
Abstract
Tumor necrosis factor alpha (TNF-α) appears to play an important role in proliferation and activation of hepatic stellate cells (HSCs), but it is unclear whether single nucleotide polymorphisms (SNPs) in the TNF-α gene influence HSCs function. In this study, we explored the effects of TNF-α A94T and P84L polymorphisms on the level of TNF-α, proliferation and activation of HSCs. It was found that A94T and P84L SNPs of TNF-α downregulated the mRNA and protein level of TNF-α in recombinant cells. Compared with wild-type TNF-α, A94T and P84L SNPs could decrease the growth or activation inhibitory effects of TNF-α on LX-2 cells, the human HSC line. In addition, A94T SNPs were associated with significantly lower expression of matrix-metalloproteinase 2 (MMP 2) or 9, but P84L SNP only decreased the mRNA level of MMP 9. A94T and P84L SNPs of TNF-α downregulated the level of IL-6. Furthermore, A94T and P84L SNPs decrease the activation inhibitory effects of TNF-α on LX-2 cells through inhibiting the phosphorylation levels of inhibitory kappa B-alpha (IκB-α) and P65. This study provides two vital SNPs for further functional or case–control studies of TNF-α SNPs.
Introduction
Liver fibrosis is characterized by produced deposition of extracellular matrix (ECM), is a compensatory response to chronic liver injury that could develop into liver cirrhosis, and even liver cancer except the removal of underlying etiology, including hepatitis B and hepatitis C virus infections, alcoholic hepatitis, and nonalcoholic steatohepatitis (NASH) (Arthur, 2002; Bataller and Brenner, 2005; Pellicoro et al., 2012). In fact, the key to the onset and progression of hepatic fibrosis is the proliferation and activation of hepatic stellate cells (HSCs), also known as Ito cells, fat-storing cells, or lipocytes (Pinzani, 1995; Pinzani and Maciasbarragan, 2010).
Several studies suggested that HSCs are targets for many cytokines. Platelet-derived growth factor, as the strongest mitogen, promotes the proliferation of HSCs (Andrae et al., 2008; Pinzani and Maciasbarragan, 2010). In addition, transforming growth factor beta 1 (TGF-βl), as the main fibrogenic cytokine, promotes HSCs proliferation, induces HSCs to transform into myofibroblast-like cells, and upregulates the expression of myofibroblast markers such as collagen type I, α-SMA, and so on (Dooley and Ten, 2012; Fabregat et al., 2016; Robert et al., 2016). Furthermore, tumor necrosis factor alpha (TNF-α), as the proinflammatory mediator, has been reported that it could inhibit HSCs proliferation (Gallois et al., 1998; Saile et al., 1999) and decrease the expression of collagen type I in vivo or in vitro (Buck et al., 1996; Knittel et al., 1997; Robert et al., 2016). Many studies (Iraburu et al., 2000; Varela-Rey et al., 2002, 2007; Novitskiy et al.,2005) have explored the underling molecular mechanism of downregulation of collagen type I by TNF-α. Novitskiy et al. (2005) and Varela-Rey et al. (2007) found that TNF-α could downregulate the transcription activity and product of collagen type I by inducing NF-κB pathway activation.
TNF-α is located in the major histocompatibility complex in which exists a high degree of polymorphism. It has been identified that many single nucleotide polymorphisms (SNPs) are located in the TNF-α promoter region, including −1031T/C, −863C/A, −857C/T, −575G/A, −376G/A, −308G/A, −244G/A, and −238G/A (Seki et al., 1999; Uglialoro et al., 1998; Bayley et al., 2004). Studies suggested that TNF-α SNPs alter the transcriptional activity (Kroeger et al., 1997; Wilson et al., 1997) and expression (Louis et al., 1998) level of TNF-α, and increased susceptibility to liver diseases including NASH (Wang et al., 2012), hepatitis B (Sghaier et al., 2015), hepatic fibrosis, and hepatocellular carcinoma (Jeng et al., 2007). Before case–control or functional studies, it is advisable to find out the vital functional SNPs. Dabhi B (Dabhi and Mistry, 2014) first found A94T (rs1800620) and P84L (rs4645843) of TNF-α by combining silico analysis with TNF-α dbSNP database using Rosanna's method (Squitti et al., 2014). They found that these SNPs could decrease the stability of TNF-α protein and further downregulate the expression of TNF-α. However, very few studies (Mortazavi et al., 2017) about these two SNPs are available to access.
