
Abstract
Congenital anomalies of the kidney and urinary tract (CAKUT) are the leading cause of end-stage renal disease in children. Our group has discovered that Holliday Junction resolvase gene Gen1 is a potential candidate gene for CAKUT. Gen1 mutant mice showed CAKUT phenotypes similar to those observed in retinoic acid (RA)-deficient models. The expression of Raldh2, which encodes the key enzyme in RA synthesis, was reduced in Gen1 mutant metanephros through RNA sequencing. By real-time reverse transcription-PCR, the expression of both Raldh2 and downstream Ret was reduced in embryonic day (E) 11.5 Gen1 mutant ureters and E13.5 kidneys, and expression of RA receptor alpha was decreased in E13.5 Gen1 mutant ureters and kidneys. Further studies showed that all-trans retinoic acid (ATRA) rescued solitary kidney phenotype and improved ureteric branching; ATRA should be administered after ureteric budding to avoid increasing the incidence of ectopic budding in Gen1 mutants. Luciferase intensity of RA response element was lower in CHO-K1 cells transfected with Gen1 siRNA than in those transfected with scrambled RNA, and this inhibitory effect could be reversed by ATRA. These findings indicate that Gen1 mutation can result in renal malformation through RA signaling and Gen1-loss-induced CAKUT can be partly rescued by ATRA.
Introduction
Congenital anomalies of the kidney and urinary tract (CAKUT) are major types of congenital malformations, accounting for 20–30% of abnormalities identified in the prenatal period, and the prevalence of CAKUT is estimated to range between three and six per 1000 births (Heidet et al., 2017). Data from our center have also shown that CAKUT are the leading cause of end-stage renal disease in children, which is in agreement with the findings of other national and regional renal registries (Nicolaou et al., 2015; Vivante et al., 2017). CAKUT constitute a series of structural malformations that are attributed to defects in the development of the upper and lower urinary tracts.
Murine working urinary system begins with the formation of the nephric duct, which extends and joins the cloaca (bladder primordium) by embryonic day (E) 9.5 (Kim et al., 2014). An outgrowth from the nephric duct, the ureteric bud, is induced by the metanephric mesenchyme at E10.5. The ureteric bud will undergo T-shaped branching at E11.5, followed by mesenchymal-to-epithelial transition and branching morphogenesis. Meanwhile, the trunk of the ureteric bud will form the ureters, which will insert into the cloaca through apoptosis of the common nephric duct (CND) for ureter/bladder maturation. These multistage processes require precise modulation at the DNA and posttranscriptional levels.
Gene mutations or exposure to environmental risk factors will disturb these steps and finally cause CAKUT (Nicolaou et al., 2015). To date, over 40 mutant genes have been identified in humans; however, these genes can only explain ∼5–15% of CAKUT cases (Hwang et al., 2014). Moreover, the relations of these known candidate genes and possible pathological mechanisms remain obscure. Thus, much research is still needed to determine the exact mechanism underlying the majority of CAKUT cases and facilitate genetic counseling.
Our group has discovered that Gen1 may be a potential candidate gene for CAKUT (Wang et al., 2018). Disruption of the Holliday Junction resolvase gene Gen1 through a piggyBac (PB) transposon led to renal abnormalities in mice. Compared with wild-type (WT) mice, Gen1PB/PB (Gen1 mutant) mice showed a significantly increased incidence of duplex budding and an obviously decreased number of branches, as determined through RNA in situ hybridization. However, determining the exact mechanism by which Gen1 mutations cause CAKUT requires further study.
Interestingly, Gen1 mutant mice showed CAKUT phenotypes similar to those observed in retinoic acid (RA)-deficient models, including renal hypoplasia, hydronephrosis and megaureter, and abnormal ureter maturation and insertion (Batourina et al., 2005). RNA sequencing showed that Raldh2 expression was obviously decreased among E10.75 Gen1PB/PB metanephros (Supplementary Table S1). Raldh2 encodes the key enzyme involved in the synthesis of RA; this enzyme is part of aldehyde dehydrogenase family 1, subfamily A2. Moreover, Gen1 mutant mice displayed kinky tails, which was a result of delayed neural tube closure. RA signal has been proved important for the development of nervous system. Thus, we speculate that the RA signal might be a potential downstream pathway by which Gen1 modulates renal development.
