
Abstract
The Qianhua Mutton Merino (QHMM) is a new variety of sheep (Ovis aries) with improved meat performance compared with the traditional Small Tail Han (STH) sheep variety. We recently reported the transcriptome profiling of longissimus muscle tissues between QHMM and STH sheep. In the present study, we aimed to evaluate key micro (mi)RNA–mRNA networks associated with sheep muscle growth and development. We used miRNA sequencing to obtain longissimus muscle miRNA profiles from QHMM and STH sheep. We identified a total of 153 known sheep miRNAs, of which 4 were differentially expressed (DE) between the 2 sheep varieties. We combined these results with mRNA library data to build an miRNA–mRNA network, including 26 target genes of the 4 DE miRNAs. Gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses showed that 26 target genes were significantly enriched in 86 biological processes, including muscle organogenesis, myoblast migration, cell proliferation, and adipose tissue development, and in 9 metabolic pathways, including carbohydrate, nucleotide, and amino acid metabolic pathways. oar-miR-655-3p and its target gene ACSM3 and oar-miR-381-5p and its target gene ABAT were selected for subsequent analysis based on GO and KEGG analyses. The binding sites of oar-miR-655-3p with ACSM3 and oar-miR-381-5p with ABAT were validated by a dual-luciferase reporter gene detection system. This represents the first integrative analysis of miRNA–mRNA networks in QHMM and STH muscles and suggests that DE miRNAs, especially oar-miR-655-3p and oar-miR-381-5p, play crucial roles in muscle growth and development.
Introduction
High-quality meat performance of domestic sheep breed is a valuable genetic resource for the global sheep industry. The Qianhua Mutton Merino (tentative name; abbreviated as QHMM) is a new sheep variety bred for both meat and wool. It was bred through graded crossing and artificial selection of the South African Mutton Merino (male parent) and Chinese Northeast Fine-Wool sheep (female parent) in recent years. It has a good body shape, with characteristics of no angle, a wide deep chest, straight back, and well-developed hindquarters (Fig. 1a).

QHMM and STH sheep.
This new breed demonstrates strong stress and roughage resistance and better meat performance compared with both of its parents. We previously investigated the transcriptome profile of muscle tissues of QHMM and Small Tail Han (STH) sheep, which is a local Chinese breed. This revealed phenotypic differences between the two breeds and identified some differentially expressed genes (DEGs) and pathways that play crucial roles in muscle growth and development (Sun et al., 2016).
Micro (mi)RNAs are endogenously expressed, small noncoding RNAs that are evolutionarily conserved and regulate gene expression in a variety of biological activities (Carrington and Ambros, 2003). For example, the growth and development of skeletal muscle is a complex process involving miRNAs such as miRNA-1 (Chen et al., 2006, 2010), miRNA-133 (Chen et al., 2006, 2009; Boutz et al., 2007; Huang et al., 2011), and miRNA-206 (Anderson et al., 2006; Kim et al., 2006; Rosenberg et al., 2006; Chen et al., 2010; Dey et al., 2011).
miRNA-regulated gene expression affects target mRNAs, leading to mRNA cleavage or translational repression (Chen et al., 2004; Rajewsky, 2006). Although several methods have been used to predict miRNA targets, including TargetScan, miRanda, and PicTar (Krek et al., 2005; Rajewsky, 2006; Betel et al., 2008), the specificity and sensitivity of these approaches are low. The expression of miRNAs and their mRNA targets is widely accepted to correlate (Selbach et al., 2008; Djuranovic et al., 2012), and in recent years, the integrative analysis of miRNAs–target mRNAs has been increasingly used to identify potential key interactions between miRNAs and mRNAs (Huang et al., 2007; Tian et al., 2008; Gennarino et al., 2009). However, the miRNA–mRNA regulatory network associated with sheep muscle growth and development has rarely been investigated.
In this study, we integrated the miRNA–mRNA paired expression profiling of longissimus muscle tissues from QHMM and STH sheep to identify miRNAs that were differentially expressed (DE) between the two breeds. Combined with the findings from our previous study (Sun et al., 2016), we integrated the miRNA–target mRNA regulation information to construct an miRNA–mRNA regulatory network associated with sheep muscle growth and development. The DE miRNAs and target genes identified in this study will offer insights into the molecular mechanism underlying sheep muscle development and provide a basis for mutton sheep breeding.
Materials and Methods
Animal sample preparation
QHMM and STH sheep were obtained from Jilin Qian'an Zhihua Sheep Breeding Co., Ltd., (Qian'an, China). All methods were carried out in accordance with relevant guidelines established by the Ministry of Agriculture of the People's Republic of China. All experimental protocols were approved by the Jilin Laboratory Animal Specialized Committee. All experimental sheep were raised under the same environment with natural light and free access to food and water. Three adult females (aged 1 year) were randomly selected from each breed and sacrificed to obtain longissimus dorsi muscle samples. All samples were immediately snap-frozen in liquid nitrogen for total RNA extraction.
Construction of mRNA and small RNA library and sequencing
Total RNA was extracted from muscle tissue samples using TRIzol reagent (Life Technologies). The RNA quality was checked using a NanoDrop 2000 spectrophotometer (Thermo Scientific) and Agilent 2200 TapeStation system (Agilent). Construction of an mRNA library and sequencing were performed as previously described (Sun et al., 2016). miRNA was purified using an miRNeasy Mini Kit (Qiagen, Germany). cDNA libraries were prepared for single-end sequencing using the Ion Total RNA-Seq (RNA sequencing) Kit, v2.0 (Life Technologies), and were size-selected by polyacrylamide gel electrophoresis for miRNA sequencing.
