
Abstract
Asthma exacerbation is induced by the interaction of genes and environmental factors such as cigarette smoke. NLRP4 counteracts the activity of the inflammasome, which is responsible for asthma exacerbation. In this study, we analyzed the association of single-nucleotide polymorphisms of NLRP4 with the annual rate of exacerbation and evaluated the additive effect of smoking in 1454 asthmatics. Asthmatics possessing the minor allele of rs1696718G > A had more frequent exacerbation episodes than those homozygous for the common allele (0.59 vs. 0.36/year) and the association was present only in current and ex-smokers. There was a significant interaction between the amount smoked and rs16986718 genotypes (p = 0.014) and a positive correlation between the number of annual exacerbation episodes and amount smoked only in rs16986718G > A AA homozygotes. The prevalence of frequent exacerbators (≥2 exacerbation episodes/year) was 2.5 times higher in rs16986718G > A minor allele homozygotes than in common allele homozygotes (12.0% vs. 5.9%). Furthermore, the prevalence was 6 times higher in rs16986718G > A minor allele homozygotes who were current and ex-smokers than in nonsmokers (25.6% vs. 4.1%). The minor allele of rs16986718G > A in NLRP4 may be a genetic marker that predicts asthma exacerbation in adult asthmatics who smoke.
Introduction
A
Identified genetic variants that cause exacerbation include rs1800925 in IL13 (Hunninghake et al., 2007) and rs7216389 in ORMDL3 in children (Bisgaard et al., 2009), rs4950928 in CHI3L1 in children and young adults (Cunningham et al., 2011), and two nonsynonymous coding single-nucleotide polymorphisms (SNPs) (rs1805011 [E375A] and rs1801275 [Q551R]) in IL4R in adult asthmatics (Wenzel et al., 2007). In addition to the well-known asthma genes, GSDMB, IL33, RAD50, IL1RL1, CTNNA3, and SEMA3D, in childhood asthmatics (Bonnelykke et al., 2014; McGeachie et al., 2015), recent genome-wide association studies (GWAS) have also identified CDHR3 (encoding cadherin-related family member 3) and CMTR1 in children and adults and TRIM24 and MAGI2 in adult asthmatics as susceptible genes (Dahlin et al., 2015).
Environmental and social factors predisposing asthmatics to exacerbation include allergenic or occupational sensitization, exposure to mold, viral infection, heavy exercise, tobacco smoke, and some medications (Saxon and Saxon, 2005; Bloomberg, 2011). Immune responses to these factors induce activation of the inflammasome. Recognition of viral RNA by a set of cytosolic sensors induces production of type I interferons (IFNs) and the assembly of inflammasome complexes that activate caspase-1 (Kanneganti, 2010). The inflammasome consists of nucleotide-binding oligomerization domain-like receptor (NLR) family members containing the pyrin domain (NLRP), the adaptor molecule apoptosis-associated speck-like protein containing a Caspase activation and recruitment domain, and caspase-1 (Schroder and Schroder, 2010; De Nardo et al., 2014).
Similarly, exposure to air pollutants and cigarette smoke elicits immune responses mediated by specific Toll-like receptor- and NLR-dependent mechanisms (Jiang et al., 2006; Bauer et al., 2012). In the latter, NLRP3 binds to procaspase-1 through ASC, subsequently activating caspase-1, which leads to maturation of proinflammatory cytokines such as interleukin (IL)-1β, IL-18, and IL-33 (Dinarello, 2009; De Nardo et al., 2014). By contrast, NLRP4 counteracts inflammasome activity by inhibiting tumor necrosis factor- and IL-1β-induced nuclear factor-κB activation (Fiorentino et al., 2002) and negatively regulates type I IFN signaling by serving as an adapter to promote the ubiquitination of activated TBK1 (Cui et al., 2012).
