
Abstract
MicroRNAs (miRNAs) can regulate a variety of biological functions such as fat and cholesterol synthesis, translocation, and utilization by regulating degradation or by inhibiting target mRNA translation. In this study, the target gene of miR-21-3p was analyzed through bioinformatics, which predicted Elovl5. Furthermore, miR-21-3p was verified for regulation of triglyceride expression. The results showed that luciferase activity was significantly lower in cells cotransfected with miR-21-3p and Elovl5 reporter vector, pmiR-RB-REPORT-Elovl5-WT, than in cells with miR-21-3p and reporter control vectors, pmiR-RB-REPORT-Elovl5-mut and pmiR-RB-REPORT-Elovl5-si, indicating that target sites exist in the 3′UTR of Elovl5. Further analysis using qPCR and Western blotting showed that the expression of miR-21-3p negatively correlated with the levels of Elovl5 mRNA and protein, suggesting that miR-21-3p might play an important role in regulating Elovl5 gene expression. Finally, transfection with bta-miR-21-3p mimics, bta-miR-21-3p inhibitor, or miRNA-ShNC in mammary epithelial cells suggested that bta-miR-21-3p promotes triglyceride production, which might be attributed to the expression of the target gene Elovl5.
Introduction
Fat is the most essential form of energy storage (Horton et al., 1995). In milk production, milk fat is an important nutrient and one of the main indicators of the quality of milk (Bauman and Griinari, 2003). In the production of beef cattle, fat is critical for evaluating meat quality (Kalscheur et al., 1997). Therefore, many scholars have studied factors that affect the milk fat percentage and methods for improving the milk fat percentage (Alzahal et al., 2008). These show that lipid metabolism research has important significance in animal fat production (Zak and Pawlak, 2006).
MicroRNAs (miRNAs) are endogenous noncoding RNAs of ∼19–25 nt in length that have been discovered in recent years and are widely involved in post-transcriptional regulation of genes (Yang et al., 2016). Most miRNAs have high sequence conservation, timing of expression, and tissue specificity. They do not encode proteins, and target genes can bind to specific sites in the 3′ noncoding region of miRNAs (Yang et al., 2016). Degradation of the target gene is initiated when it is complementary to the mRNA, and it triggers post-transcriptional gene silencing when it is partially complementary to the target mRNA (Brennecke et al., 2005). Regarding lipid metabolism, miRNAs mainly affect fat, cholesterol synthesis, translocation, and utilization by inhibiting the translation of genes related to lipid metabolism (Krek et al., 2005). Therefore, miRNAs can be used as potential mediators of lipid metabolism regulation (Doench and Sharp, 2004).
MiR-21 belongs to a family of miRNAs that are highly expressed in many types of mammalian cells (Ambros, 2004; Hwang and Mendell, 2006). In previous studies, the expression of miR-21 was associated with human colorectal, breast and lung cancer, as well as human glioblastoma cell lines (Gregory et al., 2013). Studies have suggested that miR-21 may promote cell proliferation in many ways, including pathways such as p53, transforming growth factor-beta (TGF-β), and mitochondrial apoptosis (Mizoguchi et al., 2012). Recently, miR-21-3p was found to be differentially expressed in high-fat and low-fat mammary tissues of dairy cows (Min-JiSong et al., 2014). The high expression in high-fat tissues suggests that miR-21-3p may play an important role in regulating lipid metabolism (Han et al., 2017). Long-chain fatty acid elongase 5 (Elovl5) is the rate-limiting enzyme that catalyzes the elongation reaction of arachidonic acid, oleic acid, stearidonic acid, palmitic acid, linolenic acid, and eicosapentaenoic acid (Jakobsson et al., 2006). These long-chain polyunsaturated fatty acids (PUFAs) are an essential component of the cell membrane and participate in many important signaling processes (Leonard et al., 2000). They play an imperative role in life activities (Carmona-Antoñanzas et al., 2013). Studies have shown that Elovl5 mRNA levels in breast tissue of high-fat dairy cows are significantly lower than in low-fat dairy cows, and Elovl5 negative feedback regulates fat synthesis (Gregory et al., 2013).
