
Abstract
Resistance to trastuzumab has become a limiting factor for therapeutic efficacy of human epidermal growth factor 2 (HER2)-positive breast cancer. Different expression levels of miRNAs in cancer cells have been associated with poor prognosis and response to chemotherapy. The aim of this study was to evaluate miRNAs that were thought to be associated with HER2-positive breast cancer chemoresistance. In this study, the relative expression of candidate miRNAs to U6 RNA was evaluated in trastuzumab-resistant and trastuzumab-sensitive cells using relative real-time PCR. Our results demonstrated that miR-23b-3p, miR-195-5p, miR-656-5p, and miR-340-5p were significantly dysregulated. For the first time in this study, these miRNAs were identified to be involved in trastuzumab resistance. TargetScan and miRDB were then used for predicting the potential targets of the candidate miRNAs. Our results also revealed that the predicted potential targets of these miRNAs were strongly associated with drug resistance pathways. As a relative expression of candidate miRNAs was statistically different in trastuzumab-resistant and trastuzumab-sensitive cells, their potential targets were involved in drug resistance pathways. We strongly hypothesized the dysregulation of miRNAs as a possible mechanism of trastuzumab resistance. We also assumed that the strategic manipulation of these regulatory networks might be a possible therapeutic strategy to improve the results of chemotherapy for this resistance. However, more research is needed to evaluate the role of these miRNAs in the acquisition of trastuzumab resistance.
Introduction
Human epidermal growth factor 2 (HER2)-positive breast cancer is considered as a refractory disease, which displays invasive behavior (Hynes and Lane, 2005). Currently, the standard treatment for HER2-positive breast cancer is trastuzumab. The response rate of trastuzumab monotherapy or combination therapy is about 30% to 50% in patients with early HER2-positive breast cancer. However, the majority of the patients display resistance behavior to trastuzumab within 1 year. For that reason, determining the mechanisms of trastuzumab resistance is of great importance, which may help find therapeutic strategies for the treatment of breast cancer (Ju et al., 2018).
Several mechanisms have been distinguished for trastuzumab resistance, including dysregulation of downstream signaling pathways, oligomerization with other epidermal growth factor (EGF) family members and alternative pathways (Arteaga et al., 2011).
miRNAs are short noncoding RNAs of ∼20–25 nucleotides, which potentially regulate protein synthesis at multiple post-transcriptional levels (Iorio et al., 2005). In many cases, miRNAs are reported to be associated with dysregulation in various cancers. These miRNAs can serve as either tumor suppressors, by silencing the oncogenes, or oncogenes, by targeting tumor suppressor genes (Peng and Croce, 2016). Recent discoveries of miRNAs have opened new perspectives on the efficient use of miRNAs in various aspects of cancer, including diagnosis, metastasis, progression, treatment, and drug resistance (Michael et al., 2003). Overexpression of miR-205 could enhance the chemoresistance of nonsmall cell lung cancer cells by targeting phosphatase and tensin homolog (PTEN) (Lei et al., 2013). Chen et al. (2014) showed that upregulation of miR-181a in human cervical squamous cell carcinoma enhanced the chemoresistance to cisplatin by targeting Protein Kinase C Delta (PRKCD).
miR-141 has been reported to be involved in cisplatin resistance by targeting KEAP1 in ovarian cancer cells (Van Jaarsveld et al., 2013); miR-106a enhances multidrug resistance in gastric cancer through repression of RUNX3 (Zhang et al., 2013). In contrast, miR-200b and miR-15b effectively reverse chemotherapy-induced epithelial-mesenchymal transition by targeting BMI1 in human tongue cancer cells (Sun et al., 2012). Several studies have shed light on the emerging role of miRNAs in drug resistance. However, further studies are needed to recognize other miRNAs, their critical molecular targets, and their biological process that they affect.