Consequently, downregulation of the function of NLRP4 might be responsible for the frequent exacerbation in asthmatics in specific environments, such as in smokers, in whom the number of cigarettes smoked is easily assessed. This prompted us to analyze the association of SNPs of NLRP4 with the annual rate of asthma exacerbation and to evaluate relationships of current and previous smoking with the genetic effect.
Materials and Methods
Study subjects
DNA from 1454 asthmatics (995 nonsmokers, 274 ex-smokers, and 225 smokers), who had been followed for more than 1 year since enrollment, was obtained from the biobank of Soonchunhyang University Bucheon Hospital, Korea. Informed consent was obtained from each study subject who donated DNA to the biobank. The subjects were all of Korean descent and met the criteria for asthma: a physician's diagnosis of asthma and the presence of reversible airway obstruction (short-acting bronchodilator-induced increase of forced expiratory volume in the first second [FEV1] >12% and 200 mL or variation of peak expiratory flow rate >20% over a 4-week period); improvement in FEV1 >20% after treatment with asthma medication for 2 weeks, including inhaled or systemic corticosteroids (SCSs); or airway hyper-reactivity (methacholine PC20 < 10mg/mL). FEV1, forced vital capacity (FVC), and the postbronchodilator FEV1 were measured at baseline and every 3 months. The number of exacerbation episodes in the first year after enrollment was counted. Exacerbation recovery was defined as in the American Thoracic Society/European Respiratory Society statement (Reddel et al., 2009)—as recovery from the exacerbation by increasing the dose of inhaled corticosteroid (ICS) or adding bronchodilators and antileukotrienes, or as recovery from the exacerbation by adding oral corticosteroids (OCSs; >0.5 mg/kg of prednisolone for more than 3 days), while considering hospitalization or an emergency room visit. The respective amounts of ICSs and OCSs are expressed as the equivalent dosage of fluticasone per day or prednisone per year used in the first year. A frequent exacerbator (n = 126) was defined as an asthmatic experiencing two or more exacerbation episodes over the first year and the other subjects were defined as nonfrequent exacerbators (n = 1328). The study protocol was approved by the Ethics Committee of Soonchunhyang Bucheon Hospital (SCHBC_2014_07_028).
Selection of SNPs and genotyping
SNPs in exons of NLRP4 were selected using the Asian population database from the International HapMap Project (
Statistics
Fisher's exact test was used to compare the observed numbers of each genotype with those expected for a population in Hardy–Weinberg equilibrium (HWE). Haplotypes were inferred using the expectation maximization algorithm in C (Stephens et al., 2001). A type III univariate general linear model was applied to continuous variables (number of exacerbation episodes) and multiple logistic regression to discrete variables (presence of frequent exacerbation). In the logistic regression analysis, odds ratios (ORs) and 95% confidence intervals were calculated for each genotype and haplotype. Correlation between the amount smoked and the annual number of exacerbation episodes was evaluated using Spearman's rho (ρ). To evaluate the interaction between genotype (or haplotype) and smoking, we performed a multiple regression analysis, including an interaction term (genotype × amount smoked). Data were analyzed using SAS, version 9.1 (SAS, Cary, NC), and SPSS, version 12.0 (SPSS, Chicago, IL). To correct p-values for multiple comparisons, the effective number of independent SNPs in NLRP4 was calculated using SNP spectral decomposition (
Results
Characteristics of study subjects
The study enrolled 1454 asthmatics: 126 frequent exacerbators and 1328 nonfrequent exacerbators (Table 1). The frequent exacerbator group was dominated by older asthmatics with a larger amount smoked, less atopy, and lower FEV1 and FVC compared with the nonfrequent exacerbator group (p < 0.05). The mean daily ICS and annual SCS dosages were significantly higher in the frequent exacerbator group than in the nonfrequent exacerbator group (p < 0.05). Therefore, age, amount smoked, atopy, FEV1% of the predicted value at baseline, and total ICS and SCS dosages were used as covariates in the analyses of genetic associations.
