
Abstract
The Editor-in-Chief of DNA and Cell Biology officially retracts the article entitled, “miR-142 Suppresses Endometrial Cancer Proliferation In Vitro and In Vivo by Targeting Cyclin D1” by Su Y, Wang J, Ma Z, Gong W, Yu L. (DNA Cell Biol. 2019;38(2):144–150; 10.1089/dna.2018.4441) due to apparent duplication of images published in another journal1 and after receiving a report by the PRIMeR group out of the University of Sydney (Australia) led by Professor Jennifer Byrne, identifying that the ACTB forward and reverse primers provided under the section quantitative real-time PCR are primers targeting ACTB within Rattus rattus, not humans. Within the human transcriptome, they would be expected to also target two separate genes (forward: ACTG1, reverse: ACTC1). The Editor agrees with the report.
The authors were contacted via email of the reported issues utilizing the email addresses provided to the journal at the time of submission in 2018, though no response was received.
The Editor and Publisher of DNA and Cell Biology are committed to preserving the veracity of scientific literature and the community it serves.
1. Ono C, Fukuhara T, Motooka D, et al. Characterization of miR-122-independent propagation of HCV. PLoS Pathog. Epub May 11, 2017;
Introduction
Endometrial cancer (EC), arising from the endometrium, is one of the most frequent female pelvic tumors and is a major threat to women, with >189,000 new cases and ∼45,000 mortalities reported every year (Jemal et al., 2011; Tsikouras et al., 2013; Wan et al., 2018). Moreover, EC incidence in China is rising (Chen et al., 2016a). EC patients at early stage can be treated with surgery, and their prognoses are often good (Liu et al., 2018). However, the prognosis of EC patients with regional or distant metastasis is poor (Banno et al., 2013a). Therefore, it is imperative to explore novel methods for EC treatment. The study on the underlying mechanisms of EC progression can help with the identification of promising therapeutic targets and prognostic markers.
miRNAs, consisting of ∼22 nucleotides, can bind with the 3′-untranslated regions (3′UTRs) of target mRNAs and subsequently modulate gene expression (Bartel, 2004; Bhardwaj et al., 2010; Banno et al., 2013b). Aberrant expression of miRNAs is proved to be closely associated with tumorigenesis, development, and metastasis of various cancers (Wilczynski et al., 2016). Moreover, increasing evidence proves that mature miRNAs take part in many cellular processes, such as proliferation, apoptosis, and so on (Banno et al., 2013b). According to previous studies, some miRNAs may behave as oncogenes or tumor suppressors in EC. For instance, miR-139 targeted HOXA10 and reduced the growth and migration of EC cells (Liu et al., 2018). miR-145 significantly restricted EC cell proliferation and metastasis by targeting SOX11 (Chang et al., 2017). However, miR-10b fostered the propagation and invasion of EC cells through the downregulation of HOXB3 (Chen et al., 2016b). miR-130b overexpression enhanced the invasive ability of EC cells (Li et al., 2013).
Emerging reports show that miR-142 has anti-tumor activities against different types of malignancies. For example, Isobe et al. (2014) demonstrated that miR-142 negatively modulated the properties of breast tumor stem cells. Hua et al. (2018) showed that miR-142-3p significantly suppressed hepatocellular carcinoma (HCC) proliferation. In contrast, Islam et al. (2018) proved the oncogenic effects of miR-142 on colorectal cancer. Nevertheless, miR-142's roles in EC development are yet to be explored.
Here, the aim of the present study is to elucidate the effects of miR-142 on human EC pathogenesis. We provide evidence that miR-142 was downregulated in tumor tissues from EC patients compared with adjacent normal tissues. In addition, miR-142 expression in EC patients without metastasis was higher than that in patients with metastatic EC. Moreover, Kaplan–Meier assay suggested that low-level miR-142 may be closely linked with the poor clinical outcome of EC patients. Thus, we speculated that miR-142 may have anti-tumor effects on EC. To our delight, we finally found that miR-142 functioned as a tumor suppressor in EC development by targeting cyclin D1 (CCND1).