All-trans retinoic acid (ATRA), the primary bioactive form of vitamin A, plays a key role in regulating a series of embryonic physiological events by activating the nuclear receptor superfamily (Bohnenpoll et al., 2017). These receptors, including the RA receptors and retinoid X receptors, act as transcription factors and will finally modulate the key Gdnf-Ret signal of renal development. In metanephric organogenesis, the administration of ATRA increases ureteric branching and nephron development in a dose-dependent manner; however, others have reported that this modulatory effect of ATRA on renal organogenesis is inhibited at higher concentrations (Vilar et al., 1996).
In this study, we detected the expression of molecules involved in RA signaling, and ATRA was used during nephrogenesis in vivo and in vitro to verify RA as a downstream molecule modulated by Gen1. We found that Gen1 mutation led to specific changes in the E11.5 and E13.5 metanephros of RA signaling molecule expression during metanephric morphological development. Exogenous ATRA could partly rescue the CAKUT phenotypes caused by Gen1 mutation. Moreover, Gen1 siRNA could significantly reduce the luciferase activity of the RA response element (RARE) in CHO-K1 cells, and this inhibitory effect could be reversed by ATRA. These findings suggest that the pathogenesis of CAKUT can be multifactorial and that improving specific environmental factors can ameliorate renal abnormalities. Moreover, these findings may not only provide clues to the reason different mutations in the same gene can have various effects but also might support the development of potential therapeutic approaches for CAKUT.
Materials and Methods
Gen1-deficient mice
Animal experiments were conducted according to the guidelines set by the Animal Care and Use Committee of the Institute of Developmental Biology and Molecular Medicine (IDM) at Fudan University (SYXK 2015–2016). The PB insertion was mapped to the second intron of Gen1 to generate homozygous Gen1 mutant (Gen1PB/PB ) mice, and genotyping was carried out by PCR as previously described (Wang et al., 2018). Hoxb7-EGFP plasmid was a generous gift from Dr. Frank Costantini (Columbia University, New York City, NY). Gen1 mutant and Hoxb7-EGFP (also called Hoxb7) strains were maintained on an FVB/N background. All mice were housed in ventilated cages with up to five mice and a diurnal light cycle providing 12 h of light.
Real-time quantitative reverse transcription-PCR
Molecules, including Raldh2, retinoic acid receptor alpha (RARα), RAR beta (RARβ), Ecm1 and Ret, belong to RA signal, and these molecules have been reported in GUDMAP (
Trypan blue solution injection
To detect the existence of possible ureteric obstruction, 0.4% Trypan Blue solution (Sigma) was injected into the renal pelvis of kidneys. The same speed of drip was controlled by the intravenous infusion set to push the solution through the ureter toward the bladder. The observation time was set to within 5 min.
Apoptosis of the CND
Embryos were dissected at E11.5 and E12.5, fixed in 4% paraformaldehyde (PFA) in phosphate-buffered saline (PBS) overnight, and then rinsed with PBS several times. Metanephric tissues were dissected under a microscope (Leica), soaked in 0.3% Triton X-100, and blocked with 5% donkey serum solution (Jackson) overnight at 4°C on a shaker. Tissues were incubated with anti-caspase3 primary antibody (cell signaling technology [CST], 1:100) overnight at 4°C on a shaker, washed with PBS, and then incubated with Cy5-conjugated donkey anti-rabbit antibody (Jackson; 1:500). Five embryos of each genotype were used for each analysis. Samples were mounted with 50% glycerol/1 × PBS and imaged with an LSM 710 confocal laser scanning microscope using NIH ImageJ software.
Administration of ATRA to observe ureteric budding
Hoxb7;Gen1PB/PB pregnant mice were randomly divided into two experimental groups. Mice in one group were administered low doses of 2 mg/kg ATRA (Sigma) suspended in corn oil, and mice in the other group received an equivalent volume of oil only. The control group consisted of pregnant Hoxb7 mice administered with 2 mg/kg ATRA. The treatments were administered through intraperitoneal injection from E7.5 to observe ureteric budding.
Administration of ATRA to observe CAKUT phenotypes
Hoxb7;Gen1PB/PB pregnant mice were randomly divided into two experimental groups to detect the effect of ATRA on the urinary malformations of Gen1 mutants. Mice in one group were administered 2 mg/kg ATRA suspended in corn oil from E10.5 before birth, and mice in the other group received an equivalent volume of oil only. The newborns of these treated maternal mice were euthanized, and their urinary tracts were dissected out and fixed in 4% PFA for analysis of the CAKUT phenotypes.