Data are expressed as mean ± standard error.
ES, ex-smoker; SM, current smoker.
FVC, forced vital capacity; ICS, inhaled corticosteroid.
Frequencies, heterozygosity, and HWE of SNPs of the NLRP4 gene
The HWE of the 16 SNPs was >0.05 (Supplementary Table S1; Supplementary Data available online at
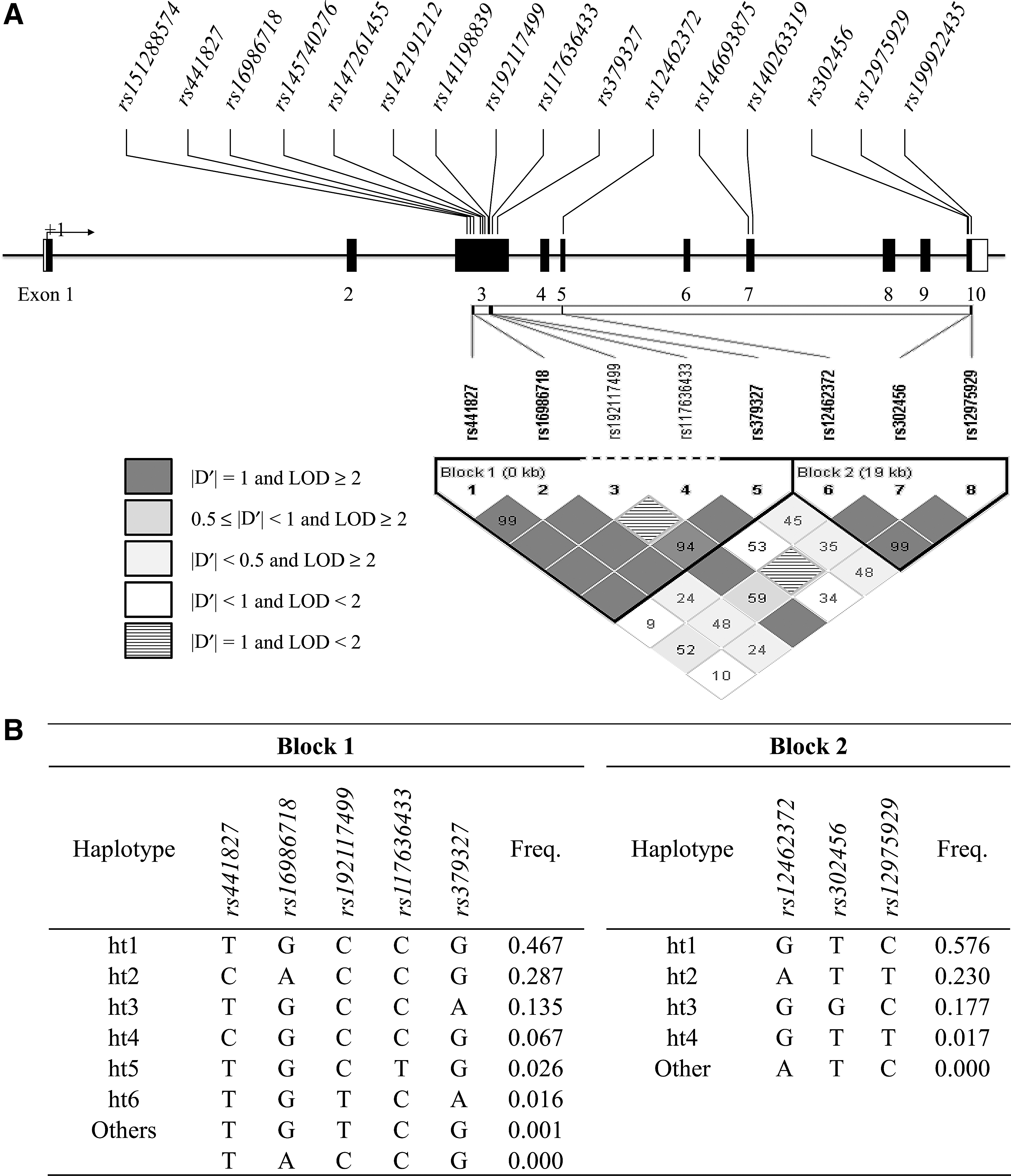
Association of SNPs of the NLRP4 gene with the annual number of exacerbation episodes
The eight SNPs of NLRP4 were analyzed for associations with the number of exacerbation episodes using a univariate general linear model. Minor allele homozygotes and heterozygotes of rs16986718G > A had significantly more annual exacerbation episodes compared with common allele homozygotes (mean ± standard error; 0.59 ± 0.13 vs. 0.59 ± 0.07 vs. 0.36 ± 0.04, p = 0.001, in dominant models, Table 2). The difference remained after correcting for multiple comparisons (p corr = 0.007). Rs441827 and ht1 in block 1 also showed an association with the number of exacerbation episodes (p = 0.022 in the dominant model), although these differences were not significant after correcting for multiple comparisons (Table 2).
Adjusted for age, smoking amount, atopy, predicted FEV1% at the first visit, and total ICS and SCS doses in the first year.
CC, common allele homozygote; CR, heterozygote; pcorr, corrected p value for multiple comparisons; RR, minor allele homozygote; SCS, systemic corticosteroid; SE, standard error of mean.