Samples were diluted and processed on an Ion OneTouch 2 instrument (Life Technologies) and enriched on an Ion OneTouch 2 ES station (Life Technologies) to prepare template-positive Ion PI™ Ion Sphere™ Particles (Life Technologies) for the Ion PI Template OT2 200 Kit, v2.0 (Life Technologies). After enrichment, template-positive Ion PI Ion Sphere Particles were loaded onto a P1v2 Proton Chip (Life Technologies) and sequenced on proton sequencers using the Ion PI Sequencing 200 Kit, v2.0 (Life Technologies), by NovelBio Bio-Pharm Technology Co., Ltd. (Shanghai, China).
RNA-seq data analysis
mRNA sequencing data analysis was performed as previously described (Sun et al., 2016). For miRNA sequencing data analysis, raw miRNA-seq reads were filtered and clean reads were mapped to the sheep (Ovis aries) mature miRNA database in miRBase (v21) and also to the Rfam database (
As no DE miRNAs were selected by the BH algorithm using FDR <0.05, we identified DE miRNAs between QHMM and STH sheep by the DESeq algorithm using the following parameters: fold change >1.5 or fold change <0.667 and p-value <0.05. According to the information of known sheep DE miRNAs and novel DE miRNAs, the volcano plot and heat map were obtained by using R language and Cluster 3.0 software, respectively.
Target gene analysis and mRNA–miRNA analysis
We used the miRanda tool to predict target genes of DE miRNAs. These target genes were then compared with transcriptome profiling data (Sun et al., 2016), and Pearson correlation coefficients were computed between DE miRNAs and DE mRNAs. Only DEGs whose expression negatively correlated with target miRNA were selected as putative miRNA targets. miRNA–mRNA interaction networks were visualized using Cytoscape software (Paul et al., 2003) by their differential expression values and according to miRNA–gene interactions in the Sanger miRBase database.
Gene ontology and Kyoto Encyclopedia of Genes and Genomes pathway analyses
Gene ontology (GO) analysis (Ashburner et al., 2000) (
Validation of RNA-seq data by quantitative reverse transcription-PCR
mRNA sequencing data were validated as previously described (Sun et al., 2016). For validation of miRNA sequencing data, four known sheep DE miRNAs (oar-miR-758-3p, oar-miR-655-3p, oar-miR-381-3p, and oar-miR-381-5p) and four novel DE miRNAs (chr20-37401-mature, chr10-23287-mature, chr15-30212-mature, and chr7-17775-mature) were selected.
Total miRNA was extracted from each muscle sample using the miRcute miRNA isolation kit (Tiangen, Beijing, China). cDNA was then synthesized using stem–loop (Chen et al., 2005; Shi and Chiang, 2005) primers (Supplementary Table S1) and a ReverTra Ace qPCR RT Kit (FSQ-101; Toyobo, Japan) from 1 μg of total miRNA samples. The U6 gene was used as a reference housekeeping gene.
qPCR reactions were performed using the Bio-Rad CFX96 system with 20-μL reactions comprising 10 μL of SYBR Green Real-time PCR Master Mix (QPK-201; Toyobo), 0.8 μL each of the forward and reverse primers (200 μM) (Supplementary Data), 2 μL of cDNA, and 6.4 μL of distilled water. The quantitative reverse transcription (qRT)-PCR program was 95°C for 60 s, followed by 40 cycles at 95°C for 15 s, 64°C for 15 s, and 72°C for 45 s, with a final stage of melting curve analysis. Pearson correlation coefficients between RT-qPCR and RNA-seq data of eight DE miRNAs were calculated using SPSS 21.0 software.
Cell culture
HEK-293T cells were cultured in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal bovine serum, 1.5 mM
Plasmid vector construction and transfection
The sequence fragment of ACSM3-3′UTR that binds oar-miR-655-3p (wild-type), ACSM3-3′UTR with an oar-miR-655-3p binding site mutation (mutant), and a positive control (oar-miR-655-3p inhibitor) and the sequence fragment of ABAT-3′UTR that binds oar-miR-381-5p (wild-type), ABAT-3′UTR with an oar-miR-381-5p binding site mutation (mutant), and a positive control (oar-miR-381-5p inhibitor) were designed by TargetScan 7.1 and RNA22, v2.0, target prediction algorithms and synthesized by total gene synthesis by Sangon Biotech Company.
Sequence information of wild-type and mutant target gene fragments and the miRNA inhibitor fragment is shown in Table 1. The pmirGLO dual-luciferase miRNA target expression vector (E1330; Promega Corporation, Madison) was used to confirm the putative binding sites of ACSM3-3′UTR with oar-miR-655-3p and ABAT-3′UTR with oar-miR-381-5p. pmirGLO was digested by SacI and XhoI and ligated to target gene fragments by T4 DNA ligase.
Sequence Information of Wild-Type and Mutant-Type Target Gene Fragments
SacI (GAGCTC) and XhoI (CTCGAG) restriction sites were added to the 5′ and 3′ ends, respectively, of the target fragment.
Before plasmid vector transfection, the cell transfection efficiency of 293T cells was evaluated by three gradients of Lipofectamine 2000 transfection reagent (Life Technologies) (0.3/0.6/1 μL) and recombinant plasmid vector (0.2/0.4/0.8 μg).