Materials and Methods
Ethical considerations
This study was approved by the institutional review board of the Affiliated Yantai Yuhuangding Hospital of Qingdao University (#037AC5). The frozen samples were harvested from 49 patients who received hysterectomy, and informed consent was obtained from all participants. The clinical data of patients are listed in Supplementary Table S1 (Supplementary Data are available online at
Cell culture and transfection
Ishikawa (ISK) and HEC-1A cells, obtained from ATCC, were kept in DMEM/F12 medium (Gibco, Grand Island, NY) containing 10% FBS (Gibco). miR-142 mimics and negative control (NC) were obtained from Sangon Biotech (Shanghai, China). Cell transfection was performed with Lipofectamine 2000 (Invitrogen, Waltham, MA).
MTT assay
Five thousand ISK or HEC-1A cells per well were seeded in 24-well plates, and MTT assay was conducted as described previously (Li et al., 2013).
Colony formation assay
Five hundred transfected EC cells in 0.35% agar were seeded on top of 0.6% agar layer. After 10 days, colonies were fixed and stained. The number of colonies were then counted. Visible colonies containing no less than 50 cells were counted under a light microscope (Olympus BX41TF, Tokyo, Japan).
Cell counting assay
ISK or HEC-1A cells were seeded at a density of 2 × 104 cells after transfection. At day 7, cells were resuspended in 0.4‰ trypan blue solution, and viable cells were counted.
Luciferase reporter assay
CCND1 3′UTR harboring wide-type or mutant miR-142-binding site was amplified and inserted into psi-check2 (Promega, Madison, WI) to generate CCND1 or CCND1-MUT, respectively. ISK cells were co-transfected with miR-142 mimics or NC, as well as psi-check2 or CCND1 or CCND1-MUT. At 48 h, luciferase activity was evaluated.
Quantitative real-time PCR
Total cellular RNA was extracted with TRIzol (Invitrogen), and cDNA Reverse Transcription kit (Takara, Dalian, China) helped with first-strand cDNA synthesis. SYBR green quantitative real-time PCR (qPCR) assay (Takara) was employed to perform qPCR. Data were analyzed using the 2–ΔΔct calculation method. Primers used are listed below: miR-142-fwd: 5′-GTCACCTGTAGTGTTTCCTACTT-3′ miR-142-rev: 5′-TATGGTTGTTCTGCTCTCTGTCTC-3′ CCND1-fwd: 5′-GCTGCGAAGTGGAAACCATC-3′ CCND1-rev: 5′-CCTCCTTCTGCACACATTTGAA-3′ β-actin-fwd: 5′-GTGGACAGTGAGGCCAGGAT-3′ β-actin-rev: 5′-GATTACTGCTCTGGCTCCTAGCA-3′
Western blot
Total protein was isolated from EC cells, subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis, and then transferred to polyvinylidene fluoride or polyvinylidene difluoride membrane. After blocking, blots were incubated with primary antibody against CCND1 (Abcam, Cambridge, MA). Subsequently, the membranes were probed with secondary antibodies, and all bands were assessed with an enhanced chemiluminescent detection system.
ISK cell xenografts in mice
ISK cells were transfected with miR-142 mimics or NC and named ISK-Control or ISK-miR-142 cells. Six-week-old male athymic nude mice were inoculated subcutaneously with 1 × 106 ISK-Control or ISK-miR-142 cells. At day 7, sizes of subcutaneous tumor mass were evaluated every 3 days. At 1 month postinjection, animals were killed and tumors were weighed and used in further experiment. Tumor volume was estimated using the formula: length × width × height × 0.5236 (Massfelder et al., 2004).
Immunohistochemical staining
After fixing and embedding, tumor slices were probed with anti-Ki67 (Abcam) and anti-CCND1 (Abcam), followed by 1 h incubation with secondary antibody. All slides were examined by two technicians, who were blinded to samples, under a light microscope. The positively dyed cells were randomly counted in 12 fields (400 × magnification). The mean of eight fields was determined as the rate of Ki67 marker.
Statistical analysis
All values are presented as mean ± standard deviation. Analysis of variance (ANOVA) or Student's t-test was used for statistical analysis. Difference was regarded statistically significant at p < 0.05.