Effect of smoking on the genetic association of NLRP4 with exacerbation
Because acute exacerbation is related to smoking (Dougherty and Fahy, 2009; Polosa and Thomson, 2013), we compared the association of the eight SNPs and haplotypes of NLRP4 with exacerbation between the current or ex-smoker (SM) and never-smoker (NS) groups. The association of rs16986718 with the annual number of exacerbation episodes was more robust in the SM group. The minor allele of rs16986718G > A showed susceptibility to developing exacerbation with a good gene–dose effect (1.12 ± 0.31 for minor allele homozygotes, 0.64 ± 0.11 for heterozygotes, and 0.39 ± 0.06 for common allele homozygotes) (p = 0.004, p corr = 0.030, in the dominant model, Table 3). By contrast, this genetic effect of rs16986718 disappeared in the NS group (Supplementary Table S2).
Adjusted for age, smoking amount, atopy, predicted FEV1% at the first visit, and total ICS and SCS doses in the first year.
When the interaction term was applied to the association analysis, the interaction between the amount smoked and the rs16986718 genotypes was significant (F = 1.572, p = 0.014). There was also a positive correlation between the number of exacerbation episodes and amount smoked in asthmatics who were rs16986718 AA homozygotes (ρ = 0.326; p = 0.00034), but not in those possessing the G allele (ρ = 0.054; p = 0.141 in GG homozygotes and ρ = 0.066; p = 0.107 in GA heterozygotes, Fig. 2).