Therefore, in this study, we aimed to validate the relationship between miR-21-3p and the target gene Elovl5 and to explore the regulatory mechanisms of miR-21-3p in lipid metabolism in the mammary epithelial cells (MECs).
Materials and Methods
Experimental samples
MECs are isolated and cultured from bovine mammary gland tissue. The MECs of cow cell line determined the cell viability within 5 and 10 generations, and the concentration of MECs was provided by the Laboratory of Animal Genetics in Jilin Agricultural University. MECs were seeded into six-well culture plates (Falcon, Franklin, Lake, NJ) at a concentration of 0.6 × 10^6 cells/well with Dulbecco's Modified Eagle's Medium/Nutrient Mixture F-12 Ham (DMEM/F12, 1:1 mixture) (Hyclone) containing 10% fetal bovine serum (Invitrogen) and cultured at 37°C in a CO2 incubator (Thermo, Marietta, OH). Three replicate wells are used for each assay at least.
Experimental reagents
The PREP RNA mini kit (Analytik Jena, Germany) was used to extract RNA, and the Reverse Transcription reagent kit (Takara Biological, China) was used for cDNA reverse transcription. The FastStart Universal SYBR Green Master from ROX was also used (Roche Company, Germany). The primers were synthesized by GENEWIZ, Inc. (Suzhou, China). The Dual-Luciferase assay kit was obtained from Promega. The Elovl5 antibody was obtained from Abcam.
Bioinformatics prediction of miR-21-3p target genes
We predicted the possible target genes of miR-21-3p by using the online software miRBase (
Vector construction
The vectors bta-miR-21-3p mimics, bta-miR-21-3p inhibitor, and bta-miR-NC were constructed by GenePharma (China). Elovl5 has two target sites for miR-21-3p. Both target site sequences of Elovl5 including Not I and Xho I restriction sites were amplified by PCR. Then, we cloned them into the pmiR-RB-REPORT vector for recombinant vectors pmiR-RB-REPORT-Elovl5-1-mut/WT/si and pmiR-RB-REPORT-Elovl5-2-mut/WT/si.
Luciferase reporter assay
In this experiment, MECs were cultured in DMEM/F12 (HyClone) with 10% FBS (Invitrogen). The pmiR-RB-REPORT-Elovl5-mut/WT/si vectors were cotransfected with miR-21-3p into MECs. We detected the activity of luciferase using the SpectraMax M5 microplate reader.
Total RNA isolation and quantitative real-time PCR
The reverse transcription primers and fluorescence-labeled primers for quantitative analysis of miR-21-3p and target genes were designed by Primer 6.0. All primers were synthesized by GENEWIZ, Inc. (Suzhou, China) (Table 1).
Primer Sequences of Real-Time PCR
Elovl5, ELOVL Fatty Acid Elongase 5.
DNA was transfected into MECs using FuGENE HD transfection reagent (Promega) according to the manufacturer's instructions. MECs were transfected with bta-miR-21-3p mimics, bta-miR-21-3p inhibitor, and miRNA-ShNC. The expression of green fluorescent protein (GFP) in transfected cells was observed after 48 h using a NikonTE2000 fluorescence microscope (Japan). Then, the cells were harvested. Total RNA was extracted according to reagent specifications. The cDNA was synthesized with the Reverse Transcription reagent kit (Takara Biological). After the total RNA was checked by agarose gel electrophoresis and the concentration was measured using a spectrophotometer, the mRNA expression levels of miR-21-3p and target genes were detected by Rox (Roche Company) using an Eppendorf AG-5341. qPCR amplification was performed in a 10-μL reaction with the following procedure: 95°C for 30 s, 40 cycles of 95°C for 5 s, and 60°C for 30 s with β-actin as the reference gene. The relative expression ratios were calculated using the 2−ΔΔCT method according to the following formula: ΔΔCT = (Ct [positive]-Ct [reference])-(Ct [control]-Ct [reference]). The data were analyzed by SPSS19.0 with completely random design of one-way ANOVA among the groups. The mean and standard deviation among triplicate were calculated, and then p values of the groups were calculated for the differential expression analysis.
