal, A5887, dilution 1:600; anti-SKP2, Abcam, ab19877, dilution 1:800; anti-BCL9, Abcam, ab37305, dilution 1:700; anti-NOTCH1, Abcam, ab52627, dilution 1:800; anti-E-cadherin, Abcam, ab6528, dilution 1:800; anti-N-cadherin, Abcam, ab18203, dilution 1:800; anti-Vimenin, Abcam, ab92547, dilution 1:600; anti-MMP-9, Abcam, ab38898, dilution 1:800) were added according to the kit instructions, and the samples were shaken at room temperature for 2 h and incubated at 4°C for 12 h. Secondary antibodies (mouse anti-human IgG, Abcam, ab1927, dilution 1:10,000; rabbit anti-human IgG, Abcam, ab6759, dilution 1:8000; rabbit anti-goat IgG, Abcam, ab6741, dilution 1:10,000; goat anti-rabbit IgG, Abcam, ab6721, dilution 1:8000) were added and incubated at room temperature for 1.5 h. Chemiluminescence detection was carried out using enhanced chemiluminescense (ECL) reagent (Huiying, Shanghai, China).
Results
Bioinformatics analysis
Twenty-five miRNAs related to ovarian cancer were obtained by miRPath v3 analysis. The results showed that the mucin type O-glycan biosynthesis pathway is associated with these miRNAs, and that miR-30a-5p is closely related to the mucin type O-glycan biosynthesis pathway (Fig. 1A). According to previous studies (Lee et al., 2009; Maupin et al., 2010; Bou Zgheib et al., 2012), O-glycan biosynthesis pathway is involved in cancer, and O-glycan has the effects of regulating tumor cell metastasis. Thus, it was important to study the relation between miR-30a-5p and EMT.

Expression of miR-30a-5p in ovarian cancer cells.
Expression of miR-30a-5p in ovarian cancer cell lines
By studying the expression levels of miR-30a-5p miRNA in human normal ovarian epithelial cell line and different ovarian cancer cell lines, we found that the expression of miR-30a-5p was significantly downregulated in ovarian cancer cell lines. Moreover, the expression level was relatively high in low metastatic cell line HO-8910, while the expression level was low in high metastatic cell line HO-8910PM (Fig. 1B).
Effects of miR-30a-5p overexpression on ovarian cancer cells
miR-30a-5p was overexpressed in HO-8910 and HO-8910PM cell lines by plasmid transfection, and transfection efficiency was detected by RT-qPCR. The results showed that miR-30a-5p expression level in miR-30a-5p mimic group was significantly higher than that in control group and mock group (Fig. 2A, E). The CCK-8 assay showed that miR-30a-5p overexpression reduced HO-8910 and HO-8910PM cell viabilities and inhibited cell growth (Fig. 2B, F).

Effects of miR-30a-5p overexpression on HO-8910PM and HO-8910 cells.
The effects of miR-30a-5p overexpression on cell migration and invasion were examined using the scratch and transwell assay. The results showed that the migration and invasion ability of HO-8910PM cells was higher than those of HO-8910 cells, and that the relative wound closure in miR-30a-5p mimic group was higher than that in mock group; however, the relative migration rate was lower than that in mock group. This indicated that the miR-30a-5p overexpression inhibited the migration and invasion of HO-8910 and HO-8910PM cells (Fig. 2C, D, G, H).
Prediction and verification of miR-30a-5p target genes
After searching for the target gene of miR-30a-5p in TargetScan, miRWalk, and miRTarBase, 10 common target genes for miR-30a-5p predicted in three databases were found as follows: MTDH, RUNX2, SNAI1, ERG MBNL3, MBNL2, SKP2, BCL9, TAB3, and NOTCH1 (Fig. 3A). The expression levels of these 10 genes in HO-8910 and HO-8910PM cells were detected by RT-qPCR, and we found that MTDH, SKP2, BCL9, and NOTCH1 mRNA were significantly overexpressed in HO-8910 and HO-8910PM cells (Fig. 3B, C).