Taking into account the crucial biologic role of TNF-α and HSCs in liver fibrosis, this study explores the effects of TNF-α A94T and P84L polymorphism on the mRNA and protein of TNF-α, proliferation, and activation of HSCs, and demonstrates the underlying molecular mechanisms.
Materials and Methods
Reagents
Cell Counting Kit-8 (CCK-8) was purchased from Biosharp (Wuhan, China). Enzyme-linked immunosorbent assay (ELISA) kits for TNF-α (EK0525), IL-6 (EK0410), and TGF-β1 (EK0513) were from Boster (Wuhan, China). Prime Script RT reagent kit was obtained from Takara (Japan). BCA protein assay kit was purchased from Thermo Fisher Scientific (Waltham). Primary antibodies against TNF-α (#3707), β-actin (#4970), and antirabbit secondary antibody IgG H + L (#5151) were purchased from Cell Signaling Technology (Danvers). Primary antibodies against P65 (ab16502), P-P65 (ab76302), IKB-α (ab32518), and P-IKB-α (ab133462) were obtained from Abcam (Cambridge, UK).
Plasmid constructs and cell culture
LX-2, the human HSC line, was obtained from Beijing North Carolina Souren Biotechnology (BNCC337957; Beijing, China). pIRES2-ZsGreen1 plasmids encoding TNF-α wild-type or the mutation A94T of TNF-α or the mutation P84L of TNF-α were obtained from Wuhan Biofavor Biotech Service Company (Wuhan, China). The TNF-α/TNF-α A94T/TNF-α P84L inserts were subcloned into pLVX-mCMV-ZsGreen-PGK-Puro, confirmed by sequencing, and transfected into LX-2 cells using the Lenti-Pac™ system. Stable recombinant LX-2 cells were selected using 1.5 μg/mL puromycin (Cayman Chemical, Ann Arbor, MI) for 7 days. Stable recombinant cells of each group (e.g., group 1: A94T, group 2: P84L, group 3: WT, and group 4: GFP) were cultured in DMEM/high-glucose medium (Hyclone) supplemented with 10% fetal bovine serum (Gibco), 100 U/mL penicillin, and 100 μg/mL streptomycin. The plates were placed in a humidified incubator with 5% CO2 at 37°C.
Enzyme-linked immunosorbent assay
The levels of TNF-α, TGF-β1, and IL-6 in the medium supernatant of stable recombinant cells were determined using ELISA kits according to the manufacturer's instructions. Optical density was detected at 450 nm using a microplate reader (MR7000; Dynatech).
Cell viability assay
In brief, stable transfected cells were seeded into 96-well plates (5 × 103 cells/well) and incubated in a humidified incubator with 5% CO2 at 37°C. After 24, 48, 72, or 96 h, CCK-8 reagent (10 μL) was added to each well and the plates were incubated at 37°C for another 1 h. Absorbance of each group at 450 nm (OD450) was determined using a microplate reader (MR7000; Dynatech).
Real-time PCR assay
Cells were harvested when transfected LX-2 cells were at ∼80% confluent. Total RNA was isolated using TRIzol reagent, and cDNA was synthesized according to the Prime Script RT reagent kit. RT-qPCR was performed using SYBR® Premix Ex Taq™ and a real-time PCR system (Applied Biosystems 7500). Levels of mRNAs encoding TNF-α, α-SMA, collagen type I, and matrix-metalloproteinase (MMP) 2 or 9 were normalized to levels of GAPDH mRNA, and relative expression was calculated according to the formula: 2−ΔΔCT. The following primers were used (Table 1).
Name of Genes, Accession Numbers, and Nucleotide Sequences of the Primers Used for Real-Time PCR
Western blot analysis
When transfected LX-2 cells were at ∼80% confluence, cells were harvested and lysed for 30 min at 4°C in RIPA buffer containing cocktail, phenylmethanesulfonyl fluoride, and phosphorylated protease inhibitors A and B. Total protein concentration was quantified using the BCA protein assay kit according to the manufacturer's protocol. Equivalent amounts of proteins were separated on 10–12% SDS-PAGE and transferred onto PVDF membranes. Membranes were blocked with 5% bovine serum albumin for 2 h, washed three times with Tris-buffered saline containing Tween-20 (TBST). Then membranes were incubated overnight at 4°C with primary antibodies against TNF-α (1:500), P65 (1:1000), P-P65 (1:1000), IKB-α (1:1000), P-IKB-α (1:1000), and β-actin (1:1000). After washing three times in TBST, membranes were incubated for 2 h with antirabbit secondary antibody IgG H + L (1:10,000). Levels of target proteins were normalized to levels of β-actin and quantified using the Odyssey system (LI-COR Biosciences).