Administration of ATRA to observe ureteric branching
Organ cultures were carried out based on previous studies (Vilar et al., 1996). E11.5 Hoxb7;Gen1PB/PB kidney rudiments were dissected from the embryos, explanted onto 0.4-mm polyester membrane Transwell supports, and cultured at the air/liquid interface for 72 h with Dulbecco's modified Eagle's medium/Nutrient Mixture F-12 (DMEM/F-12) (Gibco). ATRA and 9-cis-RA (Sigma) were dissolved in dimethyl sulfoxide (Sigma), and a final concentration of 200 nM was used in the culture medium. The culture was observed every 12 h to record the number of branches from each metanephros through a fluorescence dissecting microscope (Leica).
Generation of stable cell lines and ATRA treatment
CHO-K1 cells were seeded at 60–70% confluency in DMEM/F-12 supplemented with 10% fetal bovine serum (Gibco) and 1% penicillin/streptomycin (Gibco). The pEGFP-C1 plasmid was used to assess the transfection efficiency. The RARE sequence was cloned into the pGL4.26 plasmid to assess total endogenous RA activity. CHO-K1 cells were transiently transfected with these plasmids using Lipofectamine 2000 (Thermo Fisher) following the manufacturer's instructions. Transfected CHO-K1 cells were then treated with 200 μg/mL of hygromycin B (Sangon Biotech) for 7 days (the medium was refreshed every 24 h) to generate stable expression clones. These colonies were selected according to the luciferase activity.
The stable cell lines were transiently transfected with Gen1 siRNA or scrambled RNA (Gene Pharma). The efficiency was detected through cotransfection with pEGFP-C1 plasmid in CHO-K1 cells by flow cytometry and CellQuest Pro software. The RARE-luciferase activity was finally calculated according to the FACS data. Then, these cell lines were treated with 1 μM ATRA or not. Cells were lysed with reporter lysis buffer, and the corresponding signal was detected by the Luciferase Assay System (Promega UK). Firefly luciferase expression was normalized to that in the vehicle control group and is expressed as a fold change.
Statistical analysis
Numerical data are presented as the mean ± standard error of mean. Statistical analysis was performed using GraphPad Prism. An unpaired two-tailed t-test or Fisher exact test was used to determine differences between two groups. The significance level was set at p < 0.05.
Results
Reduced RA signal in Gen1PB/PB metanephros
Among the three isozymes Raldh1–3, Raldh2 is indispensable as it generates RA during fetal life, as demonstrated by its expression pattern in the embryo. The RA signal is vital for metanephric development and has been reported to participate in morphological development during nephric duct insertion, ureteric bud growth, and branching (Batourina et al., 2002, 2005). The expression of Raldh2 was obviously reduced in Gen1PB/PB metanephric tissues, as detected by RNA sequencing. We further extracted the kidneys and ureters of Gen1 mutant and WT mice at different embryonic days to detect potential downstream targets (Fig. 1).

Real-time qRT-PCR analysis. At least five Gen1 mutant and WT metanephric tissues were dissected in one panel. Data were normalized using the reference gene Gapdh. The expression of both Raldh2, which encodes the key enzyme in retinoic acid synthesis, and downstream Ret, were reduced in the E11.5 Gen1 mutant ureters and E13.5 kidneys, and the expression of RARα and Ecm1 was decreased in E13.5 Gen1 mutant ureters and kidneys. Unpaired t-test, *p < 0.05;**p < 0.01. Ecm1, extracellular matrix 1; Homo, Gen1 mutant; Ki, kidney; RARα, retinoic acid receptor alpha; qRT-PCR, quantitative reverse transcription PCR; Ure, ureter; WT, wild type.
Real-time qRT-PCR revealed reductions in Raldh2 expression in the E11.5 ureters and E13.5 kidneys of Gen1 mutant mice compared with WT mice (p < 0.01 and p < 0.05, respectively). The expression of Ret was reduced in the Gen1 mutant E11.5 ureters (p < 0.05) and E13.5 kidneys (p < 0.05). The expression of RARα was reduced and marginally decreased in the Gen1 mutant E13.5 ureters (p < 0.01) and kidneys (p = 0.053), respectively. However, the expression of another RA receptor, RARβ, was not changed in these tissues. Data from RNA sequencing and qRT-PCR also showed that Ecm1 expression was significantly lower in the E13.5 ureters (p < 0.05) and kidneys (p < 0.05) of Gen1 mutants. Extracellular matrix 1 (Ecm1) is a novel RA-regulated target in the metanephros and is considered to modulate the distribution of Ret in the ureteric bud (Paroly et al., 2013). These findings indicate that Gen1 can modulate metanephric development through RA signaling.