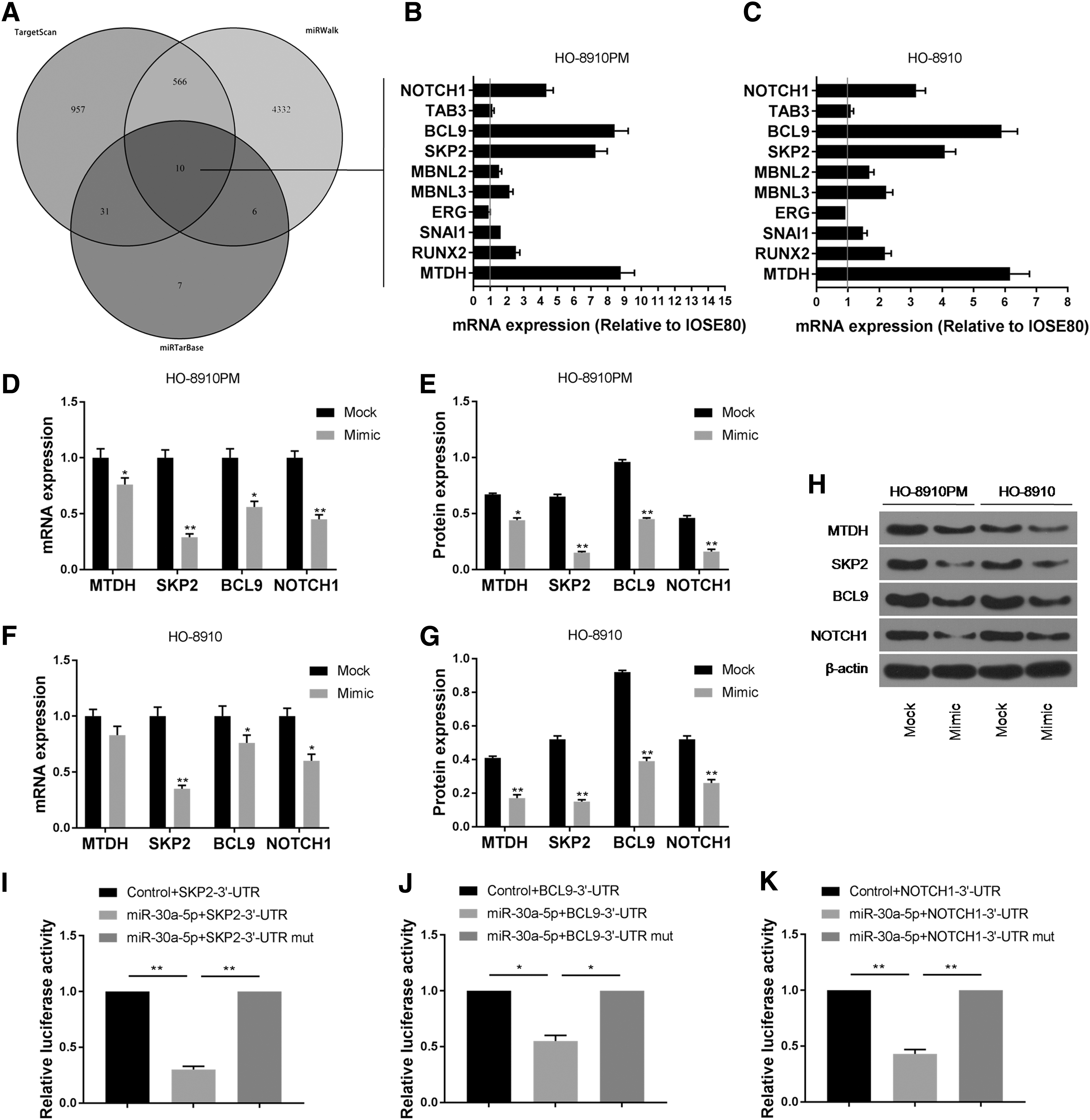
Prediction and validation of miR-30a-5p potential target genes.
To study the effects of miR-30a-5p on the expression of MTDH, SKP2, BCL9, and NOTCH1 genes, the mRNA and protein expression levels of MTDH, SKP2, BCL9, and NOTCH1 in HO-8910 and HO-8910PM cells after miR-30a-5p mimic plasmid transfection were examined by carrying out RT-qPCR and Western blotting. The results showed that the mRNA and protein expression levels of SKP2, BCL9, and NOTCH1 in miR-30a-5p mimic group were significantly lower than those in mock group (Fig. 3D–H, Supplementary Fig. S1). This indicated that the overexpression of miR-30a-5p inhibited the expression of SKP2, BCL9, and NOTCH1 genes, suggesting that SKP2, BCL9, and NOTCH1 might be the downstream targets of miR-30a-5p.