Statistical analyses
Data are presented as mean ± standard deviation. Statistical analyses were determined using Student's t-test and one-way ANOVA by SPSS 17.0 (IBM, Chicago, IL). P < 0.05 was considered statistically significant.
Results
TNF-α SNPs decrease the mRNA and protein level of TNF-α
TNF-α mRNA level in WT group was significantly higher than that in GFP group (Fig. 1A). A94T or P84L polymorphisms downregulated the mRNA level of TNF-α compared with WT group, meanwhile the inhibiting effects of A94T was more obvious than P84L. These results of Western blot (Fig. 1B) and ELISA (Fig. 1C) suggested that the reduction of A94T or P84L polymorphisms on the protein expression of TNF-α was in accordance with the findings of real-time PCR. Results showed that A94T or P84L polymorphisms reduced the content of TNF-α, which may be due to the destruction of the hydrogen bond network in the protein structure by the amino acid substitution caused by SNPs (Dabhi and Mistry, 2014).
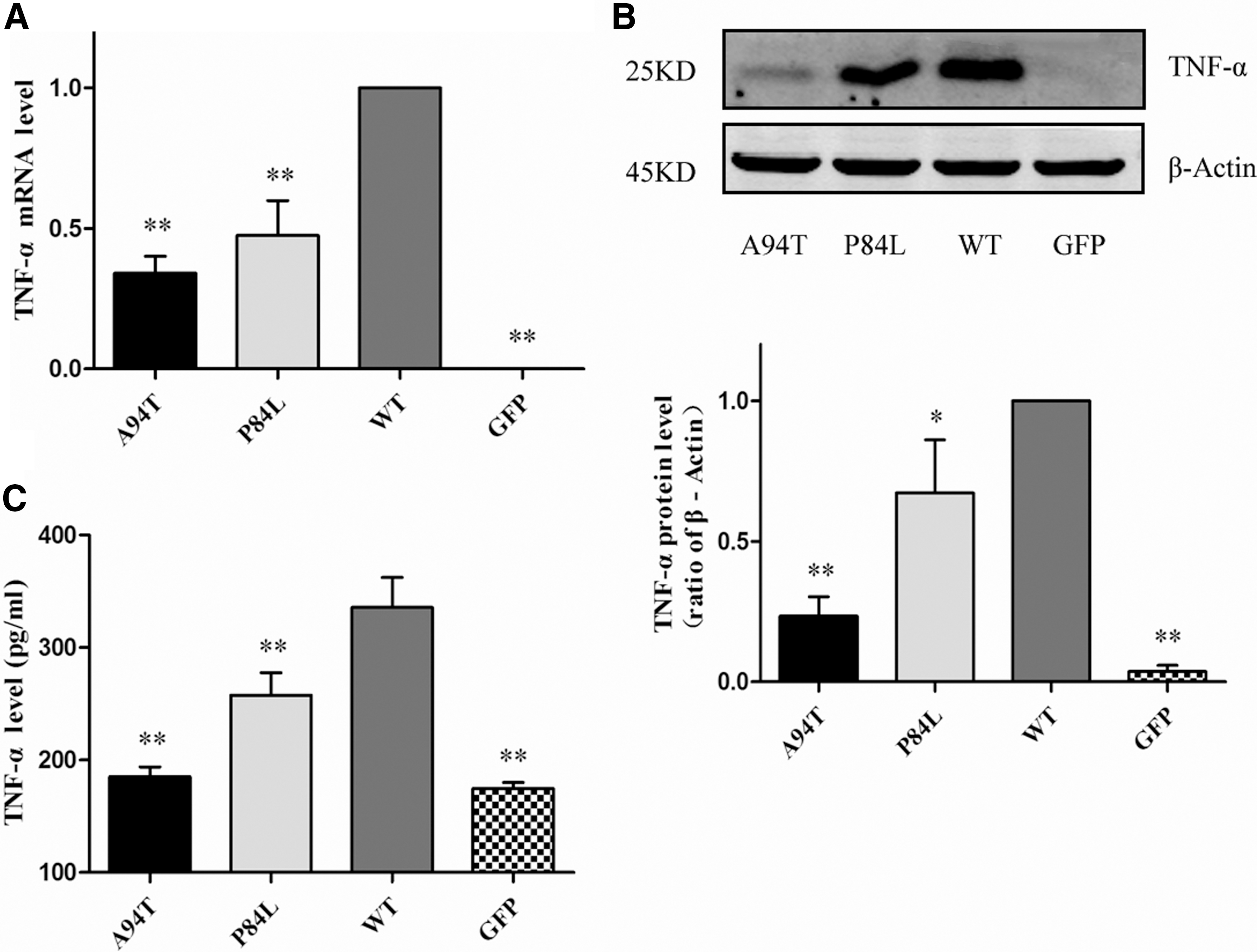
Effects of TNF-α SNPs on the mRNA and protein level of TNF-α.