Decreased CND apoptosis in Gen1 mutants
The RA signal has also been shown to be important in lower urinary tract development, especially in proper ureter maturation and ureter/bladder junction formation. Perturbations of this process in mice manifest as renal hypoplasia, incorrectly positioned distal ureters, hydronephrosis, and megaureter (Rosselot et al., 2010). Hydroureteronephrosis can be divided into functional or physical obstruction. To determine whether a physical obstruction existed in Gen1 mutants, we then performed intrapelvic dye injections. Among nearly 1/3 Gen1 mutants with hydroureteronephrosis, the injected dye was not detected in the bladder even after prolonged observation (Fig. 2A, B).
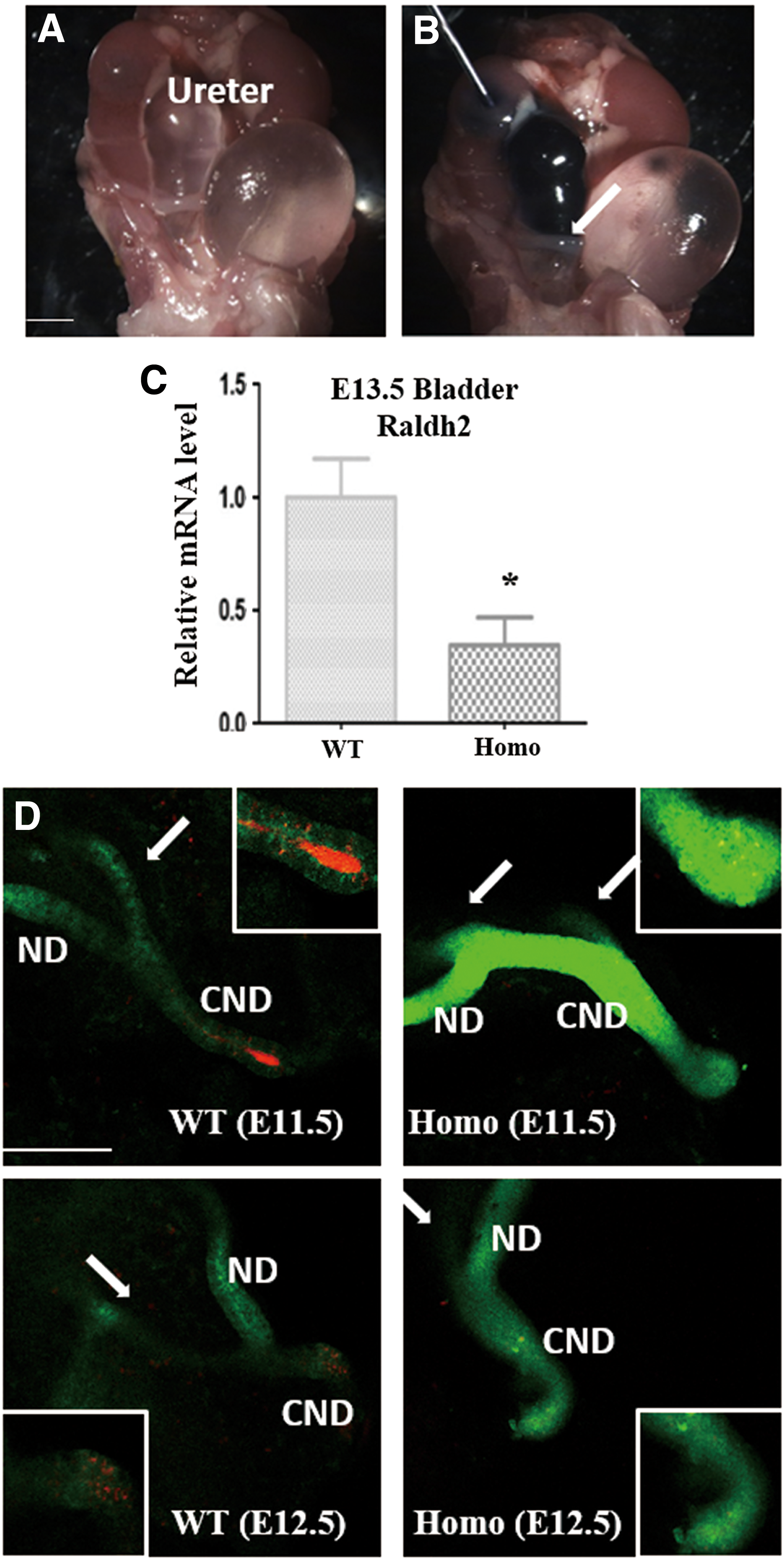
Decreased CND apoptosis in Gen1 mutants. Macroscopic view of the urogenital system in Gen1 mutants exhibited hydroureteronephrosis
Apoptosis is one of the key processes involved in CND elimination and ureter/bladder junction formation. To observe and analyze the CND dynamically, mating between Gen1 mutant and Hoxb7 transgenic mice was required and could display the morphology of the nephric duct, ureteric bud, and their derivatives (Srinivas et al., 1999). Then, we detected CND apoptosis by whole-mount staining, and Cy5 antibody was used to detect the expression of anti-caspase3. The signal intensity of the active anti-caspase3 was significantly reduced in the Gen1 mutants compared with the controls (Fig. 2D). Moreover, the expression of Raldh2 was obviously reduced in the Gen1 mutant bladders (p < 0.05) (Fig. 2C). Taken together, these results suggest that Gen1 mutation led to defects in the signaling of RA signal and its downstream molecule Ret, as well as CND apoptosis, which finally caused urinary tract malformations.
Improvement of ureteric branching in Gen1 mutants by exogenous RA
Offspring of vitamin A-deficient rodents will have renal malformations sometimes with genital anomalies. These malformations can be reversed with vitamin A or RA supplementation during metanephric development to some extent. We attempted to identify the critical window for treating Gen1 mutants with exogenous ATRA to improve metanephric organogenesis. Raldh2 expression can be detected at E7.5 in murine embryos, demonstrating that the fetus not only requires RA from maternal circulation but also from its own synthesis during embryonic development. Moreover, nephric ducts form at the caudal end of the intermediate mesoderm from E9.5 in mice. Therefore, we chose E7.5 as the starting point for ATRA administration.