Western blot analysis
Total protein was extracted from MECs transfected with bta-miR-21-3p mimics, bta-miR-21-3p inhibitor, and miRNA-ShNC using radioimmunoprecipitation assay (RIPA) buffer (BOSTER, China) following reagent specifications. The total protein concentration in the samples was determined with the enhanced bicinchoninic acid protein quantitation assay (KeyGEN BioTECH, China) using a spectrophotometer (UNIC2802H, Shanghai, China). The proteins were resolved by SDS-PAGE and transferred onto a polyvinylidene difluoride membrane (Bio-Rad Laboratories, Inc.). Immunoblotting was performed with antibodies at a 1:500 dilution. The immunoblots were developed using an ECL Advanced Western Blotting Detection Kit (Invitrogen), and the signal intensities were captured by a Tanon 5200 chemiluminescence/fluorescence image analysis system.
Triglyceride detection
Triglyceride was extracted from cells transfected with bta-miR-21-3p mimics, bta-miR-21-3p inhibitor, and miRNA-ShNC according to reagent specifications. We detected the absorbance of the samples using the SpectraMax M5 microplate reader.
Results
Prediction of miR-21-3p target genes and verification of miR-21-3p sites in the 3′ UTR of Elovl5 mRNA using a luciferase reporter assay
To find miR-21-3p target genes, a target site within the 3′UTR that binds to and complements the miR-21-3p seed sequence was chosen for bioinformatics analysis. Elovl5 is a rate-limiting enzyme that catalyzes the reaction of long-chain PUFAs. Elovl5 was selected for further validation due to its potential role in fatty acid and triglyceride formation among all the potential target genes (Fig. 1A). Elovl5 has two miR-21-3p binding sites in the 3′UTR. Cotransfection results showed that the luciferase activity of MECs transfected with the Elovl5-1-WT vector significantly decreased compared with MECs transfected with Elovl5-1-si/mut. There is no significant difference observed between MECs cotransfected with the Elovl5-1-si vector and miR-21-3p and cells cotransfected with the Elovl5-mut vector and miR-21-3p (Fig. 1B). The luciferase activity of MECs transfected with the Elovl5-2-WT vector significantly decreased compared with MECs transfected with Elovl5-2-si/mut. In conclusion, these results further confirmed that Elovl5 is a target gene of miR-21-3p, and the effectual target site sequence was cloned into the pmiR-RB-REPORT-Elovl5-1-mut/WT/si vector.

Target sites of miR-21-3p and luciferase activity.
miR-21-3p downregulates the target gene Elovl5
As shown in Figure 2, we can observe the expression of GFP in MECs transfected with bta-miR-21-3p mimics, bta-miR-21-3p inhibitor, and miRNA-ShNC, confirming successful transfection.
Transfection efficiency. Green fluorescence could be observed under a fluorescence microscope 48 h after transfection. The expression rate of green fluorescence in MECs of dairy cow cells was 70%.
The qPCR analysis results showed that the expression of miR-21-3p in MECs transfected with mimics had dramatically increased compared with cells transfected with inhibitor and shNC. Also, there was a decrease in the expression of miR-21-3p in MECs transfected with inhibitor compared with cells transfected with shNC (Fig. 3A). Subsequent analysis of the target gene showed a significant reduction in the relative expression levels of Elovl5 in MECs transfected with mimics (Fig. 3B). Furthermore, Western blot results demonstrated that MECs transfected with miR-21-3p mimics exhibited lower Elovl5 protein levels (Fig. 3C). These results indicate that miR-21-3p can downregulate the expression of Elovl5 at both the gene and protein levels.