293T cells were washed with D-Hank's solution and incubated with DMEM containing 10% fetal bovine serum for 24 h. Then, 2 μL of miRNA or miRNA inhibitor (20 μM) was mixed with 100 μL of Opti-MEM I and 0.8 μg of recombinant plasmid vector, and 3 μL of Lipofectamine 2000 transfection reagent was mixed with 100 μL of Opti-MEM I. The two mixtures were combined and incubated for 20 min at room temperature, then added to cells and incubated at 37°C for 6 h. Subsequently, 500 μL of DMEM containing 10% fetal bovine serum was added to the cells and incubated in 5% CO2 at 37°C. Cells were collected after culturing for 24 and 48 h.
Seven experimental groups for ACSM3 with oar-miR-655-3p as well as ABAT with oar-miR-381-5p were established: ACSM3-WT+mimic NC, ACSM3-WT+oar-miR-655-3p, ACSM3-MUT+mimic NC, ACSM3-MUT+oar-miR-655-3p, PC (oar-miR-655-3p inhibitor)+mimic NC, PC (oar-miR-655-3p inhibitor)+oar-miR-655-3p, and a control group; and ABAT-WT+mimic NC, ABAT-WT+oar-miR-381-5p, ABAT-MUT+mimic NC, ABAT-MUT+oar-miR-381-5p, PC (oar-miR-381-5p inhibitor)+mimic NC, PC (oar-miR-381-5p inhibitor)+oar-miR-381-5p, and a control group. There were three replicates of each group.
Dual-luciferase reporter detection
Luciferase and Renilla activities were measured using a Dual-Luciferase Reporter Assay Kit (Promega Corporation) after transfection for 24 and 48 h.
Statistical analysis
Data were analyzed using SPSS 21.0 software, and differences in significance between groups were assessed by analysis of variance. Differences were considered statistically significant when p < 0.05.
Results
Summary of RNA-seq data
We obtained a total of 14.01, 12.90, 11.21, 12.34, 13.47, and 15.76 million clean reads for QHMM sheep A1, A2, and A3 and STH sheep B1, B2, and B3, respectively. These clean reads were mapped to the sheep mature miRNA database in miRBase (v21) and the Rfam database, as shown in Table 2 and Supplementary Figure S1. Because the sheep mature miRNA sequence is incomplete, the unique mapped rate ranged from 0.166 to 0.226.
Statistical Analysis of Clean Read Alignment to the Sheep miRNA Database
A total of 153 known sheep miRNAs were identified, including 152 that were expressed in both QHMM and STH libraries and 1 (oar-miR-410-5p) that was only expressed in the STH library (Supplementary Table S2). The unmapped clean reads were then mapped to the Oar 3.1 version of the sheep genome sequence, and the distribution of mapped reads on chromosomes is shown in Supplementary Figure S2. A significant difference was observed in the chromosomal distribution of QHMM and STH mapped reads. A total of 8076 novel miRNAs were predicted.
DE miRNAs
The DE-seq program was used to identify DE miRNAs in the muscle tissues of STH versus QHMM sheep. As shown in Table 3, only four known sheep DE miRNAs were identified, including three upregulated miRNAs and one downregulated miRNA, because the sheep mature miRNA sequence is not completed. A total of 146 novel DE miRNAs were identified, including 91 that were upregulated and 55 that were downregulated.
Known miRNAs with Differential Expression Between Qianhua Mutton Merino and Small Tail Han Sheep
QHMM, Qianhua Mutton Merino; STH, Small Tail Han.
The volcano plot and heat map of DE miRNAs are shown in Figure 2; as shown on the volcano plot, the total number of DE miRNAs in the STH versus QHMM group comparison revealed 150 DE miRNAs (94 up, 56 down). The heat map showed the DE miRNA patterns in three samples from STH and three samples from QHMM sheep, and clusterization confirmed the presence of two superior clusters of the size corresponding to the number of up- and downregulated miRNAs.

The volcano plot and heat map of miRNAs.
Target gene analysis and integrative mRNA–miRNA analysis
The miRanda database was used to predict target genes of the 4 known and 146 novel sheep DE miRNAs. A total of 760 target genes (Supplementary Table S3) of the known sheep DE miRNAs and 58,349 target genes of the novel DE miRNAs were predicted. Predicted target genes were combined with transcriptome profiling data (Sun et al., 2016) to construct an mRNA–miRNA network associated with sheep muscle growth and development. Significant mRNA–miRNA pairs that exhibited reverse expression patterns between QHMM and STH muscles were selected to identify a total of 26 target genes (Supplementary Table S4) of the 4 known sheep DE miRNAs and 2069 target genes of the novel DE miRNAs. The mRNA–miRNA network is shown in Figure 3.