In this study, we investigated the expression patterns of candidate miRNAs in the human breast cancer cell line BT-474 and its trastuzumab-resistant variant BT-474-R. In these cell lines, we explored the potential of specific miRNAs to serve as predictive markers for resistance to trastuzumab treatment cancer. We assessed HER2-positive breast cancer cells with increased invasiveness and metastatic potential (Olayioye et al., 2000), screened for candidate miRNAs responsible for these malignant phenotypes, and focused on the role of miRNAs involving in metastatic breast cancer. Five out of six candidate miRNAs, including miR-340 (Maskey et al., 2017), -27a (Tang et al., 2012), -34a (Xu et al., 2018), miR-21 (Petrovic et al., 2014), miR-195-5p (Marques et al., 2018), and miR-23b (Pincini et al., 2013) were previously shown to be associated with invasiveness in breast cancer cells in vitro, and another miRNA (miR-656) identified through our in silico analysis of published gene expression changes in breast cancer cell line after neoadjuvant chemotherapy. Bioinformatic analyses were accomplished to predict gene targets of the dysregulated miRNAs and to investigate their potential roles in the acquisition of drug resistance.
Materials and Methods
Cell culture and generation of trastuzumab-resistant cells
The human breast cancer BT-474 cells were obtained from the Iranian Biological Research Center. BT-474 cells were cultured in DMEM-F12 media supplemented with 10% FBS. The cell line was incubated under a humidified atmosphere containing 5% CO2 at 37°C. Trastuzumab/Herceptin (Roche, Basel, Switzerland) was dissolved in sterile water. Trastuzumab-resistant cells were generated by culture of BT-474 cells continuously in the presence of 5 μg/mL trastuzumab for 6 months, as reported previously (Gong et al., 2011). Afterward, trastuzumab-resistant (BT-474-R) and sensitive BT-474 cells were cultured with or without trastuzumab, respectively.
RNA extraction and quantitative real-time PCR
Total RNA was isolated from BT-474 and BT-474-R cells by treatment with the Trizol reagent (Invitrogen, Carlsbad, CA). The isolated RNA samples were reverse-transcribed into double-stranded (ds) cDNA with the miScript II RT Kit (Qiagen). The samples were subjected to quantitative PCR (qPCR) in the StepOne™ Real-Time PCR System (Applied Biosystems, Inc., Hercules, CA). cDNAs were then amplified and detected using SYBR Green PCR Master Mix (Thermo Fisher, England). Mature miRNAs were detected by the forward primers of miR-195-5pF, miR-23b-3pF, miR-27a-3pF, miR-34a-5pF, miR-340-5pF, miR-21-5pF, and miR-656-5pF shown in Table 1 and the MiScript Universal Primer (of the miScript II RT Kit) used as the reverse primer for all miRNAs. U6 RNA was used as internal loading controls for miRNAs. Relative abundance of detected transcripts was calculated using the 2−ΔΔCt method. Quantitative real-time PCR (qRT-PCR) was performed in triplicate.
Primer Sequences for Quantitative Real-Time Polymerase Chain Reaction
Cell survival assay
Cell survival was quantitated using a colorimetric 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay according to the procedures. MTT was purchased from Atocel (Graz, Austria). One hundred microliters of target cell suspension (1 × 104 cells) were added to each well of a 96-well plate, and the plate was incubated for 24 h at 37°C in a humidified 5% CO2 atmosphere. The cells were incubated for another 72 h with medium containing trastuzumab (0.21–2100 μg/mL). After drug treatment, 100 μL of 10 μM MTT working solution was added in each well and the plates were incubated for 4 h at 37°C. After addition of dimethyl sulfoxide (DMSO), the mixture was agitated for 45 min at room temperature. The absorbance values were read with a microplate reader at 570 nm. The percentage of survival was calculated using the following formula: survival percentage = (absorbance of drug-treated wells – blank wells)/(absorbance of untreated wells – blank wells) × 100. The half maximal inhibitory concentration (IC50 values) was measured by the ED50 Plus v1.0 online software.