Results
Low-level miR-142 was correlated with poor prognosis of EC patients
qPCR was employed to test whether miR-142 was associated with EC progress, and the data showed that there was much lower mRNA level of miR-142 in tumor tissues of EC patients compared with matched normal tissues (Fig. 1A; p < 0.001). In addition, Figure 1B indicates that there was lower mRNA abundance of miR-142 in tumor samples in EC patients with recurrence than that in samples of EC patients without recurrence (p < 0.001). Besides, Kaplan–Meier analysis demonstrated that EC patients harboring low-level miR-142 had worse prognoses compared with EC patients bearing high-level miR-142 (Fig. 1C).
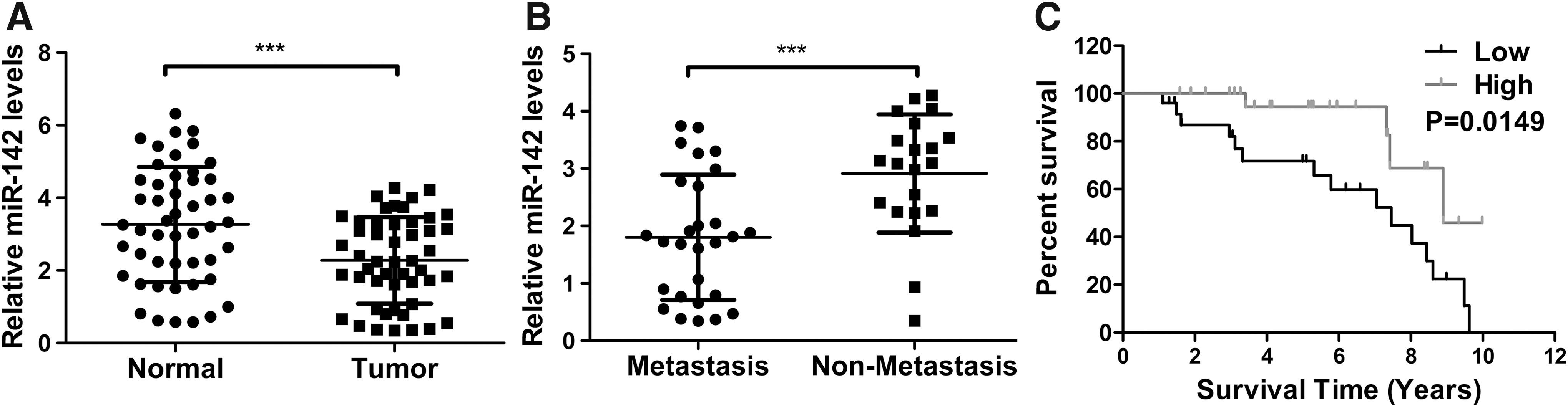
Low-level miR-142 was correlated with poor prognosis of EC patients.
miR-142 suppressed EC cell proliferation
To test the role of miR-142 on EC proliferation, ISK or HEC-1A cells were transfected with miR-142 mimics or NC. MTT assay revealed that miR-142 mimics time-dependently inhibited the viability of ISK or HEC-1A cells (Fig. 2A, B; p < 0.001). In addition, cell counting assay indicated that miR-142 inhibited ISK and HEC-1A cell proliferation to 66.67% (Fig. 2C; p < 0.05) and 71.42% (Fig. 2D; p < 0.01) in comparison with control, respectively. Besides, Figure 2E–H demonstrates that EC cells in miR-142 mimic groups had much more colonies compared with those in NC groups (p < 0.05 and <0.001 for ISK and HEC-1A cells, respectively).