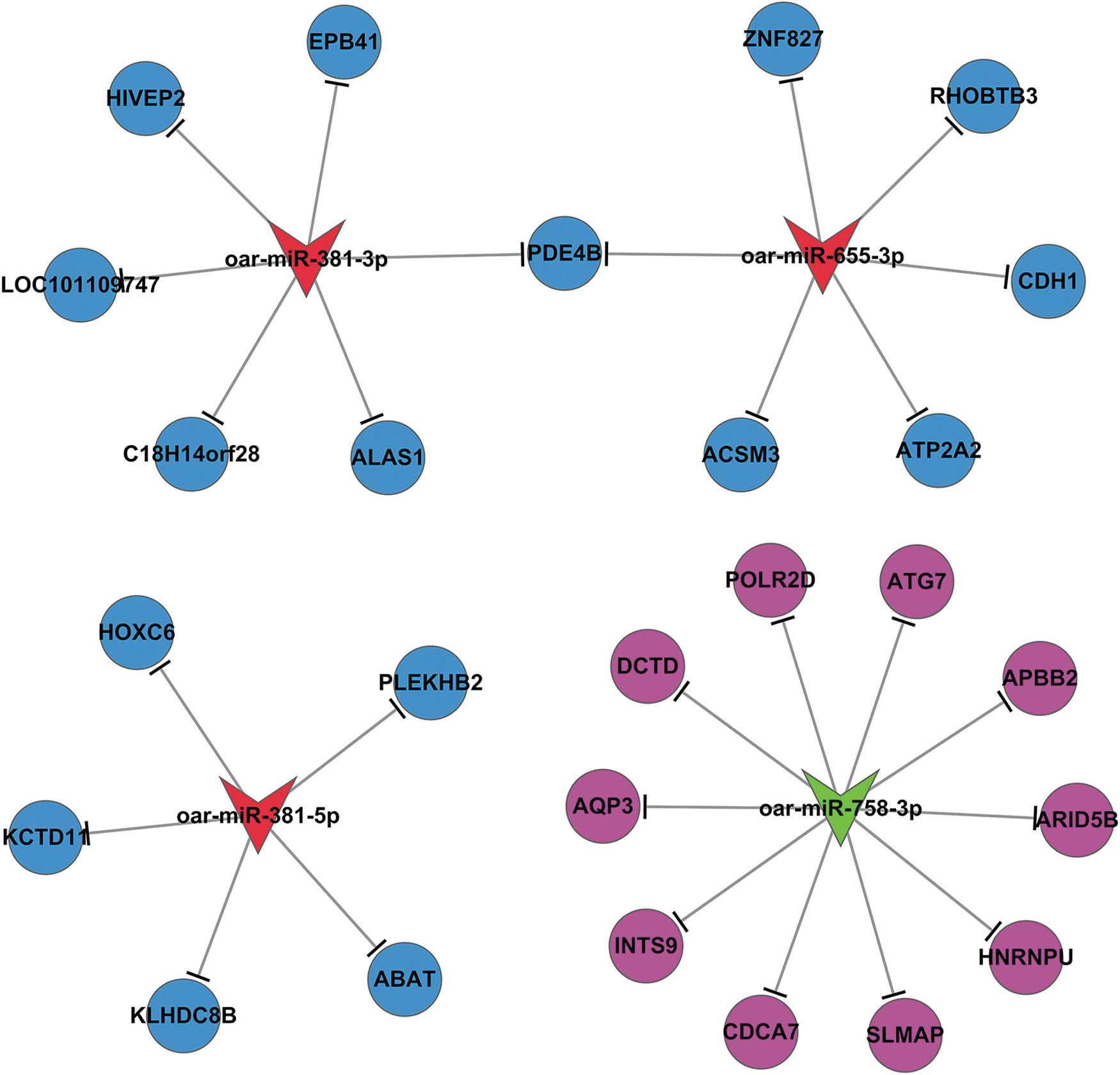
The miRNA–mRNA network in muscle tissues of QHMM and STH sheep.
GO and Kyoto Encyclopedia of Genes and Genomes pathway analyses of target DEGs
To classify the related functions of these miRNA-regulated genes, GO analysis was performed. The 26 target DEGs were annotated according to 3 GO categories: biological process, molecular function, and cellular component. Significance analysis revealed significant enrichment (p < 0.05) of 86 GO terms (including the gamma-aminobutyric acid catabolic process, positive regulation of immune system process, and regulation of cell proliferation), 19 GO terms (including the mitochondrial matrix, spectrin-associated cytoskeleton, and smooth endoplasmic reticulum), and 29 GO terms (including catalytic activity, fatty acid ligase activity, and 4-aminobutyrate transaminase activity) for the 26 target DEGs in the categories of biological process, molecular function, and cellular component, respectively (Supplementary Table S5). The top 15 significant GO terms among the three categories are shown in Figure 4a–c.