TNF-α SNPs decrease the growth inhibitory effects of TNF-α on LX-2 cells
Because of the important role of A94T or P84L polymorphisms, the present study analyzes the effects of TNF-α SNPs on HSCs proliferation by CCK-8. Results are as shown in Figure 2, the absorbance of WT group was significantly lower than that of GFP group in 72 or 96 h, indicating that TNF-α could inhibit the growth of HSCs in accordance with previous studies (Gallois et al., 1998; Saile et al., 1999). What is more, OD value in A94T or P84L group significantly increased compared with that in WT group in 72 or 96 h. It means that A94T or P84L polymorphisms of TNF-α could significantly decrease the growth inhibitory effects of TNF-α on LX-2 cells.
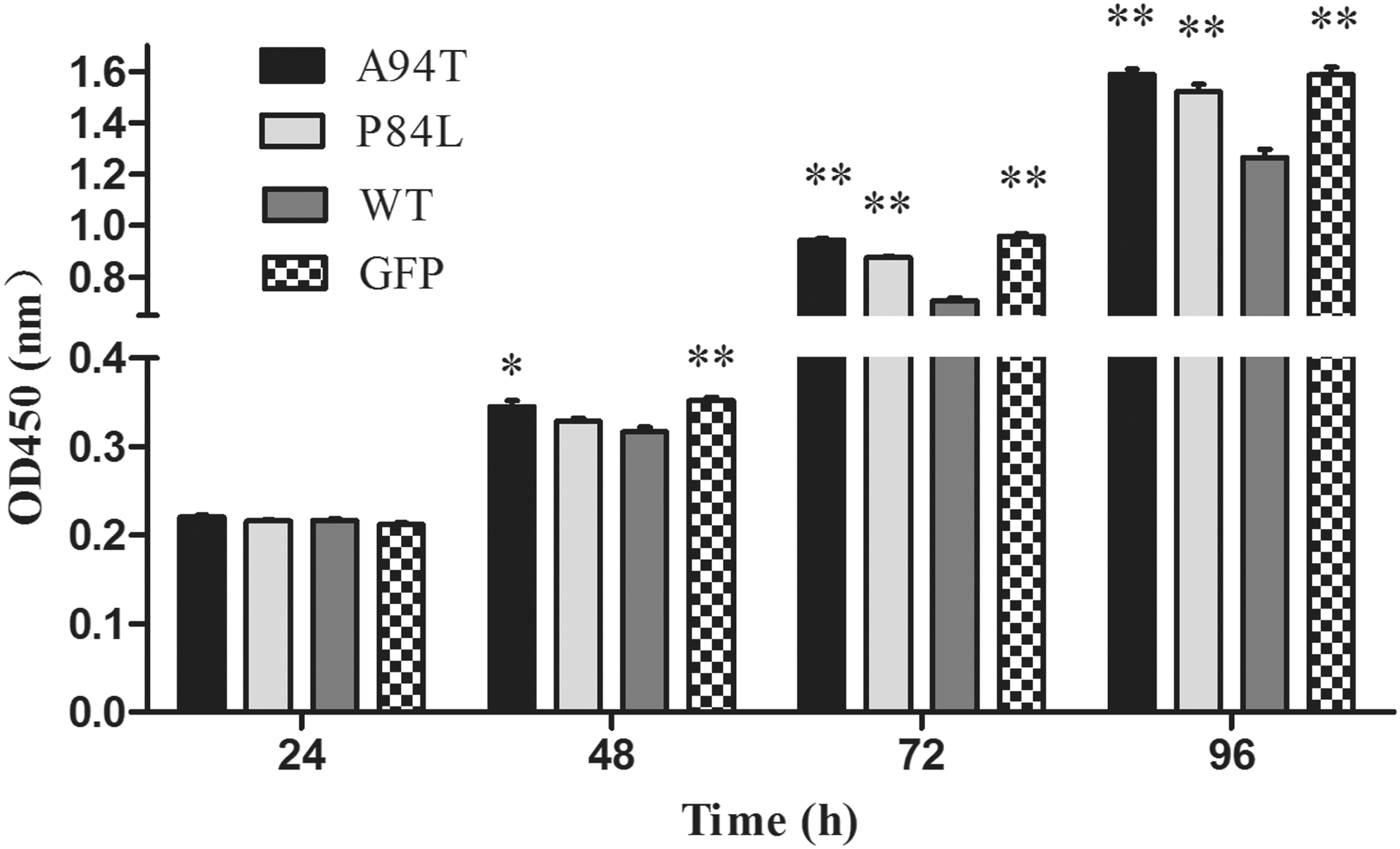
Effects of TNF-α SNPs on HSCs proliferation. Stable recombinant cells were seeded into 96-well plates (5 × 103 cells/well), HSCs proliferation were determined by CCK-8 after 24, 48, 72, or 96 h. Each column shows mean ± SD. Significance was set at *p < 0.05; **p < 0.01 versus WT group. CCK-8, Cell Counting Kit-8; HSCs, hepatic stellate cells.
TNF-α SNPs decrease the activation inhibitory effects of TNF-α on HSCs
Considering the effects of TNF-α SNPs on TNF content and the activation inhibition of TNF-α, we investigated the effects of TNF-α SNPs on HSCs activation. Results are as shown in Figure 3, WT group downregulated mRNAs encoding α-SMA and collagen type I, which are markers of HSCs activation, compared