Prediction of target genes
We used TargetScan (Release 7.0,
Venny analysis
A Venn Diagram analysis “Venny
KEGG pathway analysis of target genes
Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis was done to compare the specific miRNAs targets with the whole reference genes background using Database for Annotation, Visualization and Integrated Discovery (DAVID; version 6.7,
EnrichR clustergram
Analysis of gene set EnrichR Clustergram was performed by EnrichR API (Chen et al., 2013a), using the complete list of gene targets of the dysregulated miRNAs. Gene targets and biological pathways are shown in Figure 4B.
Statistical analysis
Statistical analyses were done using SPSS for Windows version 24. The t-test was used to analyze cells sensitivity to trastuzumab and the significance of differences among the qPCR results was assessed by Mann–Whitney test. Only the dysregulated miRNAs with a fold change >2 and adjusted p-value <0.05 were considered as statistically significant.
Results
Trastuzumab-resistant breast cancer cells display survival over parental cells
The resistant cells demonstrated the same growth rate as the parental cells in the trastuzumab-free cell culture media. To define cell proliferation, parental and resistance BT-474 cells were treated with trastuzumab for 72 h, and cell survival was evaluated by MTT assay. Through calculation, the IC50 of trastuzumab on BT-474 cells and BT-474-R was 1.78 and 15.261 mg/L, respectively. As shown in Figure 1, the parental cells showed a significant more reduction in cell proliferation than resistant cells (p < 0.01). At the highest dose of trastuzumab (2100 μg/mL), about 80% of the resistant cells remained viable, whereas only about 44% of the sensitive cells survived. These results suggest that trastuzumab-resistant HER2-positive breast cancer cells exhibit proliferation advantages in vitro over parental cells.
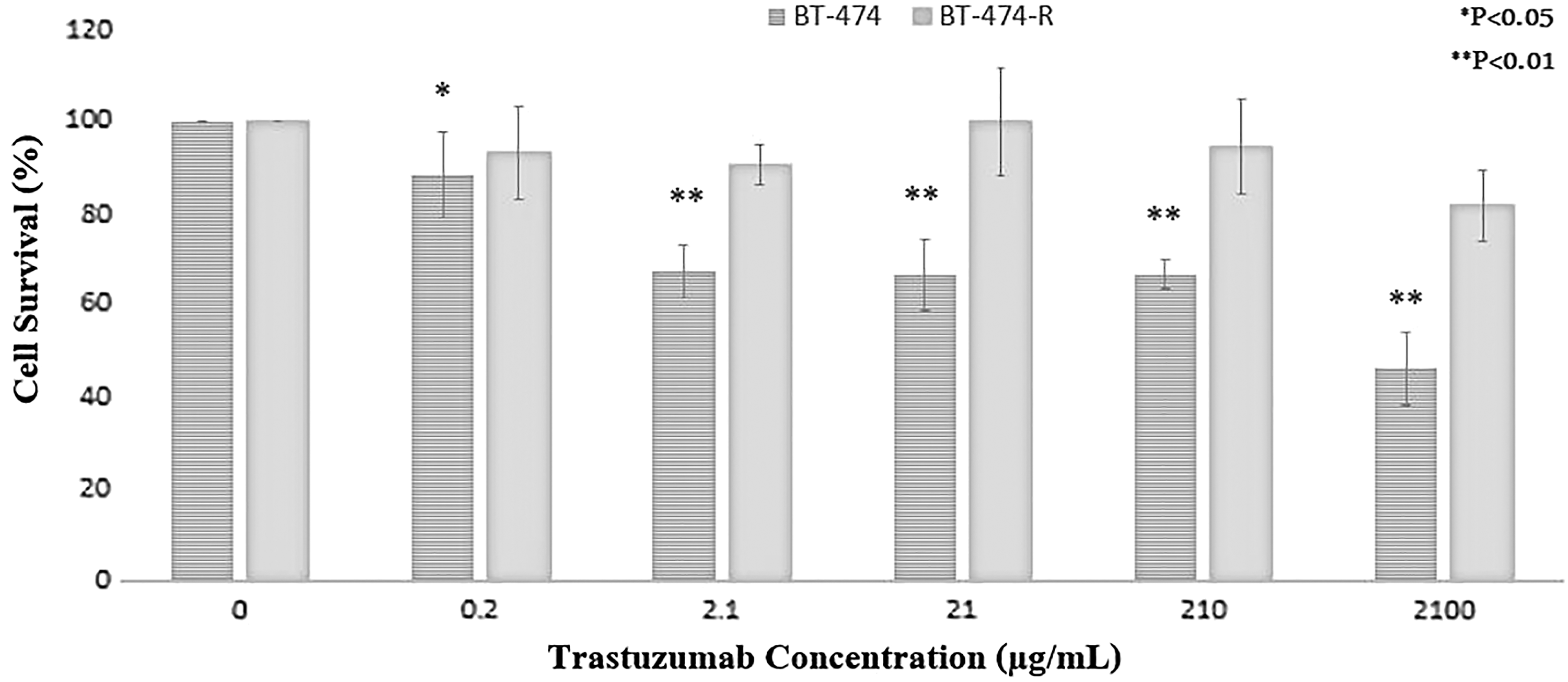
Analysis of sensitivity to trastuzumab by MTT cell survival assay. Cells were plated in 96-well plates and trastuzumab (0.21–2100 μg/mL)-containing medium was added 24 h later. MTT assay was performed after 72 h incubation. *p < 0.05 or **p < 0.01 statistically significant when compared between two groups using t-test. MTT, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide.
Expression status of the seven selected miRNAs detected by qRT-PCR