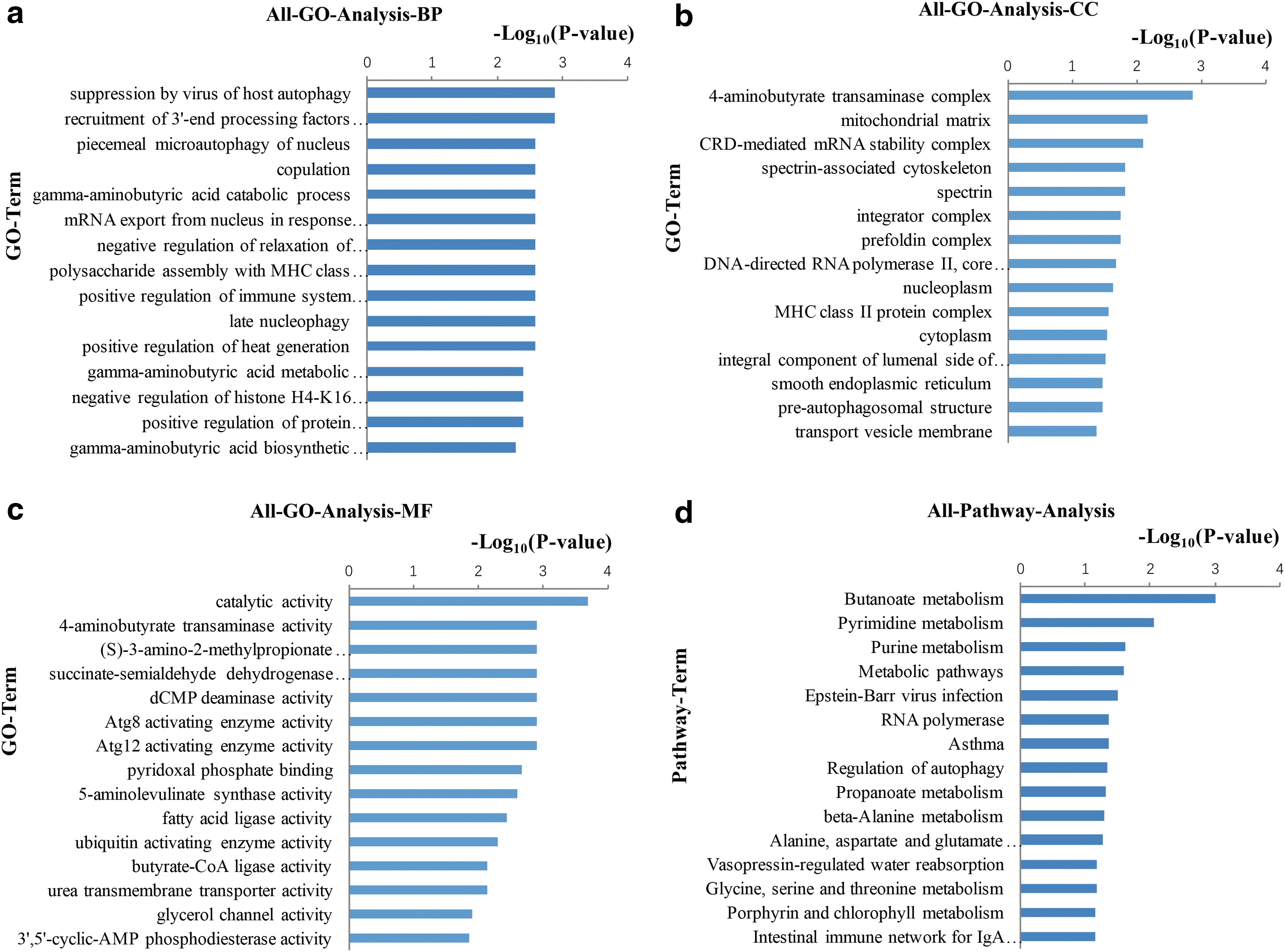
The top 15 significant GO terms and pathways of DE genes (STH vs. QHMM; p < 0.05).
Pathway annotation of DEGs was performed using the Kyoto Encyclopedia of Genes and Genomes (KEGG) database. Nine pathways (including butanoate metabolism, pyrimidine metabolism, and purine metabolism) were significantly (p < 0.05) enriched among the 26 target DEGs (Supplementary Table S6). The top 15 significant pathways are shown in Figure 4d.
After construction of the mRNA–miRNA network and GO and KEGG analyses, we found that ACSM3 is associated with biological processes, including the fatty acid metabolic process, fatty acid biosynthetic process, and lipid metabolic process, as well as the carbohydrate metabolism pathway, including butanoate metabolism. ABAT is associated with biological processes, including positive regulation of insulin secretion, and the carbohydrate metabolism pathway, including butanoate metabolism, and the amino acid metabolism pathway, including valine, leucine, and isoleucine degradation and alanine, aspartate, and glutamate metabolism. GO and pathway terms related to ACSM3 and ABAT are shown in Table 4. We selected ACSM3 as a target gene of oar-miR-655-3p and ABAT as a target gene of oar-miR-381-5p for a follow-up study.
Gene Ontology Terms and Pathway Analysis Terms of ACSM3 and ABAT
SacI (GAGCTC) and XhoI (CTCGAG) restriction sites were added to the 5′ and 3′ ends, respectively, of the target fragment.
GO, gene ontology.
Validation of miRNA-seq data
mRNA sequencing data validation results were reported previously (Sun et al., 2016). Eight DEGs were randomly selected to validate miRNA sequencing data by qRT-PCR. The 2–ΔΔCt method (Livak and Schmittgen, 2001) was used to calculate relative quantification, as shown in Figure 5a and b. The known sheep DE miRNAs, oar-miR-655-3p, oar-miR-381-3p, and oar-miR-381-5p, were significantly upregulated in STH sheep compared with QHMM sheep (p < 0.05), while oar-miR-758-3p was significantly downregulated (p < 0.05) in STH sheep. The novel DE miRNAs, chr10-23287-mature and chr15-30212-mature, were significantly upregulated (p < 0.05), while chr20-37401-mature and chr7-17775-mature were significantly downregulated (p < 0.01 and p < 0.05, respectively) in STH sheep.