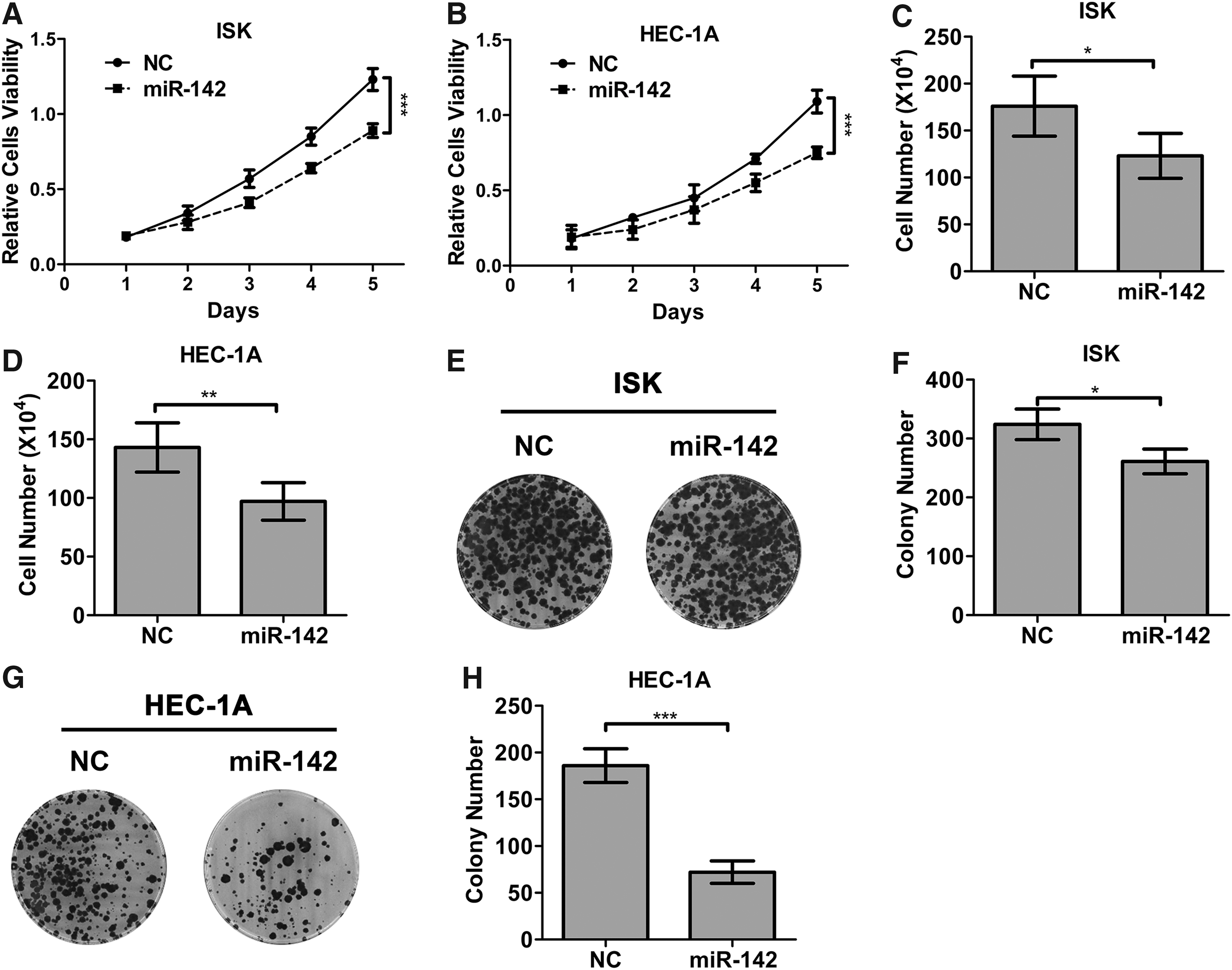
miR-142 suppressed EC cell proliferation.
CCND1 was a direct target of miR-142
Targetscan analysis predicted that there was putative miR-142 binding site in CCND1 3′UTR (Fig. 3A). Figure 3B and C proves that transfection of miR-142 mimics significantly inhibited CCND1 mRNA (p < 0.001) and protein expression in ISK cells compared with those in NC group, respectively. Moreover, Figure 3D indicates the negative relationship between miR-142 and CCND1 expression in EC patients. Besides, luciferase reporter assay proved that miR-142 mimic significantly decreased luciferase expression in ISK cell bearing wild-type CCND1 reporter (p < 0.001), and this inhibitory activity of miR-142 was blocked by mutating the miR-142 binding site in CCND1 3′UTR (Fig. 3E).

CCND1 was a direct target of miR-142.
High-level CCND1 was correlated with poor prognosis of EC patients
As shown by Figure 4A, there was significantly higher level of CCND1 mRNA in tumor samples from EC patients in comparison with matched normal tissues (p < 0.001). Moreover, Figure 4B shows that CCND1 mRNA abundance in tumor samples from EC patients with recurrence was much higher than that in samples of EC patients without recurrence (p < 0.05). In addition, Figure 4C suggests that EC patients bearing high-level CCND1 may have worse prognoses compared with those with low-level CCND1.