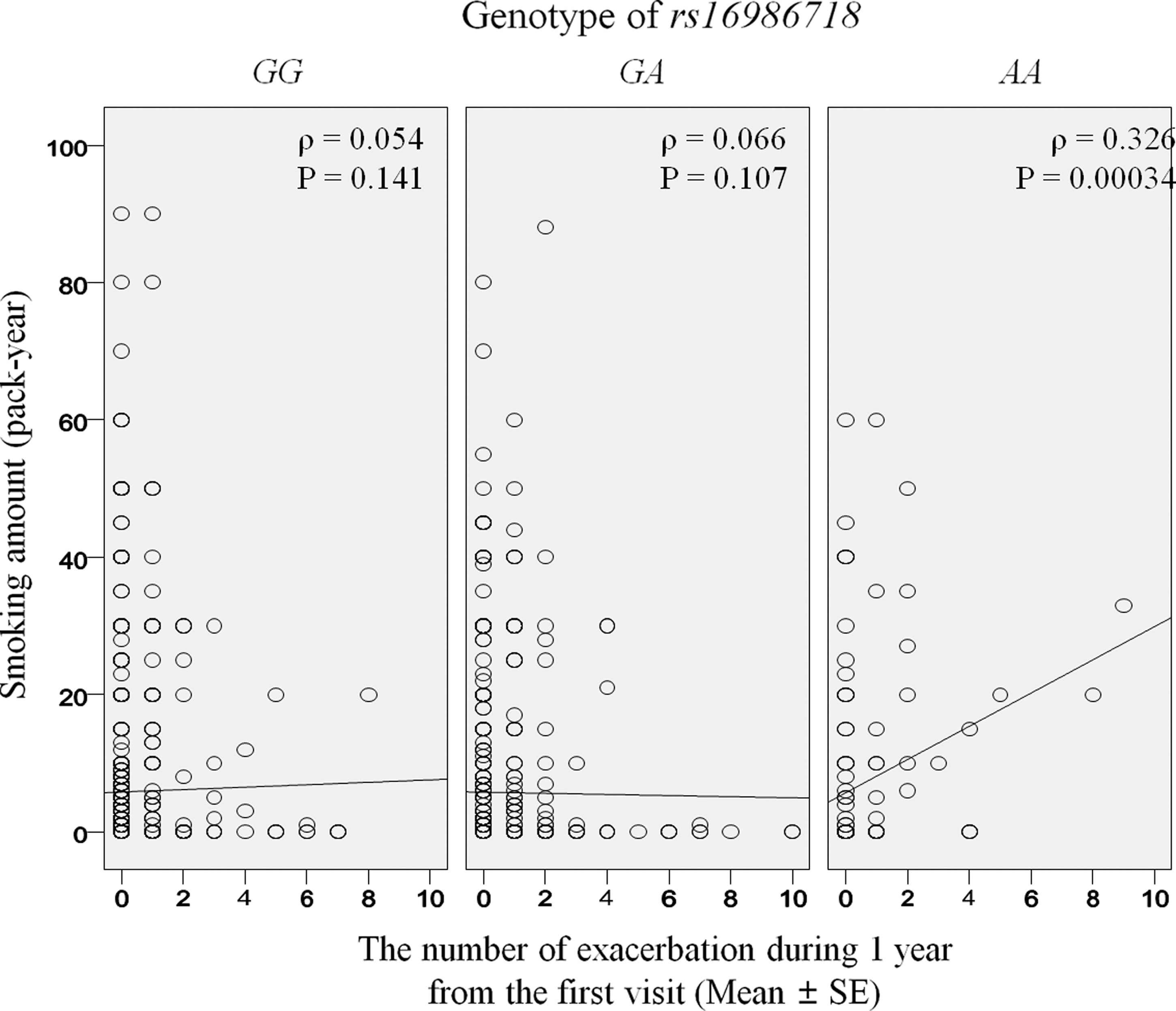
Correlation between smoking amount and the number of exacerbation episodes for 1 year in asthmatics according to the genotype of rs16986718. The correlation was evaluated using Spearman's Rho (ρ test).
Association of frequent exacerbators with SNP genotypes in the NLRP4 gene
The eight SNPs were analyzed for associations with the risk of frequent exacerbation using logistic regression analysis. The frequency of asthmatics who were rs16986718 G > A AA homozygotes and GA heterozygotes was significantly higher in the frequent exacerbator group compared with nonfrequent exacerbators (65.3% vs. 47.9%, p = 0.000067, OR = 2.56, Supplementary Table S3). The difference remained after correcting for multiple comparisons (p = 0.00047 in the dominant model).