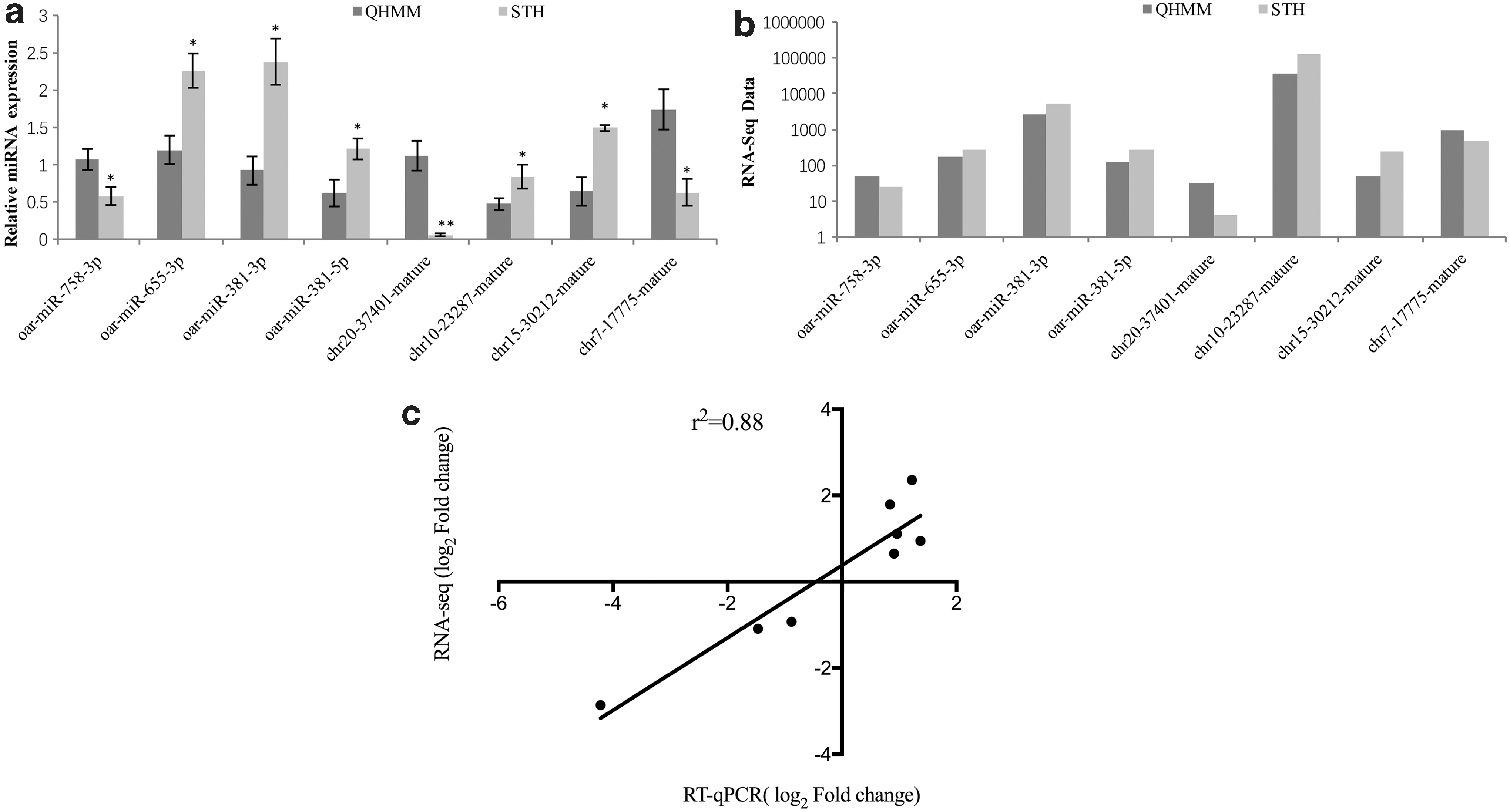
Real-time PCR validation of miRNAs DE between QHMM and STH sheep.
A Pearson correlation coefficient of 0.88 is observed between RT-qPCR and RNA-seq data of eight DE miRNAs, as shown in Figure 5c. The expression levels of these miRNAs determined by qRT-PCR were consistent with miRNA-Seq data, which validated their accuracy.
oar-miR-655-3p downregulates ACSM3 expression and oar-miR-381-5p downregulates ABAT expression
Based on GO and pathway analysis, oar-miR-655-3p and ACSM3 and oar-miR-381-5p and ABAT were selected to verify their targeted regulatory relationship. The binding sites of oar-miR-655-3p with ACSM3 and oar-miR-381-5p with ABAT are shown in Figure 6a and b, and the pmirGLO plasmid vector and its working principle are shown in Figure 6c and d. The pmirGLO plasmid vector achieved a transfection efficiency of more than 90%, even with low concentrations of transfection reagents and plasmids, suggesting that it could be used for follow-up tests (Supplementary Fig. S3). Luciferase assays confirmed the potential binding between oar-miR-655-3p and ACSM3 and oar-miR-381-5p and ABAT to further validate the miRNA–mRNA interactions.

Binding sites of miRNAs with target genes and the pmirGLO plasmid vector.
In the case of ACSM3 pmirGLO wild-type, the luciferase activity of the oar-miR-655-3p mimic group was significantly decreased (p < 0.05) compared with oar-miR-655-3p-NC, while there was no significant difference in luciferase activity between oar-miR-655-3p mimic and oar-miR-655-3p-NC groups (p > 0.05) with respect to the ACSM3 pmirGLO mutant type at 24 h and 48 h (Fig. 7a, b). In the case of ABAT pmirGLO wild-type, the luciferase activity of the oar-miR-381-5p mimic group was significantly decreased (p < 0.05) compared with oar-miR-381-5p-NC, while there was no significant difference in luciferase activity between oar-miR-381-5p mimic and oar-miR-381-5p-NC groups (p > 0.05) with respect to the ABAT pmirGLO mutant type at 24 h and 48 h (Fig. 7c, d).