Surprisingly, ATRA treatment increased the incidence of bilateral duplex ureteric budding among the Gen1 mutant mice (Fig. 3A), whereas when supplemented with oil alone, the mice presented incidences of ectopic ureteric budding similar to those in previous studies (Fig. 3B). Meanwhile, Hoxb7 mice treated with ATRA, all showed normal budding (Fig. 3C). E9.5 was selected as the second time point for ATRA administration, and the findings were similar to those found for E7.5. Administration of ATRA from either E7.5 or E9.5 unavoidably increased the incidence of duplex ureteric budding in Gen1 mutants (p < 0.001) (Fig. 3D). Thus, in Gen1 mutants a low dose of ATRA should be administered until budding occurs. We suggested E10.5 as the correct time point for ATRA administration, yielding an incidence of duplex budding comparable to that in the oil group.

Hoxb7;Gen1PB/PB
mice treated with ATRA during nephrogenesis in vivo. ATRA treatment from E7.5 increased the incidence of bilateral duplex ureteric budding (white arrow) among the Hoxb7;Gen1PB/PB
mice
We performed an analysis of phenotypes in Gen1 mutant newborns treated with ATRA or oil alone from E10.5. The distribution of CAKUT phenotypes showed significant differences between these two groups using Fisher's exact test (p < 0.05) (Fig. 3E–I). Then, two–two comparisons were further compared between different CAKUT phenotypes, and the incidence of solitary kidney was significantly lower among Gen1 mutants treated with ATRA. Among these Gen1 mutant newborns, the solitary kidney phenotype was not detected, which might be attributed to the further ureteric branching induced through RA supplementation. Analysis of the branching patterns of Hoxb7;Gen1 mutant cultures with and without RA verified this speculation.
Cultured in vitro from E11.5, some metanephros without RA revealed branching defects by 48 h, and fewer ampullae could be detected by 72 h. We found that without duplex budding, supplementation with RA could obviously increase the branching, and RA administration helped branches to extend continuously and angle laterally to communicate with one another (Fig. 4A′, C′, and E); however, with unilateral duplex budding, branchings on both sides of the metanephros were limited even when RA was administered (Fig. 4B′, D′). Moreover, RA supplementation slightly increased Gen1 expression in these metanephroi. These results demonstrate that RA can partially rescue the branching defects caused by Gen1 mutation.