The dual-luciferase reporter assay showed that the relative luciferase activity was significantly decreased after being co-transfected with wild-type SKP2, BCL9, and NOTCH1 expression vector and miR-30a-5p, while the relative luciferase activity was not significantly changed after being co-transfected with mutant SKP2, BCL9, and NOTCH1 expression vector and miR-30a-5p (Fig. 3I–K). This demonstrated that SKP2, BCL9, and NOTCH1 are direct target genes of miR-30a-5p, and that miR-30a-5p directly regulates SKP2, BCL9, and NOTCH1 expression by 3′UTR.
Verification of miR-30a-5p by targeting SKP2, BCL9, NOTCH1 gene regulation in the migration and invasion of ovarian cancer cells
To study whether miR-30a-5p regulates ovarian cancer cell migration and invasion by targeting SKP2, BCL9, and NOTCH1 genes, HO-8910 and HO-8910PM cells were used to construct five experimental groups as follows: mock group, miR-30a-5p mimic group, miR-30a-5p mimic+SKP2 group, miR-30a-5p mimic+BCL9 group, and miR-30a-5p mimic+NOTCH1 group, and each group was detected and compared for its relative wound closure and relative migration rate. The results showed that the OD value of the co-transfected group was higher than that of the miR-30a-5p mimic group (Figs. 4A and 5A). For HO-8910PM cells, the relative wound closures in miR-30a-5p mimic+SKP2 group and miR-30a-5p mimic+NOTCH1 group were lower than that in miR-30a-5p mimic group. Our results also showed that the migration rate in miR-30a-5p mimic+SKP2 group was higher than that in miR-30a-5p mimic group (Fig. 4B–E). For HO-8910 cells, co-transfection of SKP2, BCL9, and NOTCH1 produced limited effect on relative wound closure. The relative migration rate in miR-30a-5p mimic+SKP2 group was found to be higher than that in miR-30a-5p mimic group (Fig. 5B–E). This suggests that the upregulation of SKP2 and NOTCH1 gene expression might relieve the inhibition of miR-30a-5p overexpression in ovarian cancer cell migration and invasion.
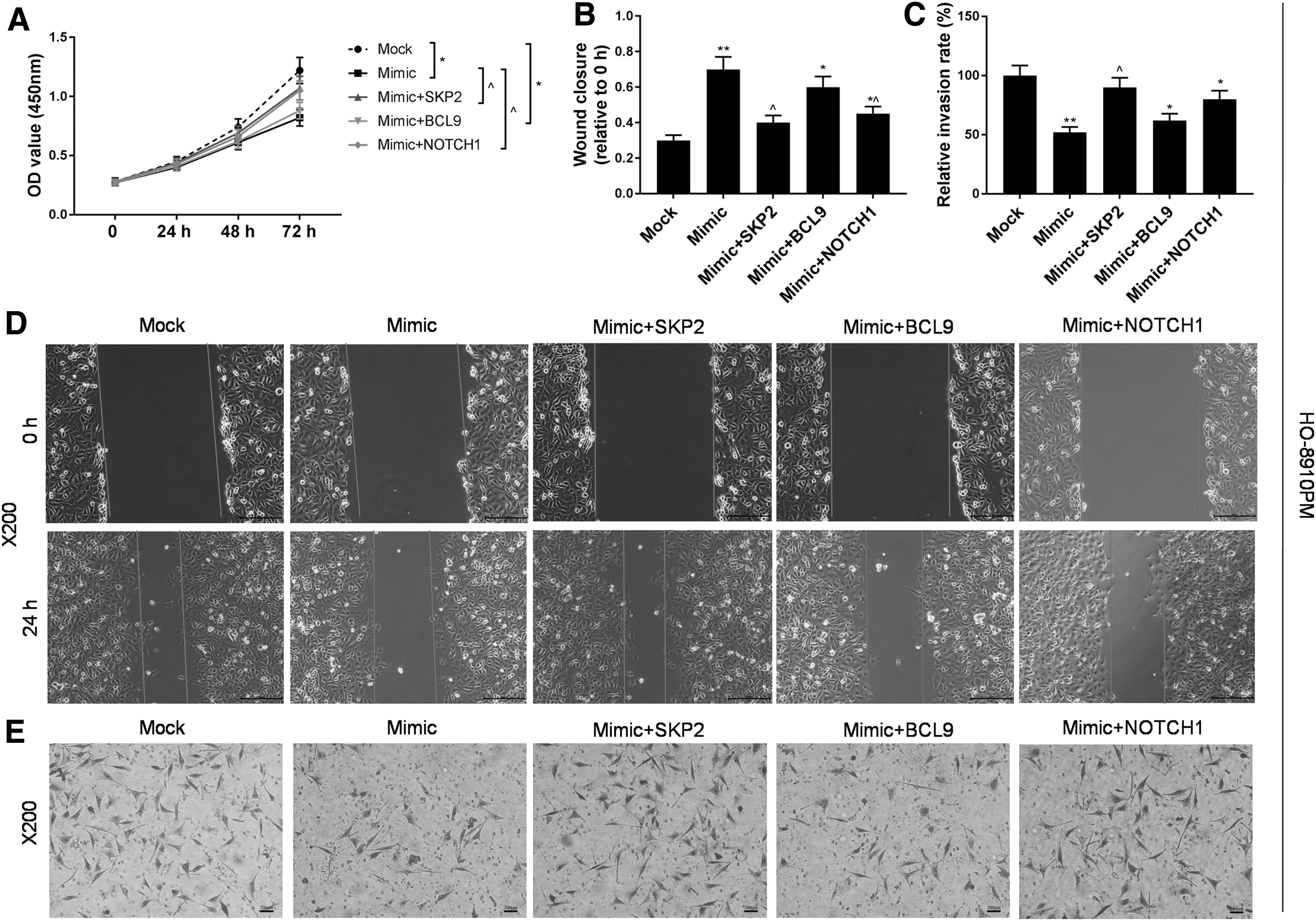
Effects of miR-30a-5p and downstream target genes on HO-8910PM cells.
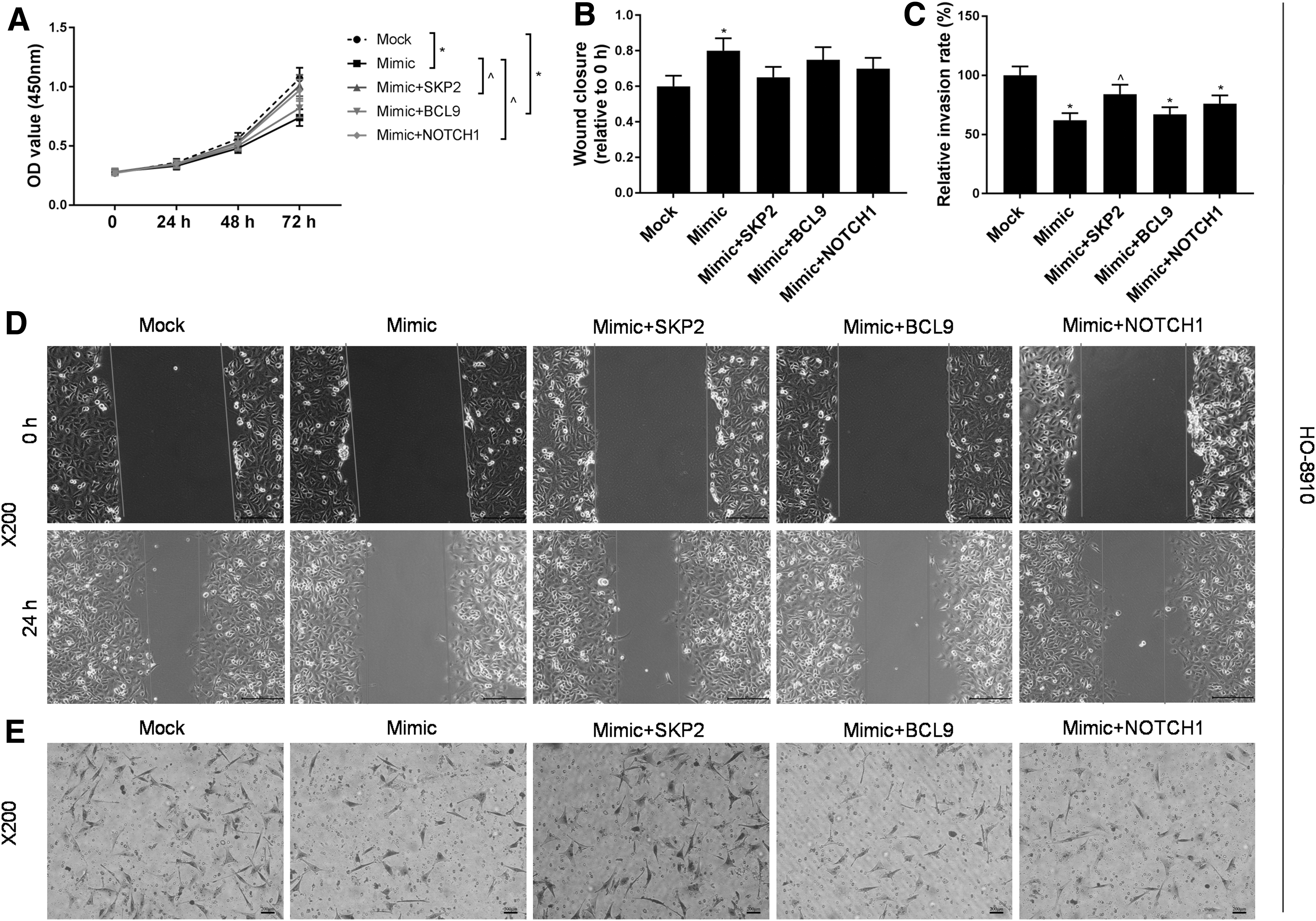
Effects of miR-30a-5p and downstream target genes on HO-8910 cells.