Detection of interactions between oar-miR-655-3p and ACSM3 and between oar-miR-381-5p and ABAT by the dual-luciferase reporter system.
These results indicated that oar-miR-655-3p directly targets ACSM3-3′UTR to downregulate ACSM3 expression and that oar-miR-381-5p directly targets ABAT-3′UTR to downregulate ABAT expression.
Discussion
Various regulatory factors control key gene expression, including transcriptional factors, epigenetic factors, and post-transcriptional factors (Zhang et al., 2015). miRNAs play an important regulatory role, mainly through the post-transcriptional binding of target gene 3′ untranslated region seed sequences, which degrades or inhibits the expression of mRNAs. Several studies have shown that ∼30% of animal genes are regulated by miRNAs (Lewis et al., 2005; Friedman et al., 2009), so identification of target mRNAs is essential to understand miRNA biological functions.
However, validation of target genes is difficult and time-consuming because of the high false-positive rate of miRNA prediction programs (Lewis et al., 2005; Sethupathy et al., 2006; Peterson et al., 2014). In recent years, microarray-based techniques have been used to identify mRNA–miRNA interactions by determining negative expression correlations between miRNAs and target mRNAs (Tang et al., 2015; Mach et al., 2016; Xu et al., 2016). In the present study, we identified potential core mRNA–miRNA interactions that occur during muscle growth and development in two breeds of sheep. This provides information to further characterize mRNA–miRNA interactions in sheep muscle growth and development.
We previously analyzed mRNA expression profiles of QHMM and STH sheep using high-throughput RNA technology (Sun et al., 2016). In the present study, we combined these earlier findings with current miRNA expression profiles to create an integrative miRNA–mRNA analysis of muscle tissues from QHMM and STH sheep. We identified 4 DE miRNAs and 26 target genes using miRNA-seq and bioinformatic analysis.
Previous studies identified 16 DE miRNAs and 98 target genes by integrative miRNA–mRNA analysis between Dorper and STH sheep (Miao et al., 2015), while skeletal muscle development has been shown to be regulated by many miRNAs, including miRNA-1 (Chen et al., 2006, 2010), miRNA-23a (Wang et al., 2012), miRNA-24 (Sun et al., 2008), miRNA-26a (Wong and Tellam, 2008; Dey et al., 2012), miRNA-27b (Crist et al., 2009), miRNA-29 (Wei et al., 2013), miRNA-125b (Ge et al., 2011), miRNA-133 (Chen et al., 2006, 2009; Boutz et al., 2007; Huang et al., 2011), miRNA-148a (Zhang et al., 2012), miRNA-155 (Seok et al., 2011), miRNA-181 (Naguibneva et al., 2006), miRNA-206 (Anderson et al., 2006; Kim et al., 2006; Rosenberg et al., 2006; Chen et al., 2010; Dey et al., 2011), miRNA-208a/b (van Rooij et al., 2009), miRNA-221/222 (Cardinali et al., 2009), miRNA-214 (Juan et al., 2009; Liu et al., 2010), miRNA-322/424/50 (Sarkar et al., 2010), miRNA-486 (Dey et al., 2011), and miRNA-499 (Naguibneva et al., 2006; Wang et al., 2011). The function of these miRNAs is mainly associated with proliferation and differentiation of skeletal muscle cells and regulation of skeletal muscle myogenesis.
However, the identity of many miRNAs involved in mammalian muscle development remains unknown. We identified a total of 153 known miRNAs in QHMM and STH sheep, of which 4 were DE between QHMM and STH sheep muscle tissues. These DE miRNAs have not been reported to date, so our findings will enrich what is known about miRNAs associated with muscle growth and development. Construction of an mRNA–miRNA network using the integrative method indicated that these 4 DE miRNAs may play an essential role in regulation of sheep muscle growth and development through their interaction with 26 target genes. These results generated accurate data for sheep miRNA research.
To further investigate how these DE miRNAs regulate muscle growth and development, GO and KEGG analyses were performed for the 26 target genes.
GO analysis showed that the genes were significantly enriched in gamma-aminobutyric acid biosynthetic and gamma-aminobutyric acid catabolic processes. Gamma-aminobutyric acid was previously shown to improve the meat performance of pigs and increase food intake, eye muscle area, and the slaughter rate of pigs (Fan et al., 2007). The target genes were also significantly enriched in peptide antigen assembly and polysaccharide assembly with the major histocompatibility complex (MHC) class II protein complex. MHC is an important component of myosin that functions in muscle growth and contraction (Zhang et al., 2009). We identified significantly enriched GO terms directly associated with muscle growth and development, including muscle organ morphogenesis, fibroblast migration, cardiac muscle cell development, regulation of cell proliferation, and adipose tissue development.
This suggested that DE miRNAs regulate growth and development of sheep muscles by controlling these biological functions of target genes.
KEGG analysis was similarly performed, which showed that the target genes were significantly enriched in carbohydrate metabolism pathways, including butanoate and propanoate metabolism, and nucleotide metabolic pathways, including pyrimidine and purine metabolism. Previous research found that beta-hydroxy-beta-methylbutyrate has a regulated effect on muscle tissue protein synthesis, muscle fiber diameter, and early muscle development (Soares et al., 2001; Moore et al., 2005; Kovarik et al., 2010). The target genes were also enriched in amino acid metabolism pathways, including alanine, aspartate, and glutamate metabolism and glycine, serine, and threonine metabolism. Therefore, we predicted that DE miRNAs regulate sheep meat performance using these pathways associated with metabolism and accumulation of nutrients.