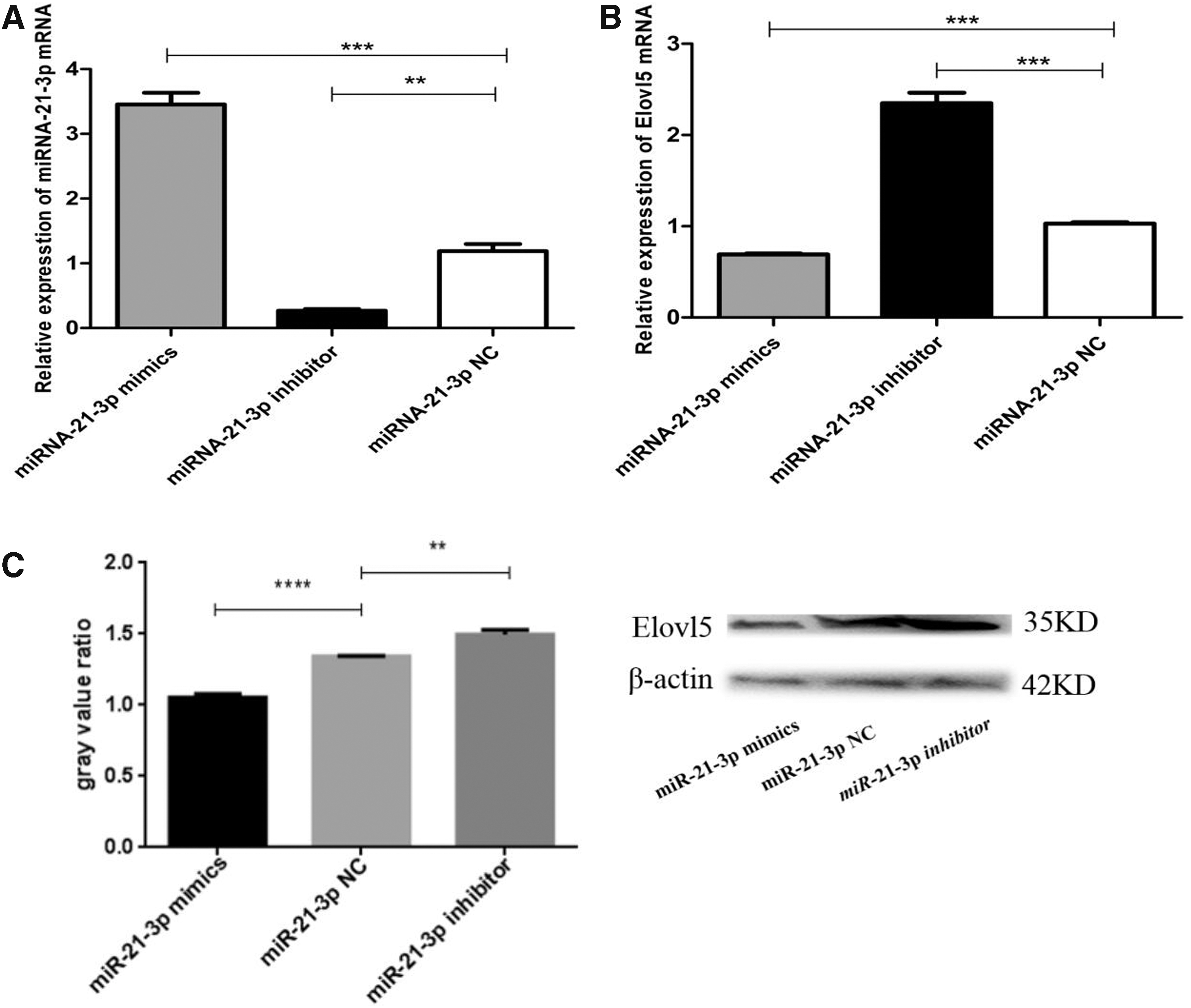
The expression of miR-21-3p and target genes.