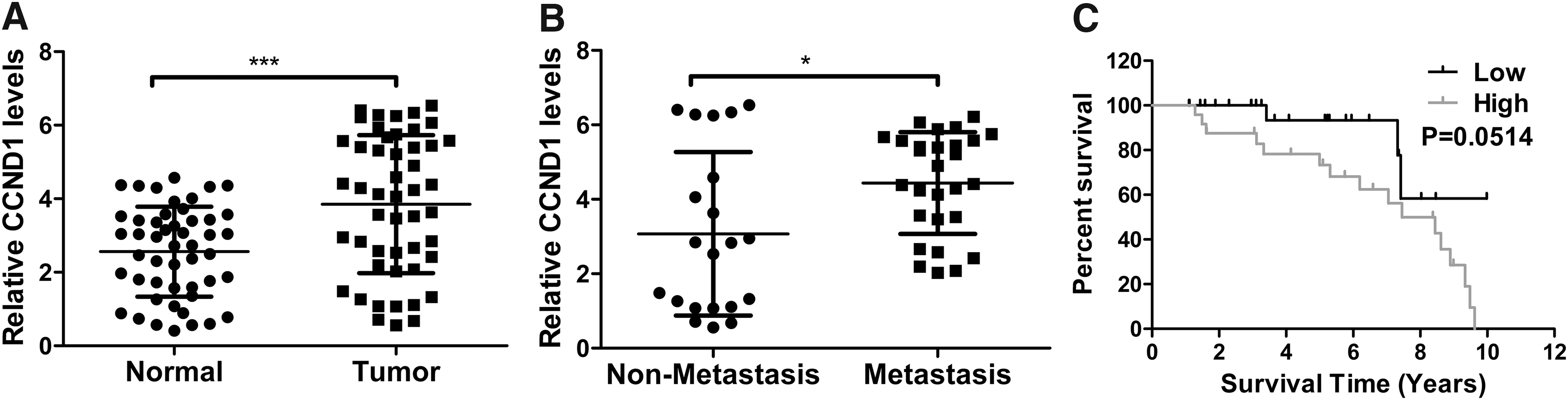
High-level CCND1 was correlated with poor prognosis of EC patients.
miR-142 suppressed tumor growth of EC in vivo
Figure 5A indicated that subcutaneous tumors in the miR-142 group grew much slower compared with control (p < 0.001). At 1 month, the average tumor weight in the miR-142 group was 149 ± 37 mg as against 300 ± 65 mg in the control group (Fig. 5B; p < 0.001). In addition, the average final mice weight in control group was much lower than that in miR-142 group (Fig. 5C; p < 0.01). Therefore, miR-142 overexpression time-dependently suppressed tumor growth. Besides, the expression level of Ki67 and CCND1 in excised tumors was assessed with immunohistochemical (IHC) staining, and the results proved that there were fewer Ki67+ cells (Fig. 5D, E; p < 0.01) and fewer CCND1+ cells (Fig. 5F) in miR-142 group compared with control.
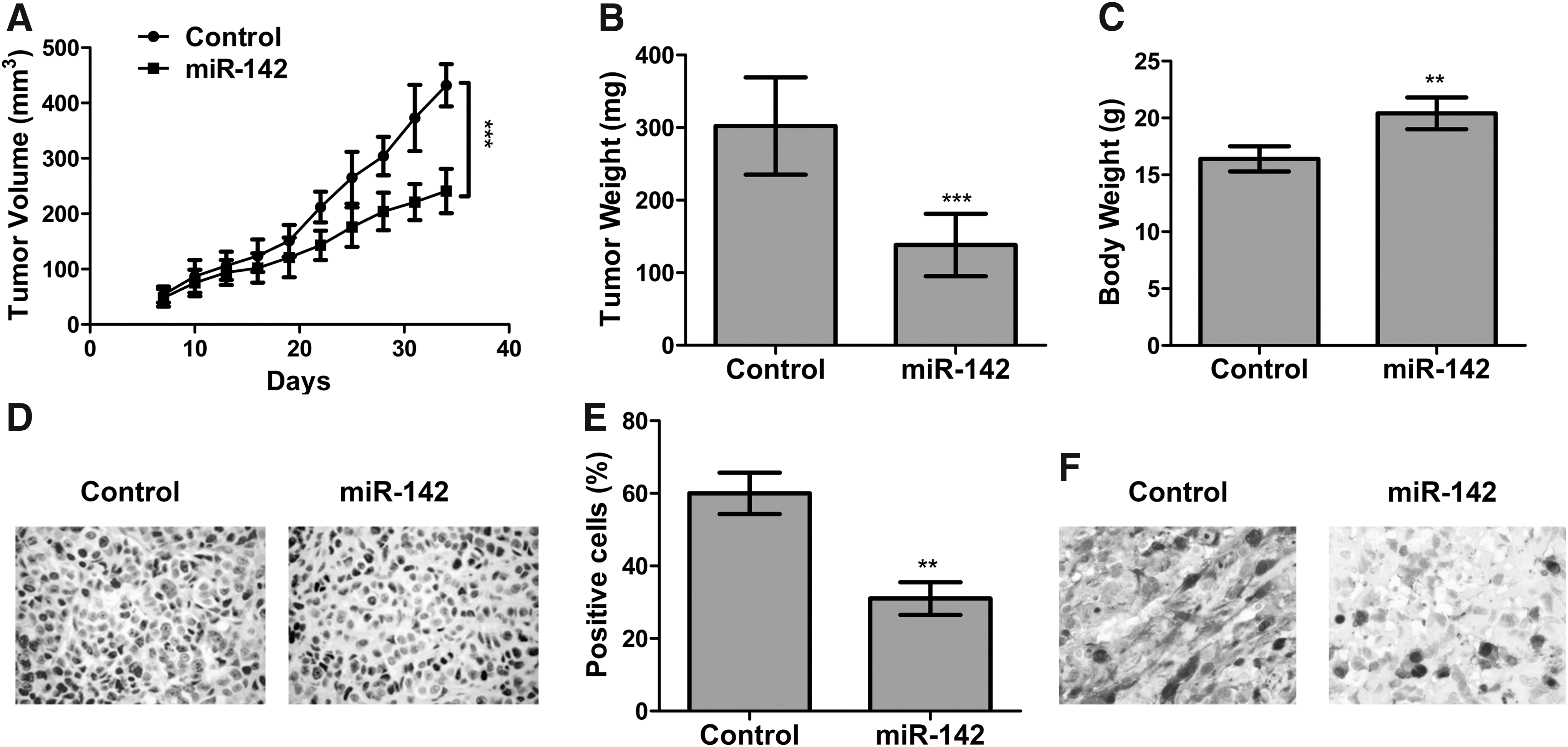
miR-142 suppressed tumor growth of EC in vivo.
Discussion
EC, caused by obesity, type II diabetes, excess estrogen, and some other factors (Widodo et al., 2016), afflicts patients and may lead to mortality. Generally speaking, there are two types of EC, including type I and type II (Chen et al., 2016c). Type I EC, also called estrogen-dependent EC, is the most frequent type of EC, and its initiation is linked with the long-term stimulation of estrogen (Chen et al., 2016c). However, type II EC is estrogen-independent, which accounts for 20–30% of cases (Lu et