As mentioned earlier, seven functionally related miRNAs involved in chemoresistance were chosen. The expression status of the seven specific selected miRNAs was then evaluated in the resistant and sensitive BT-474 cells. Within this subset of miRNAs, miR-23b-3p was significantly upregulated in the BT-474_R cells, whereas miR-656-5p, miR-340-5p, and miR-195-5p were downregulated (Fig. 2).

Relative expression of seven miRs dysregulated in BT-474 and BT-474-R.
Analysis of involved pathways
The target genes of four significant dysregulated miRNAs shown in Figure 2C were compared with the whole reference gene background. To recognize possible pathways involved in drug resistance, Cytoscape software V3.5.1 was then applied to decipher the KEGG pathway and explain their biological functions. The predicted target genes of the four selected miRNAs were involved in 27 pathways with p-value <0.05 (Fig. 3).
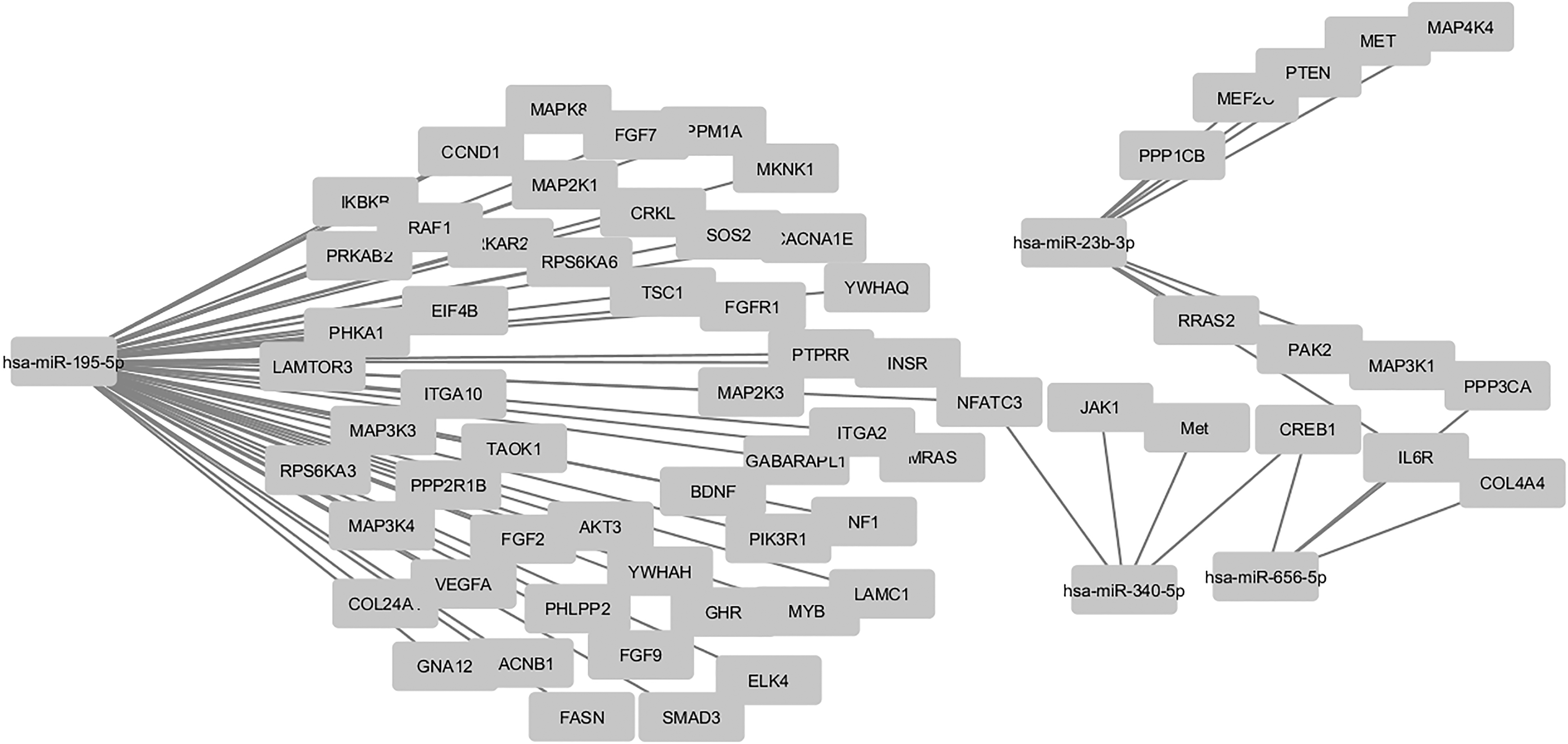
The network between the specific deregulated miRNAs and their predicted targets in this study. Selected overexpressed (miR-23b-3p) and underexpressed (miR-656-5p, miR-195-5p, and miR-340-5p) miRNAs are used to reconstruct the regulatory network. Their experimentally validated target mRNAs show various expression patterns: some are stably expressed, and others are up- or downregulated.