Regulation of triglyceride production by miR-21-3p
Triglyceride production was upregulated in MECs transfected with the miR-21-3p mimic. In addition, triglyceride production was downregulated in MECs transfected with the miR-21-3p inhibitor (Fig. 4) (p < 0.05).
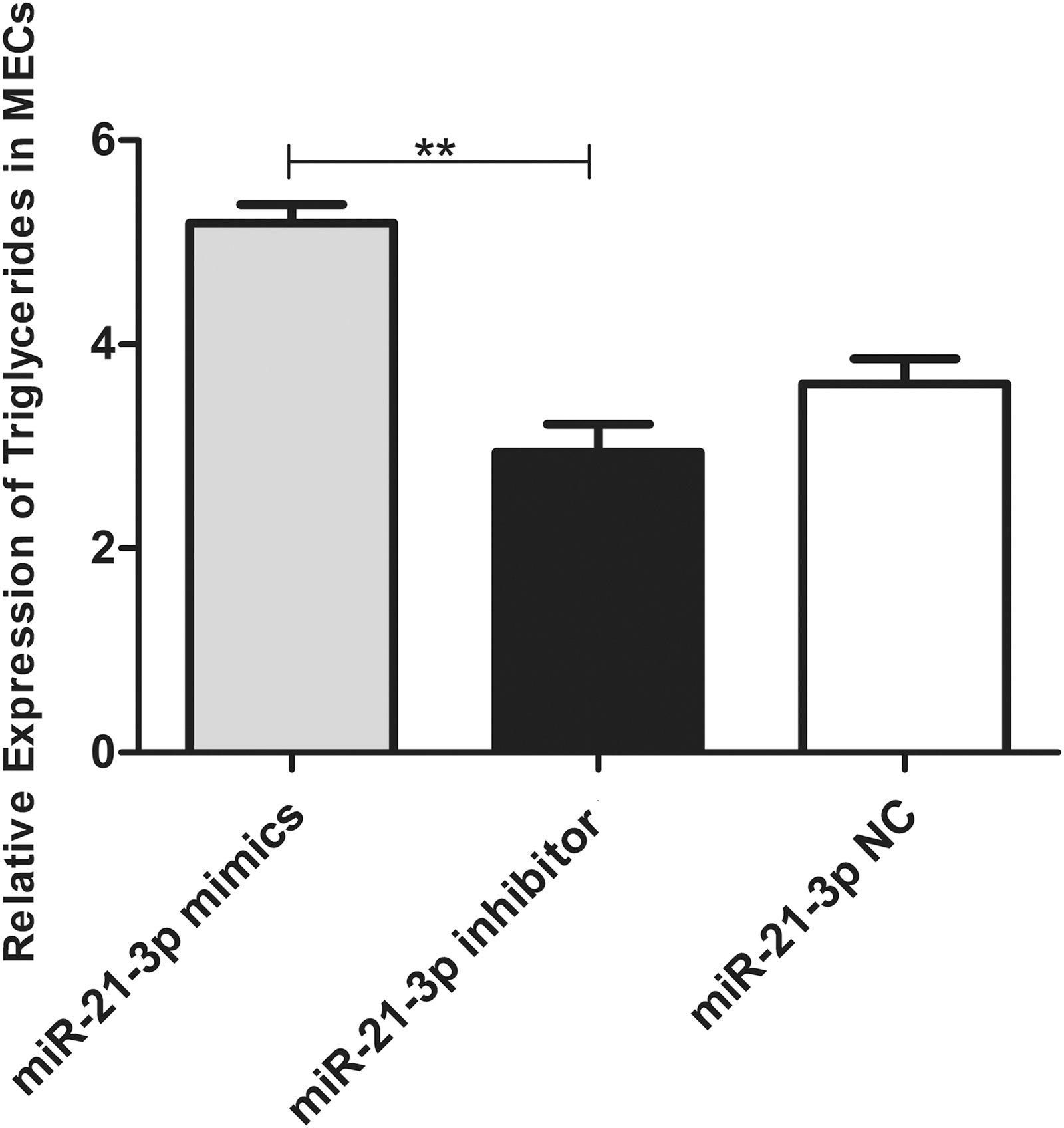
Triglycerides expression in MECs: Triglycerides expression in MECs of transfected bta-miR-21-3p mimics, bta-miR-21-3p inhibitor, and bta-miR-shNC (**p < 0.01).