al., 2015). In addition, ISK and HEC-1A cells are known to exhibit typical characteristics of type I and type II EC, respectively (Treeck et al., 2003; Sun et al., 2006). Thus, both ISK and HEC-1A cells were employed in our study.
According to previous studies, many tumor-associated miRNAs play an important role in the initiation and progress of EC, such as miR-139, miR-145, miR-10b, and miR-130b (Li et al., 2013; Chen et al., 2016b; Chang et al., 2017; Liu et al., 2018). As for miR-142, it has been proved to act as a tumor suppressor in many tumors, such as breast cancer, HCC, and so on (Isobe et al., 2014; Hua et al., 2018). In contrast, miR-142 has oncogenic activities in other cancers, including renal cell carcinoma and colorectal cancer (Li et al., 2016a; Islam et al., 2018). Here, we found that miR-142 expression was downregulated in human EC tumor samples compared with adjacent normal tissue samples. In addition, miR-142 expression was also inhibited in metastatic EC tissues in comparison with non-metastatic controls. Moreover, EC patients with low-level miR-142 had worse prognoses than those bearing high-level miR-142. Hence, we made a hypothesis that miR-142 may have tumor-inhibitory activities in EC development. To test the above hypothesis, the proliferation of EC cells transfected with miR-142 mimics was evaluated with MTT, cell counting, and colony formation assay, and the results revealed that miR-142 suppressed the propagation of both ISK and HEC-1A cells.