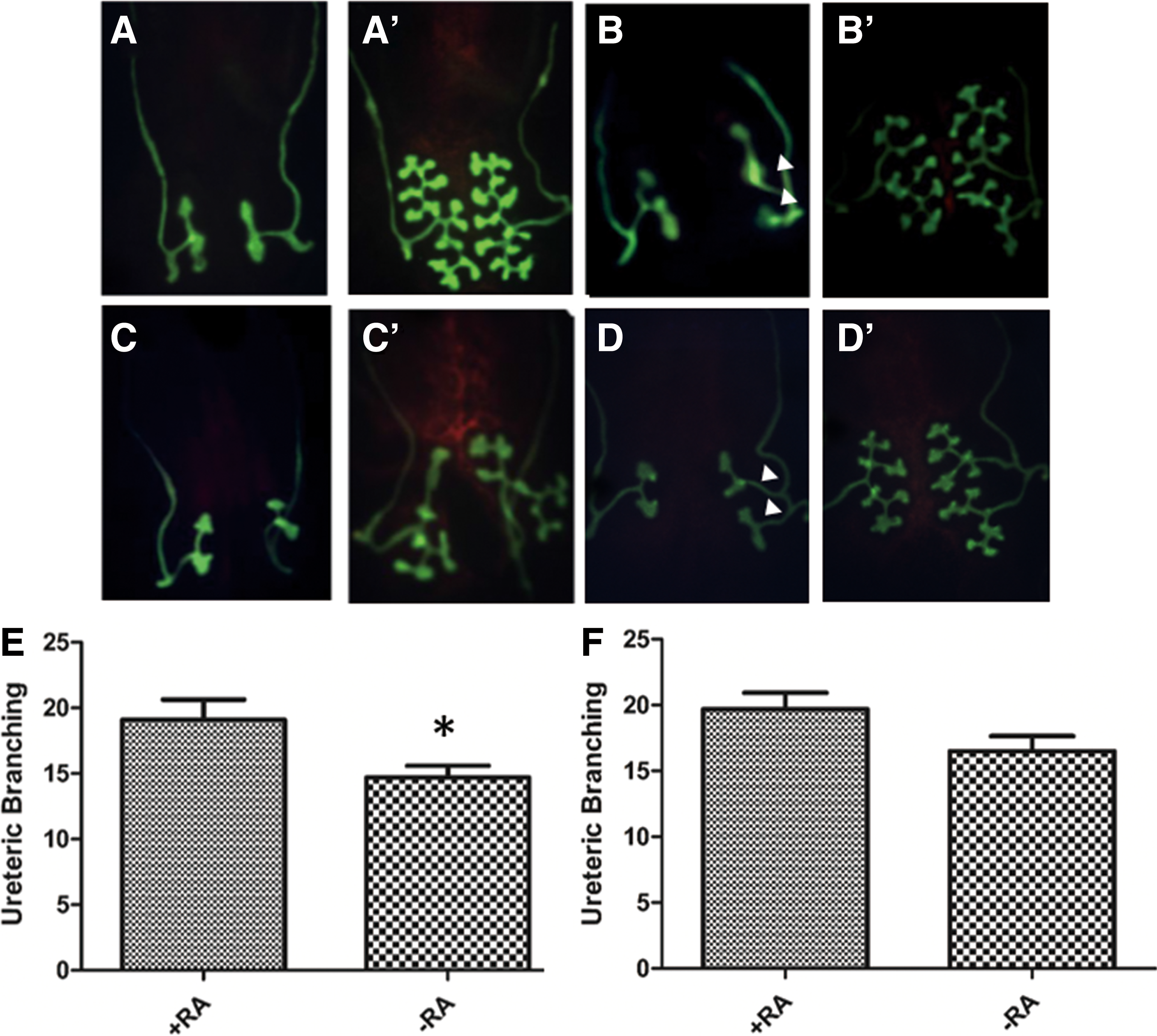
Improvement of ureteric branching in Gen1 mutants by exogenous RA in vitro. Hoxb7;Gen1PB/PB
kidney rudiments were cultured for 72 h in F12/DMEM with RA (ATRA +9-cis-RA) dissolved in DMSO
RA-dependent transcriptional activity affected by Gen1 mutation
RA receptors form heterodimers binding to the regulatory DNA segment, RARE, which can reflect RA-dependent transcriptional activity, and after binding to RARE, these heterodimers can modulate both target gene transcription and crosstalk with other signaling pathways (Wong et al., 2012). Accordingly, we next determined whether Gen1 mutation would affect RA-dependent transcriptional activity using the reporter plasmid pGL4.26 with the RARE sequence. Compared with cells transfected with scrambled RNA, CHO-K1 cells transfected with Gen1 siRNA showed significantly reduced Gen1 expression (p < 0.01) and slightly reduced Raldh2 expression (Fig. 5A). Indeed, the RARE-luciferase activity was obviously reduced when CHO-K1 cells were transfected with Gen1 siRNA (p < 0.05) (Fig. 5B). After CHO-K1 cells transfected with Gen1 siRNA were treated with exogenous 1 μM ATRA for 18 h, the activity was similar to that in the controls and did not show a significant difference (Fig. 5C), suggesting that the inhibitory effect of Gen1 siRNA could be reversed by ATRA supplementation.