To further investigate the effects of miR-30a-5p and SKP2, BCL9, NOTCH1 gene expressions on the migration and invasion of ovarian cancer cells, EMT-related proteins and mRNA in the five groups were detected respectively using RT-qPCR and Western blotting. The results showed that E-cadherin in miR-30a-5p mimic group was significantly higher than that in mock group, while N-cadherin, Vimenin, and MPP-9 were significantly lower than those in mock group. E-cadherin and mRNA levels in miR-30a-5p mimic+SKP2 group, miR-30a-5p mimic+BCL9 group, and miR-30a-5p mimic+NOTCH1 group were lower than those in miR-30a-5p mimic group, while N-cadherin, Vimenin, and MPP-9 were significantly higher than those in miR-30a-5p mimic group (Fig. 6A–F, Supplementary Fig. S2 and Supplementary Fig. S3). These results indicated that the overexpression of miR-30a-5p inhibited the EMT process, while the upregulation of SKP2, BCL9, and NOTCH1 gene expression levels promoted the EMT process.

Effects of miR-30a-5p and downstream target genes on EMT-related gene expression of HO-8910PM and HO-8910 cells.
Discussion
Ovarian cancer is one of the most common malignancies in females. However, the pathogenesis of the cancer still remains unclear, and no specific treatment method has been found. Recently, a research has found that miRNAs play an important role in ovarian cancer. Although changes in miRNA expression levels have been observed in the development, progression, and metastasis of many types of malignant tumors, the regulation of tumor cell miRNAs and their molecular mechanisms still requires further research (Lu et al., 2005; Iorio et al., 2007; Tovar-Camargo et al., 2016). miRNA target prediction algorithms are a valuable tool for elucidating miRNA function. Current implementation of the program is realized mainly by identifying 3′UTR, and such a method still needs experimental verification (Thomson et al., 2011; Paraskevopoulou et al., 2013; Vlachos and Hatzigeorgiou, 2013). miRPath v3 is considered a more advanced method for the statistical analysis of miRNAs (Vlachos et al., 2015).
In this study, we discovered 25 miRNAs closely related to ovarian cancer. It was predicted by miRPath v3 that these miRNAs are closely related to mucin type O-glycan biosynthesis, in which miR-30a-5p has the highest correlation. It was also confirmed by preliminary RT-qPCR experiments that miR-30a-5p was lowly expressed in ovarian cancer cell lines. The mucin type O-glycan biosynthesis is involved in tumor migration and invasion (Yoshida et al., 2014; Kudelka et al., 2015; Ye et al., 2015). Therefore, this study mainly explored the relation between miR-30a-5p and ovarian cancer metastasis.
To investigate the role of miR-30a-5p in the regulation of migration and invasion of ovarian cancer cells, we used low metastatic cell line HO-8910 and high metastatic cell line HO-8910PM as target cells to construct miR-30a-5p-overexpressing cells by plasmid transfection. The results showed that HO-8910PM cells had higher migration and invasion ability compared with HO-8910 cells, and that the overexpression of miR-30a-5p inhibited cell migration and invasion. Studies have shown that by targeting IGF-1R, PETN, or DNMT1, a low expression of miR-30a-5p induced tumor cell resistance to chemotherapeutic drugs such as cisplatin. A feedback loop between miR-30a/c-5p and DNMT1 is involved in the mechanism of ovarian cancer being resistant to cisplatin, and such a mechanism is achieved by affecting the EMT process, and this feedback loop is considered a potential target for improving the prognosis of treatment (Han et al., 2017). miR-30a-5p could also inhibit the proliferation of human ovarian cancer cells by targeting forkhead box D1 (FOXD1) (Wang et al., 2018). Studies also found that a low expression of miR-30a-5p promotes the migration and invasion in human upper tract urothelial carcinoma, hepatocellular carcinoma, and breast cancer cells (Chen et al., 2016; Li et al., 2016a; Chung et al., 2017). However, little is known about the relation between miR-30a-5p and the ability of ovarian cancer cells to migrate and invade.