It is well known that miRNAs often modulated the expression of multiple genes. Hence, it is necessary to identify the target of miR-142 in EC. To date, several targets of miR-142, such as LDHA in HCC (Hua et al., 2018), HMGA1 and FZD7 in breast cancer (Jia et al., 2018), and sirtuin 1 in ovarian cancer (Gao et al., 2018), have been characterized. With the help of targetscan analysis, we chose CCND1 as a candidate for the next experiment. To our delight, CCND1 was characterized as a miR-142's direct target by luciferase reporter analysis. According to Shen et al. (2013), miR-142-3p mimics downregulated CCND1 expression in colon cancer cells. Here, we found that miR-142 mimics significantly inhibited CCND1 mRNA and protein expression in ISK cells, which was consistent with the previous study by Shen et al. (2013). Moreover, miR-142 expression was negatively linked with CCND1 expression in tumor samples of EC patients. Taken together, miR-142 can target CCND1 and then regulate the expression of CCND1 in EC.
CCND1, a member of highly conserved cyclin family, can function as a modulator of cyclin-dependent kinase (CDK)4 or CDK6 and subsequently regulate G1/S transition (Baldin et al., 1993). Retinoblastoma, one of the tumor suppressors, can interact with CCND1 and positively modulate the expression of CCND1 (Li et al., 2016b). In addition, previous studies have indicated that CCND1 overexpression is often found in many types of human tumors and may be involved in tumorigenesis and tumor progression (Li et al., 2016b; Huang et al., 2018). For instance, Wang et al. (1994) have shown that CCND1 may have oncogenic effects on breast cancer. Huang et al. (2018) have proved that CCND1 expression was positively linked with the chemotherapy resistance of colorectal cancer. Dai et al. (2016) have revealed that the inhibition of CCND1 expression could block ovarian cancer cell proliferation and stimulate apoptosis. Moreover, increasing evidence shows that CCND1 has important roles in EC. According to Xu and Lin (2018), mutations in CCND1 c-terminus were enriched in EC samples, which may contribute to EC pathogenesis. Consistent with previous studies, we observed that CCND1 was enhanced in tumor samples of EC patients compared with matched normal tissues. In addition, CCND1 expression was also enhanced in tumor samples of patients with metastatic EC compared with tumor samples of patients with primary EC. Moreover, lower-level CCND1 was linked with better prognosis. Collectively, our data revealed that miR-142's anti-tumor roles were correlated with the downregulation of CCND1 expression.
Tumorigenicity analysis in nude mice was conducted to further investigate miR-142's effects on EC in vivo. The data showed that the overexpression of miR-142 significantly inhibited tumor growth in nude mice compare with control, as shown by a dramatical decrease in both volume and weight of subcutaneous tumors and a significant increase in the body weight of mice bearing ISK-miR-142 cells. Moreover, IHC staining proved that there was much fewer Ki67+ cells in tumor samples of mice inoculated with ISK-miR-142 cells than in control group. Ki-67 (also called MKI67), a nuclear protein, is an excellent marker of cell proliferation (Scholzen and Gerdes, 2000). Besides, compared with control group, tumor samples of mice inoculated with ISK-miR-142 cells had fewer CCND1+ cells. Thus, the above tests proved that miR-142 suppressed EC proliferation by downregulating CCND1 in vivo.
In summary, the above findings proved that that the downregulation of miR-142 and the upregulation of CCND1 in human EC tissues were closely linked with EC development and metastasis. Moreover, miR-142 exhibited an inhibitory role on EC proliferation by directly targeting CCND1 both in vitro and in mouse tumor model. Hence, EC patients bearing low-level miR-142, whose clinical outcomes were often poor, might be treated with miRNA-based methods, which enhanced the expression of miR-142. In addition, our results also suggested that CCND1 may be a potential marker of EC prognosis and a candidate therapeutic target.
Conclusions
miR-142 suppressed EC proliferation by targeting CCND1, which suggested the potential use of miR-142 as a therapeutic target of EC treatment.
Footnotes
Authors' Contributions
Performed the experiments and analyzed the data: Y.S., J.W., Z.M. Designed the study and wrote the article: W.G., L.Y. All authors approved the final submission.
Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.