The association of SNP rs16986718 G > A with the risk of frequent exacerbation became more apparent in the SM group: the frequency of minor allele homozygotes of rs16986718 G > A was three times higher in frequent exacerbators than in nonfrequent exacerbators (22% vs. 7.2%; OR = 3.52, p = 0.001, in the dominant model, Supplementary Table S4). However, this genetic effect of rs16986718 on the risk of frequent exacerbation disappeared in the NS group (Supplementary Table S5).
Comparison of the prevalence of frequent exacerbators among genotypes of rs16986718 G > A in the NLRP4 gene
The overall prevalence of frequent exacerbators was two times higher in those carrying the minor allele rs16986718G > A than in those carrying the common allele (12% in rs16986718AA, 11% in rs16986718GA, and 5.9% in rs16986718GG, OR = 1.84 [1.33–2.55], p = 0.000205, p corr = 0.0014, Fig. 3). The genotype-dependent difference in the prevalence of frequent exacerbators became more robust in the SM group: the prevalence of frequent exacerbators was four times higher in those carrying minor allele homozygotes of rs16986718G > A than in those carrying common allele homozygotes (25.6% in rs16986718AA vs. 6.3% in rs16986718GG, OR = 2.44 [1.53–3.9], p = 0.00018, p corr = 0.0013, Fig. 3). By contrast, the prevalence of frequent exacerbators did not differ between the rs16986718GA genotypes in the NS group.
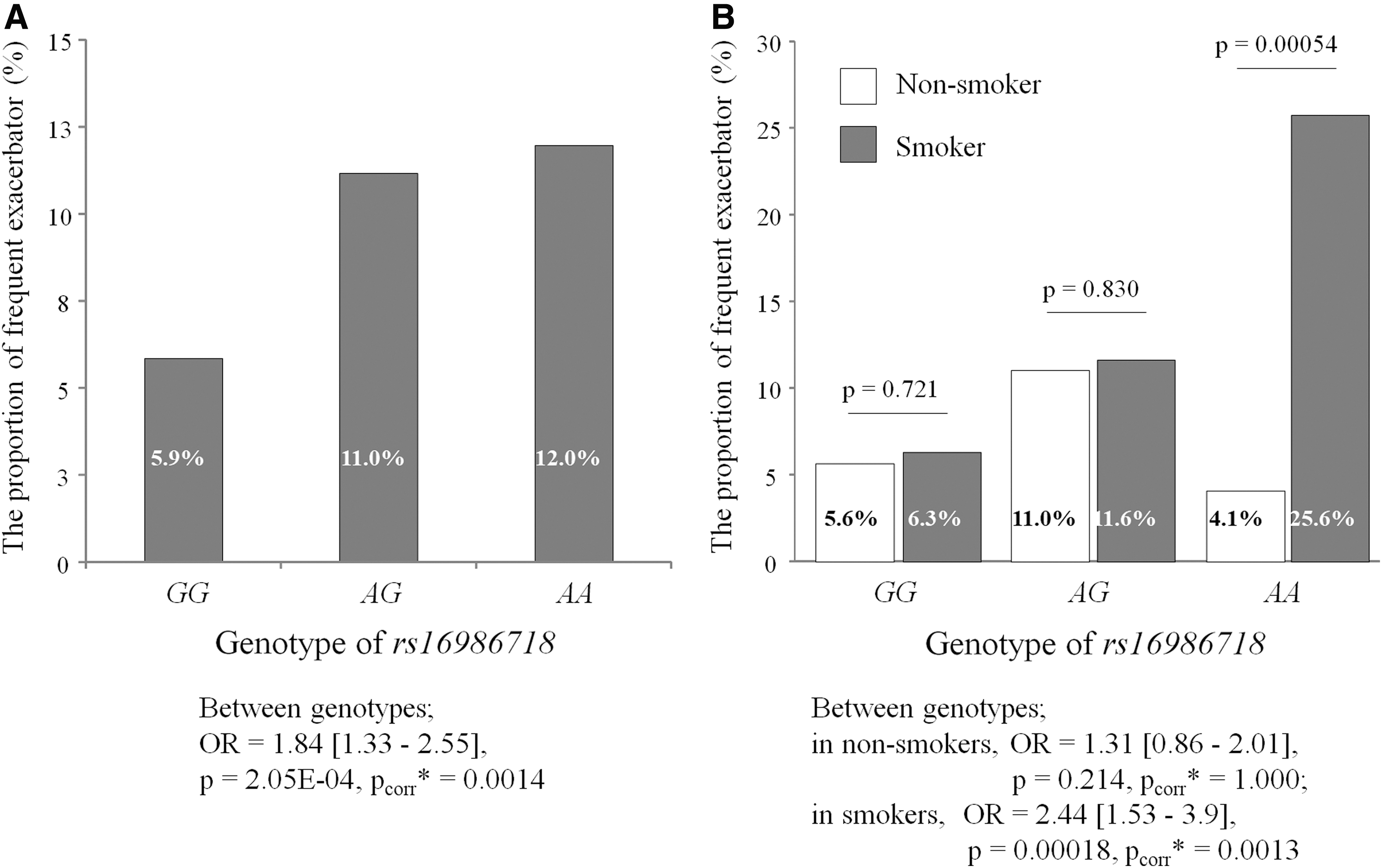
Proportion of frequent exacerbators (≥2 in 1 year) among indicated genotypes of rs16986718 in total study subjects (n = 1454)
When comparing SM and NS groups, the prevalence of frequent exacerbators was six times higher in the minor allele homozygotes of rs16986718GA in the SM group than in the NS group (25.6% vs. 4.1%, p = 0.00054, Fig. 3). By contrast, there was no difference in the prevalence of frequent exacerbators in the common allele homozygotes and heterozygotes of rs16986718GA between the SM and NS groups.