Analysis of EnrichR clustergram
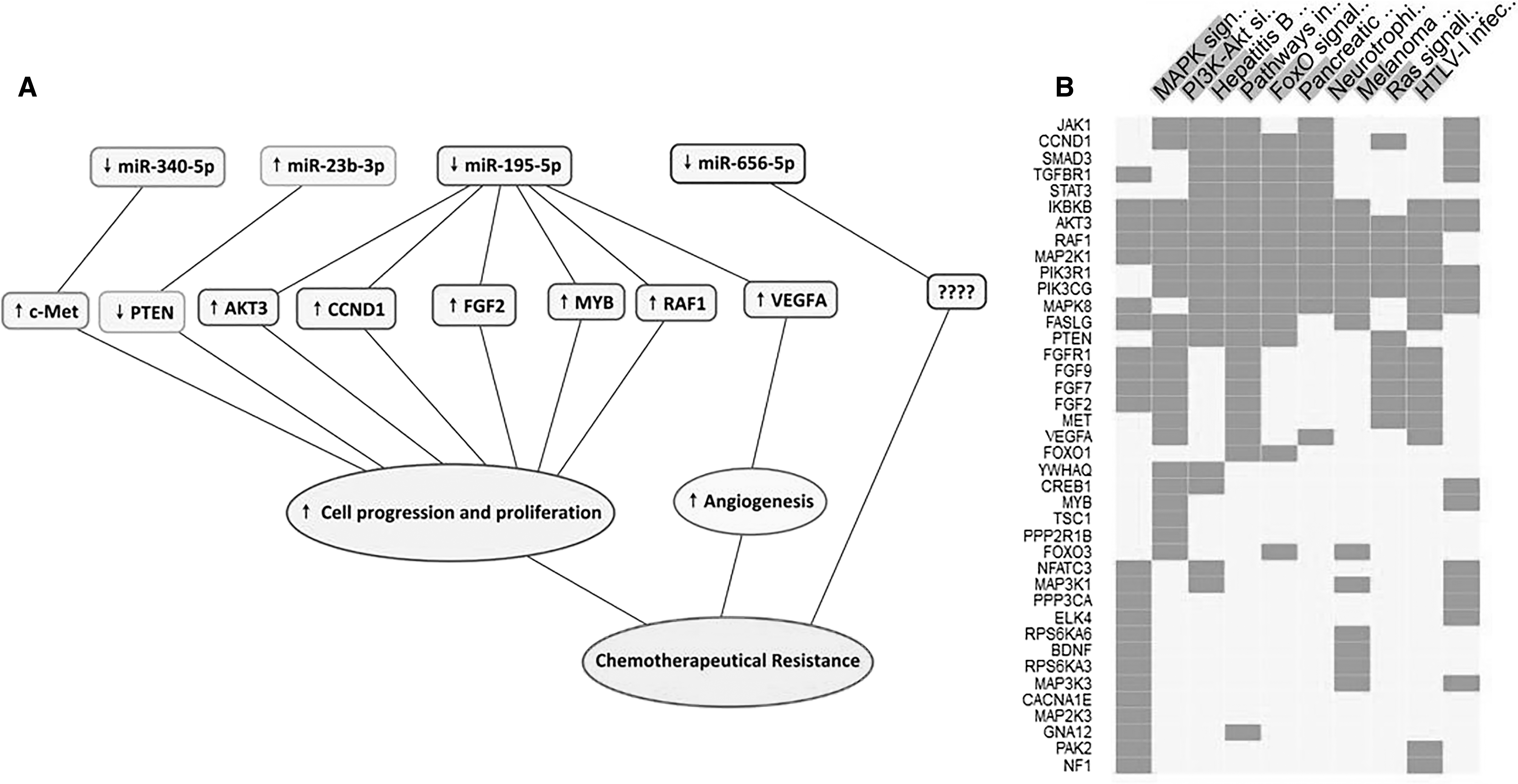
The most enriched pathways related to chemoresistance were MAPK, PI3K-AKT, and foxO signaling pathways. The response to MAPK and PI3K-AKT pathways has the most intense interactions. As shown in Figure 4B, gene clusters corresponding to the top enriched biological pathways revealed.
Discussion
HER2 overexpressing breast cancers display a pathological type with high metastasis incidence and poor prognosis. Trastuzumab as a monoclonal HER2 antibody has the potent tumor-suppressive effect, but the primary and acquired resistance is the major clinical obstacle (Eccles, 2002).
miRNAs can regulate a wide variety of biological functions, such as survival, metastasis, tumor progression, and cancer cell differentiation (Valastyan et al., 2009). Recently, miRNAs have become as key mediators in controlling the cellular responses of cancer cells to treatment. Patient's response to chemotherapy has confirmed to be closely associated with the functional situation of miRNAs (Chen et al., 2013b). The mechanisms of drug resistance regulated by miRNA are still largely unclear.