To explore the mechanism by which miR-30a-5p affects the migration and invasion of ovarian cancer cells, we predicted the possible target genes for miR-30a-5p via miRTarBase, Targetscan, and miRWalk. Among them, 10 common target genes for miR-30a-5p predicted in three databases were found as follows: MTDH, RUNX2, SNAI1, ERG, MBNL3, MBNL2, SKP2, BCL9, TAB3, and NOTCH1. The expression characteristics of these 10 genes in HO-8910 and HO-8910PM cell lines were examined. Our results showed that mRNA expression levels of MTDH, SKP2, BCL9, and NOTCH1 were significantly upregulated. The results also showed that miR-30a-5p overexpression significantly inhibited the expression of SKP2, BCL9, and NOTCH1 genes. Moreover, dual-luciferase reporter assay also confirmed that SKP2, BCL9, and NOTCH1 genes are the downstream target genes of miR-30a-5p.
To further investigate the mechanism by which miR-30a-5p regulates the migration and invasion of ovarian cancer by regulating SKP2, BCL9, and NOTCH1 gene expression, the effects of co-transfection on HO-8910 and HO-8910PM cell lines were respectively examined. Our results demonstrated that the transfection of SKP2, BCL9, and NOTCH1 overexpressing plasmids in different cell lines would alleviate the inhibitory effects of miR-30a-5p on cell migration and invasion. Among them, miR-30a-5p and four genes had stronger effects on the high metastatic cell line HO-8910PM. Further studies showed that the overexpression of miR-30a-5p inhibited the EMT process, while the upregulation of SKP2, BCL9, and NOTCH1 gene expression levels promoted the EMT process. In addition, according to previous studies, miR-30a-5p acted as a suppressor of several tumors (Wei et al., 2016; Li et al., 2017). Thus, we speculated that knocking down miR-30a-5p may promote cell growth and EMT of ovarian cancer.
The ability of cells to migrate and invade is related to EMT. In tumor cells, E-cadherin is often converted to Vimenin, causing the cells to lose their connection to the basement membrane and gain metastatic ability (Kalluri and Weinberg, 2009). The EMT process is regulated by a variety of genes and pathways. The overexpression of SKP2 is involved in the migration and invasion process in osteosarcoma cells, glioma cells, and oral squamous carcinoma cells (Wang et al., 2015; Yamada et al., 2016; Ding et al., 2017). miR-7 (Sanchez et al., 2013), miR-30 (Qi et al., 2015), and mi-340 (Fernandez et al., 2015) inhibit SKP2 expression at the post-transcriptional level. Studies have found that BCL9 promoted cell proliferation, migration, invasion, and metastasis of tumor cells via activated Wnt signaling pathways (Mani et al., 2009). A research has identified BCL9 as a direct target of miR-30 in some tumors (Jia et al., 2011; Zhao et al., 2014). Other evidence showed that Notch signaling increased transforming growth factor-β (TGF-β)/Smad signaling by upregulating the expression of TGF-β and TGF-β type-1 receptors in epithelial ovarian cancer cells, therefore promoting EMT (Gupta et al., 2013). It has been found that BCL9 also participates in pancreatic cancer metastasis by acting on TGF-β-induced EMT (Ling et al., 2016). Another study also found that miR-30a induces apoptosis of pancreatic beta cell apoptosis by targeting NOTCH1 pathway (Li et al., 2016b).
Conclusion
In summary, we investigated the potential role of miR-30a-5p in ovarian cancer metastasis and found that miR-30a-5p was lowly expressed in ovarian cancer and is closely related to ovarian cancer metastasis. SKP2, BCL9, and NOTCH1 are target genes downstream of miR-30a-5p, which might inhibit the migration and invasion of ovarian cancer cells by downregulating the expression of SKP2, BCL9, and NOTCH1 genes.
Footnotes
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.