The miRNA–mRNA network obtained in our study revealed the regulatory relationship between miRNAs and target genes after transcription. Taking all our results into account, we propose that the miRNAs, oar-miR-655-3p and oar-miR-381-5p, play critical roles in sheep meat production performance and meat quality by regulating biological processes such as fatty acid metabolic and biosynthetic processes, lipid metabolic processes, and the carbohydrate metabolism pathway through their target genes, ACSM3 and ABAT.
Recently, the dual-luciferase reporter gene method was shown to be the most convenient for verifying the relationship between miRNAs and their target mRNAs. We used this method to show that oar-miR-655-3p downregulates ACSM3 expression and oar-miR-381-5p downregulates ABAT expression. miR-655 was previously reported to be a key miRNA in human disease, with a role in regulating cancer progression through the transforming growth factor-beta signaling pathway by targeting ZEB1 and TGFBR2 (Harazono et al., 2013); miR-655 also regulates breast cancer by regulating Prrx1 (Lv et al., 2016).
Thus far, little is known about miR-655 in ruminant muscle tissue. In this study, we showed for the first time that ACSM3 is the target gene of oar-miR-655-3p. ACSM3 is a member of the acyl-CoA synthetase medium-chain family (ACSM), which is involved in fatty acid-related biological activities (Watkins et al., 2007). ACSM3 was first identified in kidneys of hypertensive rats in 1991 (Iwai and Inagami, 1991), and most research on this gene has focused on human diseases (De Preter et al., 2012). Previous studies reported that the ACSM family is involved in human hypertension (Iwai t al., 2002), obesity (N et al., 2002; Benjafield et al., 2003), and diabetes (Lindner et al., 2006), but no research has reported on its role in growth and development of animal muscles.
We combined GO and pathway analyses to reveal that ACSM3 is involved in biological processes such as fatty acid biosynthesis and metabolism, lipid metabolism, and the butanoate metabolism pathway.
miR-381 also plays an essential role in human disease (Zhou et al., 2015; Xue et al., 2016) by inhibiting the invasion of renal cell carcinoma by regulating the expression of CBP, LEF-1, and the β-catenin gene (Chen and Liu, 2015). It also regulates the excessive growth of chondrocytes by targeting HDAC4 (Zhang et al., 2016). However, no studies have reported its role in ruminant muscle tissue. In this study, we verified for the first time that ABAT is the target gene of oar-miR-381-5p. ABAT encodes the key enzyme γ-amino butyric acid transaminase, which is involved in decomposition of γ-amino butyric acid.
Research into ABAT has mainly focused on human diseases such as nervous system diseases (Barnby et al., 2005), digestive system diseases (Jirholt et al., 2011), and tumors (Jansen et al., 2015), but no studies have reported a role in growth and development of animal muscles. Our GO and pathway analyses revealed that ABAT is associated with biological processes, including positive regulation of insulin secretion, the carbohydrate metabolism pathway, including butanoate metabolism, and the amino acid metabolism pathway, including valine, leucine, and isoleucine degradation and alanine, aspartate, and glutamate metabolism.
Therefore, we can infer that oar-miR-655-3p decreases ACSM3 expression and oar-miR-381-5p decreases ABAT expression to regulate sheep muscle growth and development, although specific biological functions and detailed regulatory mechanisms require further study.
In this study, we established miRNA profiles of longissimus muscles in QHMM and STH sheep, which have different meat performances. We identified key candidate miRNAs and a core functional miRNA–mRNA network associated with sheep muscle growth and development. We also selected oar-miR-655-3p and its target gene ACSM3 and oar-miR-381-5p and its target gene ABAT for subsequent analysis. The integrative miRNA–mRNA analysis of QHMM and STH muscles is reported for the first time and suggests a role for miRNA–mRNA interactions in promoting sheep muscle growth and development. Our study is also the first to reveal that ACSM3 is targeted by oar-miR-655-3p and ABAT is targeted by oar-miR-381-5p in this function. Further investigation of the selected miRNA–mRNA pairs will provide novel insights into molecular mechanisms of sheep muscle growth and development.
Footnotes
Acknowledgments
The authors thank the Jilin Qian'an Zhihua Sheep Breeding Co., Ltd. (Qian'an, China), for providing the experimental animals and NovelBio Bio-Pharm Technology Co., Ltd. (Shanghai, China), for help with RNA sequencing. This study was supported by the Jilin Province Livestock and Poultry Breeding Special Fund (No. 2015062), the Jilin Province Modern Agricultural Industry Technology System Special Fund (No. 201541), the Jilin Province Science and Technology Development Project (No. 20110237), and the Jilin Provincial Department of Education “12th Five-Year” Science and Technology Development Project (No. 2015186).
Availability of Data and Materials
Raw sequence and processed data have been submitted to the Gene Expression Omnibus database (GEO dataset;
Authors' Contributions
L.M.S. and H.Z.J. designed the project; L.M.S. conducted all experiments and wrote the manuscript; S.Y.L., M.B., L.J.X., J.R.L., and C.J. prepared animal samples. All authors reviewed the manuscript.
Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.