In this study, we showed that the expression levels of miR-23b-3p, miR-195-5p, miR-656-5p, and miR-340-5p were significantly dysregulated in HER2-positive breast cancer resistant to BT-474 cells. Pathway mapping tool was applied to recognize appropriate pathways of the gene targets of the miRNAs with a significant expression between BT-474 and BT-474-R. The result demonstrated that the predicted target genes of four miRNAs were supplemented in 27 pathways. Among these pathways, PI3K-AKT (Guerrero-Zotano et al., 2016), MAPK (Javidi-Sharifi et al., 2015), and foxO (Zhao and Lam, 2012) signaling pathways have been determined to be associated with drug resistance. As a result, only the target genes were chosen that were present in these pathways (Fig. 3). Moreover, we found the gene targets that their interaction was detected biologically by luciferase assays in previous studies (Table 2 and Fig. 4A).
Confirmed Targets of Four Selected miRNAs in PI3K-AKT, MAPK, and foxO Signaling Pathways
miR-195-5p as a key regulator of gene expression plays significant roles in proliferation, stress response, and metabolism. In this study, we assessed both the expression of miR-195-5p in BT-474 human breast cancer cells and its possible role in drug resistance. First, using qPCR, we found that the expression level of miR-195-5p in BT-474 resistant breast cancer cells was significantly lower than that for parental BT-474 cells. This suggests that the expression of miR-195-5p is related to the drug resistance of breast cancer cells and that may serve as a tumor suppressor. Marques et al. (2018) illustrated that miR-195 and let-7 were downregulated in the malignant breast cancer patient group compared with the benign and control groups. Thakur et al. evaluated miRNA expression in tissue and serum of breast cancer patients and compared them with normal samples. They detected that miR-195 was downregulated in tissue and serum of triple negative breast cancer patients (Thakur et al., 2016).
miR-195 was also reported to be downregulated in the circulation of a breast cancer animal model (Waters et al., 2012). As reported previously, upregulation of miR-195 expression has been inhibited cell proliferation and invasion by targeting the Ccnd1 and Raf1 in MCF7 and ZR-75-30 human breast cancer cells (Li et al., 2011). Once activated, Raf1 can phosphorylate to activate the Raf-MEK-ERK pathway that adjusts cell cycle, proliferation, apoptosis, and migration (McCubrey et al., 2007). Although numerous studies have revealed that Raf1 is overexpressed in different cancers, the mechanism of Raf1 regulated by miRNA is unclear. Vascular endothelial growth factor A (VEGFA) was predicted as a direct target of miR-195-5p and confirmed by a dual luciferase experiment. Upregulation of VEGFA directly adjusted signaling pathway that was involved in angiogenesis. Furthermore, myeloblastosis (MYB) is regarded as an oncogene in a number of human tumors (Fang et al., 2009). Dysregulation of MYB activates several key gene targets, such as cyclooxygenase-2 (COX-2), Bcl-2, and c-Myc to promote tumor development (Biroccio et al., 2001), which can play significant roles in diverse processes, including proliferation, invasion, and migration.
Another important confirmed target of miR-195 is AKT3 that previously reported to inhibit the proliferative, migrated, and invasive properties of GC cells through PI3K-AKT signaling pathway. Recently, emerging evidence shows that miR-195 can increase the chemotherapy sensitivity in different cancers. For instance, it has been described that miR-195 has a positive effect on the chemotherapy sensitivity of doxorubicin in colon cancer cells by targeting BCL2L2 mRNA (Qu et al., 2015). Also, another study found that the overexpression of miR-195-5p enhanced chemotherapy sensitivity to 5-FU and L-OHP by targeting P-gp, BCL-2, and MRP1 in GC cells (Nie et al., 2018).