Discussion
To our knowledge, this is the first study to demonstrate that the minor allele of rs16986718 G > A in NLRP4 was strongly associated with frequent exacerbation in adult asthmatics and that smoking had a decisive effect on the occurrence of asthmatics with the susceptible SNP. The smoking effect seems to be dependent on the amount smoked by subjects carrying the minor allele of rs16986718: the amount smoked was related to the number of acute exacerbation episodes only in minor allele homozygotes of rs16986718 G > A. Furthermore, even in minor allele homozygotes of rs16986718GA, smoking robustly increased the prevalence of frequent exacerbators. Based on these results, we suggest that the genetic effect of the rs16986718 G > A minor allele on asthma exacerbation is exerted under the complete influence of smoking in adult asthmatics.
Although it is still not known how the genetic component and smoking exert their interactive effect on exacerbation of asthma, the smoking-induced attenuation of the defense mechanism against respiratory viral infections is a plausible explanation. Of all asthma exacerbation episodes, 50% in adult asthmatics and ∼80% in childhood asthmatics are induced by respiratory viral infections (Dougherty and Fahy, 2009; Kim et al., 2018). For episodes of exacerbation, rhinovirus is the virus most frequently detected in adults, followed by influenza A, parainfluenza, respiratory syncytial virus, and adenovirus (Seo et al., 2017).
One of the mechanisms underlying the susceptibility to viral infection in asthmatics is the change in epithelial function, including an increase in intracellular adhesion molecule-1, disruption of epithelial barrier function, impaired apoptosis, increased cell lysis, and deficient Th1 response involving IFNs (Wark and Gibson, 2006). The airway epithelial cell responses to type II IFN are critical for regulating the defense against these respiratory viral infections.