RARE-luciferase activity inhibited by Gen1 siRNA and reversed by ATRA administration. CHO-K1 cells transfected with Gen1 siRNA showed significantly reduced Gen1 and Raldh2 expression, respectively
Discussion
RA signaling is critical for various physiological events, especially during embryonic development. RA signaling has been reported to affect the morphology of both the upper and lower urinary tracts, including during ureteric growth and development of the connection between the ureters and bladder (Rosselot et al., 2010). Previous studies have demonstrated that signaling between stromal cells that synthesize RA and ureteric bud cells that respond to RA is vital for regulating ureteric bud formation and branching through the control of Ret expression (Mendelsohn et al., 1999). Gen1 is a newly reported candidate gene for CAKUT; however, its exact mechanism of action is still obscure.
Gen1 disruption leads to ectopic budding, reduced branching, and abnormal CND apoptosis. Gen1 mutant mice show CAKUT phenotypes similar to those observed in RA-deficient models. Our RNA sequencing data show that the expression of Raldh2, a key enzyme for RA synthesis, and the novel RA-regulated target gene Ecm1 is obviously reduced in Gen1 mutant metanephroi compared with WT metanephroi. Thus, we first detected the expression of RA signaling molecules and found that Raldh2, RARα, Ret, and Ecm1 expression was reduced in the Gen1 mutant metanephric tissues. These results suggest that RA is a potential downstream target of Gen1 for modulating renal development.
RA activity was detected during the earliest stages of metanephric development before ureteric budding was observed (Lee et al., 2012). Therefore, we provided a low dose of ATRA to Gen1 mutants starting from E7.5; however, the findings were unexpected: Gen1 mutant metanephroi showed bilateral ectopic ureteric budding. Even after supplementation with ATRA from E9.5, at the onset of kidney formation for primary ureteric bud outgrowth, the result was similar. However, Hoxb7 mice did not show ectopic ureteric budding with the administration of low doses of ATRA. Finally, we delayed the time of administration until ureteric budding had finished. We speculate that ATRA may act as a specific environmental factor that increases the incidence of ectopic budding of Gen1 mutants. This phenomenon that was only observed in Gen1 mutants, or also can be applied in other CAKUT animal models, requires further study. Moreover, the overexposure of WT mice to RA at E9.0 has been reported to cause abnormal nascent metanephros apoptosis and finally bilateral renal agenesis (Lee et al., 2012). These results further verify that the ATRA dose plays a role in metanephric development.
Nascent ureteric branches have a T shape and will contact one another after further branching. Ureteric branch morphology will continue to change over the course of renal development; however, the exact factors that cause these changes are not clear. Previous studies have reported that RA stimulates further ureteric branching without serum in vitro, and RA receptors are considered to mediate this effect by regulating Ret expression in metanephric tissues (Takayama et al., 2014). Mid-gestation RA treatment has shown a similar effect, as the ureteric bud begins initial branching at this time. Moreover, renal development in the postnatal preterm kidney may not be affected by RA or vitamin A administration (Gray et al., 2012). Importantly, our previous studies have revealed that Gen1 mutant metanephric tissue will display distorted ureteric branching or abnormal ureteric tip formation. Thus, low-dose ATRA supplementation from E10.5 was adopted for subsequent rescue experiments in vivo.