Our results revealed that the expression of miR-340 was specifically higher in parental HER2-positive BT-474 cells, indicating that miR-340 target might be an important molecular mediator of drug resistance. Chen et al. (2016) and Wu et al. (2011) in different studies showed that miR-340 might play a significant role in suppressing breast cancer invasion and metastasis. An important study showed microarray-based analysis of miRNA expression profile in breast cell lines with a various potential for invasion and observed that miR-340 was significantly decreased in breast cancer cell lines with high invasive capacity (Wu et al., 2011). Using in silico analysis, Korn et al. (2002) demonstrated that miR-340 was predicted to target several genes that were defined to be upregulated in breast cancer samples after anthracycline-based chemotherapy. Several recent studies suggest that miR-340 is downregulated in many tumors. The cancer-related downregulation of miR-340 appeared from the miRNA expression profile of the NCI-60 human cancer cell lines (Fernandez et al., 2015). miR-340 inhibited c-Met expression through direct targeting of the c-Met gene and inhibited c-Met expression. The c-Met is an oncogenic cell surface receptor tyrosine kinase, which is overexpressed in a variety of tumors, including breast cancers (Wu et al., 2011). Shattuck et al. (2008) identified Met as a receptor tyrosine kinase that was often co-expressed with HER2 in HER2-positive breast cancer and contributed to trastuzumab resistance of HER2-overexpressing breast cancer cells by the activation of AKT Serine/Threonine Kinase (AKT).
miR-23b-3p roles have been reported in various types of cancer. miR-23b role in the pathogenesis of breast cancer is still debated (Gong et al., 2011); some reports support the role of miR-23b as a tumor suppressor, but others offer the oncogenic function that depends on cellular model (Donadelli et al., 2014). Clinical analysis showed that miR-23b has oncogenic roles in breast cancer and associated with breast cancer prognosis (Jin et al., 2013). Another study showed that upregulation of miR-23b in primary tumors, compared with their corresponding lymph-node metastases, has an important role in the enhancement of invasion (Pellegrino et al., 2013). In human colon cancer, miR-23b-3p downregulates and regulates MAP3k1 to mediate in vivo metastasis (Zhang et al., 2011). In renal cell carcinoma, miR-23b-3p could serve as an oncogenic miRNA and inhibited PTEN (Zaman et al., 2012).
miR-656 belonging to the miR-379/miR-656 miRNA cluster is one of the largest known clusters of miRNAs in vertebrates (Glazov et al., 2008). Several mature miRNAs of this cluster have been experimentally found and studied (Khan et al., 2013), but the studies of miR-656 expression in cancer cells have just been reported. There is a lack of functional study on this specific miRNA in human cell line models and clinical samples. In this study, we demonstrated, for the first time, that miR-656 was statistically significant downregulated in a trastuzumab-resistant breast cancer cell. In silico analysis of another HER2-positive breast cancer cell line, SKBR-3 (GEO accession no. GSE47011), showed that miR-656 was significantly downregulated in SKBR-3 trastuzumab-resistant HER2-positive breast cancer cell line, which agreed well with our findings. We assumed that CREB1 might be potential targets of miR-656. Studies showed that CAMP Responsive Element Binding (CREB) Protein increases the expression of several target genes, involving in various cell functions such as metabolism, cell cycle, survival, and DNA repair. These functions showed that CREB was a critical factor for the growth, survival, and migration (Kew et al., 2014). Therefore, for the first time, we postulate that miR-656 may involve in trastuzumab resistance through targeting CREB1 transcripts.
Conclusions
In conclusion, we identified four miRNAs that might play important roles in trastuzumab resistance of breast cancer. To the best of our knowledge, this is the first study suggesting that HER2-positive breast cancer trastuzumab resistance might be associated with dysregulation of miR-23b-3p, miR-195-5p, miR-656-5p, and miR-340-5p. We assumed that thoughtful changes in expression status of dysregulated miRNAs might be a potential therapeutic option to improve adjuvant trastuzumab therapy in HER2-positive breast cancer. In this study, bioinformatic approaches revealed that the mentioned dysregulated miRNAs are strongly associated with trastuzumab resistance pathways; however, we suggest further experimental studies to confirm our results.
Footnotes
Acknowledgments
This work was financially supported by the University of Sistan and Baluchestan, Zahedan, Iran. The authors give their appreciation to the Birjand University of Medical Sciences for the constructive support on this article.
Disclosure Statement
No competing financial interests exist.