with GFP group. Furthermore, A94T or P84L polymorphisms significantly increased the mRNA level of α-SMA and collagen type I compared with WT group. A94T polymorphisms, but no P84L, increased the mRNA level of α-SMA compared with WT group. It means that A94T or P84L could decrease the inhibitory effects of TNF-α on collagen type I.
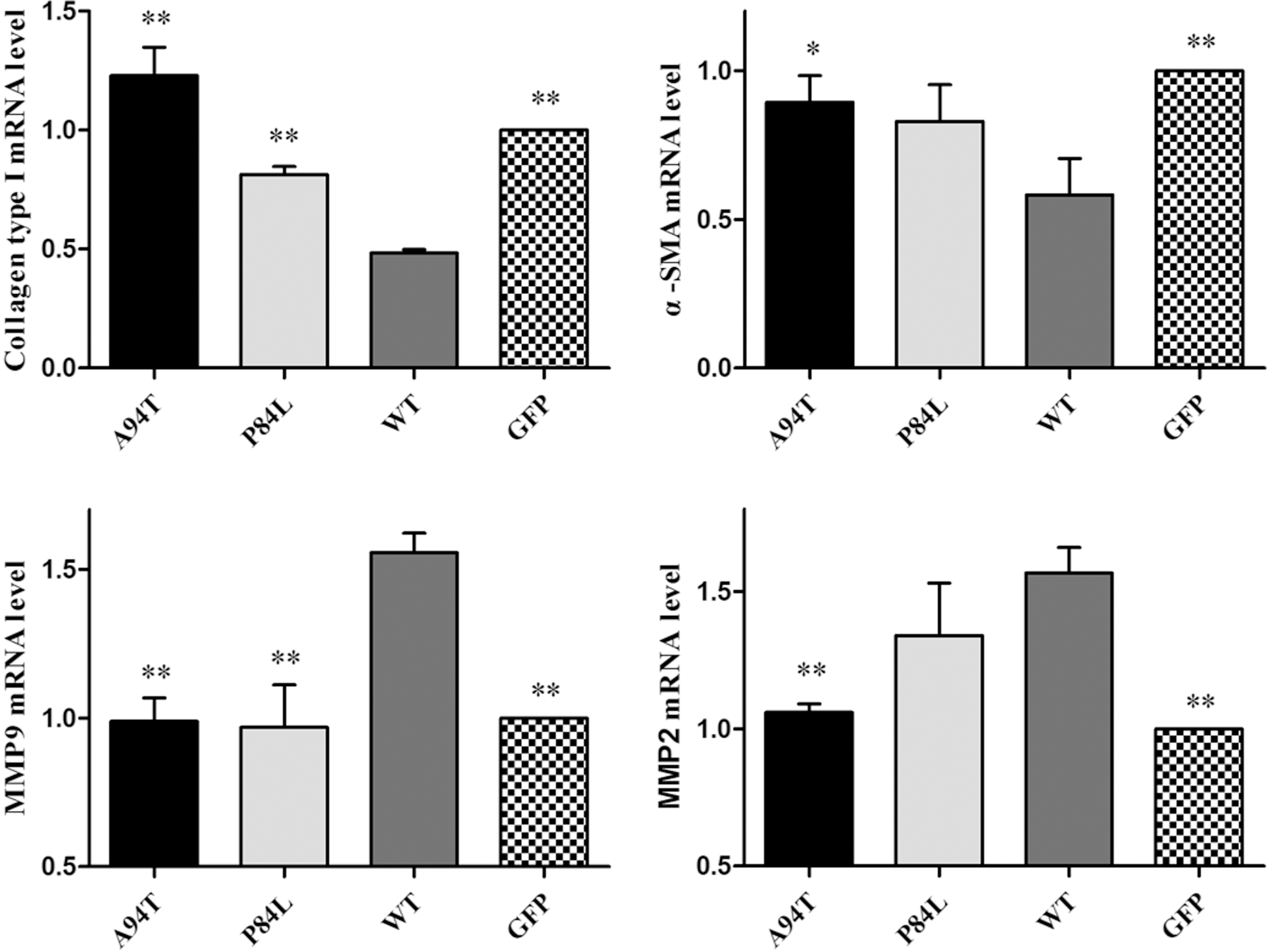
Effects of TNF-α SNPs on mRNAs encoding α-SMA, collagen type I, MMP 2, or MMP 9. The mRNA level of α-SMA, collagen type I, MMP 2, or MMP 9 was measured in stable recombinant LX-2 cells by real-time PCR. Each column shows the mean ± SD. Significance was set at *p < 0.05; **p < 0.01 versus WT group. MMP, matrix-metalloproteinase.
Apart from ECM synthesis, HSCs are also correlated with ECM degradation by production of MMPs. Compared with the mRNA level of MMP 2 or 9 in the GFP group, the mRNA level of MMP 2 or 9 was significantly increased in the WT group. What is more, mRNA encoding MMP 9 in A94T or P84L was significantly lower than that in WT group. A94T, but no P84L, decreased the mRNA level of MMP 2 compared with that in WT group. The experimental results showed that TNF-α SNPs could downregulate the expression of MMPs.
TNF-α SNPs decrease the promoting effects of TNF-α on IL-6