The solitary kidney phenotype in Gen1 mutant mice was ameliorated by ATRA administration, and we believe that this effect may be attributed to improved ureteric branching. After supplementation with exogenous ATRA from E11.5, the whole metanephros without duplex budding grew larger, and more ureteric branch tips were detected; however, the branching was not as complete as that in the WT metanephros. Moreover, the incidence of renal hypoplasia/dysplasia in Gen1 mutant newborns did not change with ATRA supplementation. The fact that the rescue was not complete indicates the following: (1) other pathways may be involved in the morphology; (2) the dose of ATRA may not be sufficient to induce complete branching; or (3) disturbances in RA signaling between the metanephric mesenchyme and ureteric bud may lead to other secondary changes.
Correct ureterovesical junction formation depends on the connection between the nephric duct and cloaca. The ureteric bud is indirectly connected to the cloaca through the CND. Defects in the morphology of the CND may cause lower urinary tract malformations, such as vesicoureteral reflux or obstruction. RA and downstream Ret signaling have been verified to regulate the morphology of the lower urinary tract, especially through remodeling of the CND (Kim et al., 2014). Moreover, endogenous renal RA receptor activity has been found in collecting duct cells derived from the ureteric bud. Our studies demonstrate that CND apoptosis is obviously reduced in Gen1 mutants. Compared with WT ureters, the expression of Raldh2, RARα, and Ret was also decreased in Gen1 mutant ureters.
Indeed, our RNA sequencing data did not reveal abnormal expression levels of Gata3, EphA4/EphB2, RPTP-LAR, and Yap/Taz, or clues of RET1015 defects, which have been reported to participate in the modulation of CND morphology (Reginensi et al., 2015; Stewart et al., 2017). These findings suggest that defects in CND survival and apoptosis can be attributed to abnormal RA signaling caused by Gen1 mutation. By extension, we speculate that RA production and/or secretion by Gen1-expressing mesenchymal cells can mediate CND apoptosis through regulating RA receptor activity in ureteric epithelial cells. However, we cannot rule out the possibility that Gen1 may modulate these molecular activities at a posttranslational level. Hydroureteronephrosis was first detected in Gen1 mutants at E16.5, and the incidence of this condition was not decreased with RA supplementation. We infer that this effect may be attributed to ectopic ureteric budding or even nephric duct elongation. Perhaps the time point selected was not optimal for rescuing these defects; however, earlier supplementation with ATRA will inevitably increase the risk of ectopic budding.
RA signaling has also been shown to be involved in neurulation and subsequent neural tube development and to participate in the morphological development of the spinal cord and somites (Santos-Guzman et al., 2003). These observations reveal that neurogenesis requires a relatively high level of endogenous RA activity to maintain embryonic development (Wilson et al., 2003). Aside from kidney and urinary tract defects, Gen1PB/PB mice also have kinky tails as a result of delayed neural tube closure, whereas kinky tails have not been detected in Gen1PB/+ mice (Wang et al., 2018). However, treatment with ATRA cannot rescue the phenotype of neural defects, such as kidney anomalies, in Gen1 mutants. These findings may suggest that ATRA has a different threshold for renal development and neurogenesis or that other mechanisms are involved in neural tube development. Moreover, RA is known as a teratogen during embryonic development. Thus, at least low-dose exogenous ATRA does not increase the severity of neural defects or the incidence of other abnormalities in Gen1 mutant mice.
Conclusion
In summary, our findings demonstrate that Gen1 mutation can lead to specific changes in the E11.5 and E13.5 metanephros of RA signaling molecule expression (Raldh2-RARα-Ret) during metanephric organogenesis. These studies also suggest that CAKUT induced by Gen1 mutation can be partially rescued by exogenous ATRA; however, more investigation is required to determine the optimal timing and dose for ATRA administration. Indeed, Gen1 mutations can cause CAKUT through RA transcriptional signaling in mice, confirming a possible gene/environment interaction that could help explain the heterogeneity of renal malformations. Further studies are required to understand the complexity of the genetic network and multiple factors, for example, environmental factors, involved in CAKUT.
Footnotes
Acknowledgments
The authors thank Dr. Zhisheng Ye, Mr. Xiaoxiang Song, and Mr. Wuhou Dai for technical assistance. This work was supported by Key Developing Discipline of Shanghai Municipal Commission of Health and Family Planning (Pediatrics, 2016ZB0101), Shanghai Natural Science Foundation (15ZR1404400), and the National Natural Science Foundation (81400684 and 81670609).
Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.