Interestingly, cigarette smoke extracts decrease the inhibitory effect of IFN-γ on proliferation of viruses such as respiratory syncytial virus (Modestou et al., 2010). Cigarette smoke extracts also suppress IFN-γ-dependent gene expression by airway epithelial cells (Modestou et al., 2010) and the antiviral effects of type I IFN through phosphorylation-dependent downregulation of the IFNAR1 subunit of type I IFN receptor (HuangFu et al., 2008). As the NLRP4 signalosome is tightly related through negative regulation of type I IFN signaling (Lin et al., 2016), marked attenuation of the type I IFN pathway is expected in smoking asthmatics with the A allele of rs16986718G > A, although this has not been validated because it was not feasible to obtain respiratory epithelium from the study subjects.
The effect of smoking on asthma exacerbation was observed only in about 25% of subjects carrying the minor allele of rs16986718 G > A. This indicates that other genetic and environmental factors contribute to inducing frequent exacerbators. A recent review found that acute exacerbation is associated with SNPs of several candidate genes, including IL13, IL4R, CHI3L1, and ORMDL3 (Park and Tantisira, 2017). Using GWAS, SNPs of CDHR3, CTNNA3, and SEMA3D have been identified as new risk genetic components.
More importantly, gene–environment interactions seem to be pivotal in asthma exacerbation. Rs512625 in ADAM33 located on chromosome 20p13 showed significant interactive effects with environmental tobacco smoke on severe exacerbation (Bukvic et al., 2013). In addition, exposure to house dust mites significantly modifies the relationship of SNPs (rs2486953, rs4950936, and rs1417149) of CHIT1 with severe exacerbation (Wu et al., 2010). Two genetic variants (rs2915863 in CD14 and rs17226566 in LY96) showed different relationships between endotoxin exposure and severe exacerbation (Kljaic-Bukvic et al., 2014). However, most of these studies examined childhood asthmatics, so interactive effects between genetic factors and indoor environment exposure in adult asthmatics, as in the present study, need to be investigated further.
Our previous cluster analysis study demonstrated two types of exacerbation-prone asthma in nonsmoking asthmatics: late-onset nonatopic asthma with impaired lung function and early-onset atopic asthma with severely impaired lung function (Kim et al., 2017). In our study, the prevalence of atopy was comparable between SM and NS groups. Therefore, atopy may not affect the interaction of smoking and rs16986718 G > A with the risk of frequent exacerbation.
This study has several limitations. First, the SNPs evaluated were located in exons. Especially because rs16986718 is a synonymous coding SNP (Thr185Thr), the allelic difference of this SNP is unlikely to affect the primary structure of NLRP4. However, silent SNP codons have been reported to affect gene function such as substrate specificity, which may be caused by changes in the timing of cotranslational folding when frequent codons are replaced by rare codons in a cluster of infrequently used codons (Kimchi-Sarfaty et al., 2007; Komar, 2007). Rs16986718 changes the 185th codon ACG to ACA in the NACHT domain of the NLRP4 protein, which may be important in regulating NLRP4 function. The original codon (ACG) is 2.38 times more frequent than the altered codon (ACA) in the human genome, and there are 10 human tRNA genes for the ACG codon, but only seven for the ACA codon (Lander et al., 2001). However, rs16986718 is not a site for transcription factor binding, splicing, splicing regulation, or microRNA molecular functions, according to a functional evaluation of SNPs in Asian populations (SNPinfo Web Server,
Conclusions
The NLRP4 genetic polymorphism was significantly associated with frequent acute exacerbation through an interaction with smoking status in adult asthmatics. The minor (A) allele of rs16986718 may be a risk factor for the occurrence of acute exacerbation in smokers. Our observations suggest that NLRP4 plays a role in the smoking-induced pathogenesis of asthma exacerbation and that genetic polymorphism is a promising marker for predicting frequent asthma exacerbation while considering the smoking behavior of asthmatic individuals.
Footnotes
Acknowledgments
The study was supported by a research grant from Soonchunhyang University to H.S.C. and by a grant from the Korean Health technology Research and Development project, Ministry of Health and Welfare, Republic of Korea (2016-ER7402-00). Clinical data and DNA were provided by a biobank of Soonchunhyang University Bucheon Hospital.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.