Discussion
miRNA is a small noncoding RNA involved in the regulation of various biological processes such as adipocyte differentiation and lipid metabolism (Rodriguez et al., 2004). It is also regulated by transcription factors, adipocytokines, and environmental factors (Song et al., 2015). These complex interactions constitute the regulatory network of miRNAs and play a regulatory role in gene expression. Studies have shown that miR-27 (Kim et al., 2010), miR-122 (Gatfield et al., 2009; Benatti et al., 2014), miR-370, miR-33 (Wijesekara et al., 2012), and miR-143 can regulate the level of post-transcriptional regulation of target genes, affect the expression of lipid metabolism-related proteins, and then regulate the body's lipid metabolism. In recent years, it has also been discovered that miRNAs play a crucial role in the regulation of fatty acid and cholesterol metabolism by targeting lipid metabolism genes. Studies have shown that the target genes of miR-122 mainly include AMPK, PMVK, CYP7A1, and PPARβ (Gatfield et al., 2009). After inhibition of miR-122, liver function is affected and cholesterol synthesis efficiency is reduced. MiR-122 is a key miRNA that regulates lipid metabolism and plays an essential role in protecting liver function (Girard et al., 2008).
It has been reported that miRNA-21 may be involved in a variety of regulatory pathways including p53, TGF-β, and mitochondrial apoptosis (Gabriely et al., 2008). miRNA microarray analysis revealed that miR-21 expression was increased after berberine treatment in HepG2 human hepatoma cell lines, and miRNAs can regulate tumor cells (Gabriely et al., 2008). Elov15 is one of the seven fatty acid elongase subtypes expressed in humans and rodents (Holman, 1986). Studies have shown that Elov15 regulates lipid metabolism by regulating TG levels in the liver by controlling the abundance of adipocyte triglyceride lipase, also known as the patatin-like phospholipase domain (Tocher, 2003). Therefore, it is speculated that miRNA-21-3p regulates lipid metabolism through targeted regulation of Elovl5.
First, the presence of the miR-21-3p target site in the 3′UTR of Elovl5 was confirmed by bioinformatics analysis, and the dual luciferase reporter assay further confirmed that Elovl5 was targeted by miR-21-3p. In this study, qPCR and Western blot analyses also verified our results. In MECs, the relative expression level of miR-21-3p is negatively correlated with the mRNA and protein levels of the target gene Elovl5, indicating that Elovl5 is targeted by miR-21-3p. The results showed that miR-21-3p upregulated triglyceride production, confirming that miR-21-3p is involved in the regulation of lipid metabolism.
Conclusion
Elovl5 is an important gene that regulates lipid metabolism and is targeted and regulated by miR-21-3p. Also, miR-21-3p can affect the formation of triglycerides in MECs. Therefore, miR-21-3p can be used as an important indicator to evaluate milk fat quality and mark assisted cattle breeding.
Footnotes
Acknowledgments
We are very grateful to Mengyan Wang of our lab for helping during the revision process. This work was supported by the National Major Special Project on New Varieties Cultivation for Transgenic Organisms (2016ZX08009003-006), the National Natural Science Foundation of China (no. 31372278 and no. 31672389), the National High Technology Research and Development Program (863 Program, no. 2013AA102505), the Jilin Scientific and Technological Development Program (no. 20170519014JH), and the Jilin province industrial technology research and development program (2016C032).
Disclosure Statement
The authors declare that there is no conflict of interest that could affect the research reported.