In subsequent experiments, we explored effects of TNF-α SNPs on IL-6 or TGF-β1. Results are as shown in Figure 4, there was no significant difference in the mRNA level of TGF-β1 among the four groups. We found that WT group upregulated IL-6 production compared with GFP group, but A94T or P84L polymorphisms significantly decreased the expression of IL-6, indicating TNF-α SNPs could downregulate the expression of IL-6.

Effects of TNF-α SNPs on IL-6 or TGF-β1. The expression of IL-6 or TGF-β1 was measured in stable recombinant LX-2 cells by ELISA. Each column shows the mean ± SD. Significance was set at *p < 0.05; **p < 0.01 versus WT group. TGF-β1, transforming growth factor beta 1.
TNF-α SNPs may induce HSCs activation through NF-κB pathway
To investigate the molecular mechanism of HSC activation induced by TNF-α SNPs, the protein expression of NF-κB pathway, were detected by Western blot. The experimental results (Fig. 5) showed that there was no significant difference in the level of total P65 protein among the groups. The protein expression of P-IκB-α and P-P65 significantly upregulated in the WT group and inhibitory kappa B-alpha (IκB-α) protein markedly decreased compared with those in the GFP group, indicating that TNF-α induces the activation of NF-κB pathway consistent with other studies (Novitskiy et al., 2005; Varela-Rey et al., 2007). In addition, TNF-α SNPs significantly inhibited the activation of NF-κB by upregulating the protein level of IκB-α and downregulating the protein level of P-IκB-α and P-P65, suggesting that the activation-promoting effects of A94T or P84L polymorphisms on HSCs activation may be related to NF-κB signaling pathway.
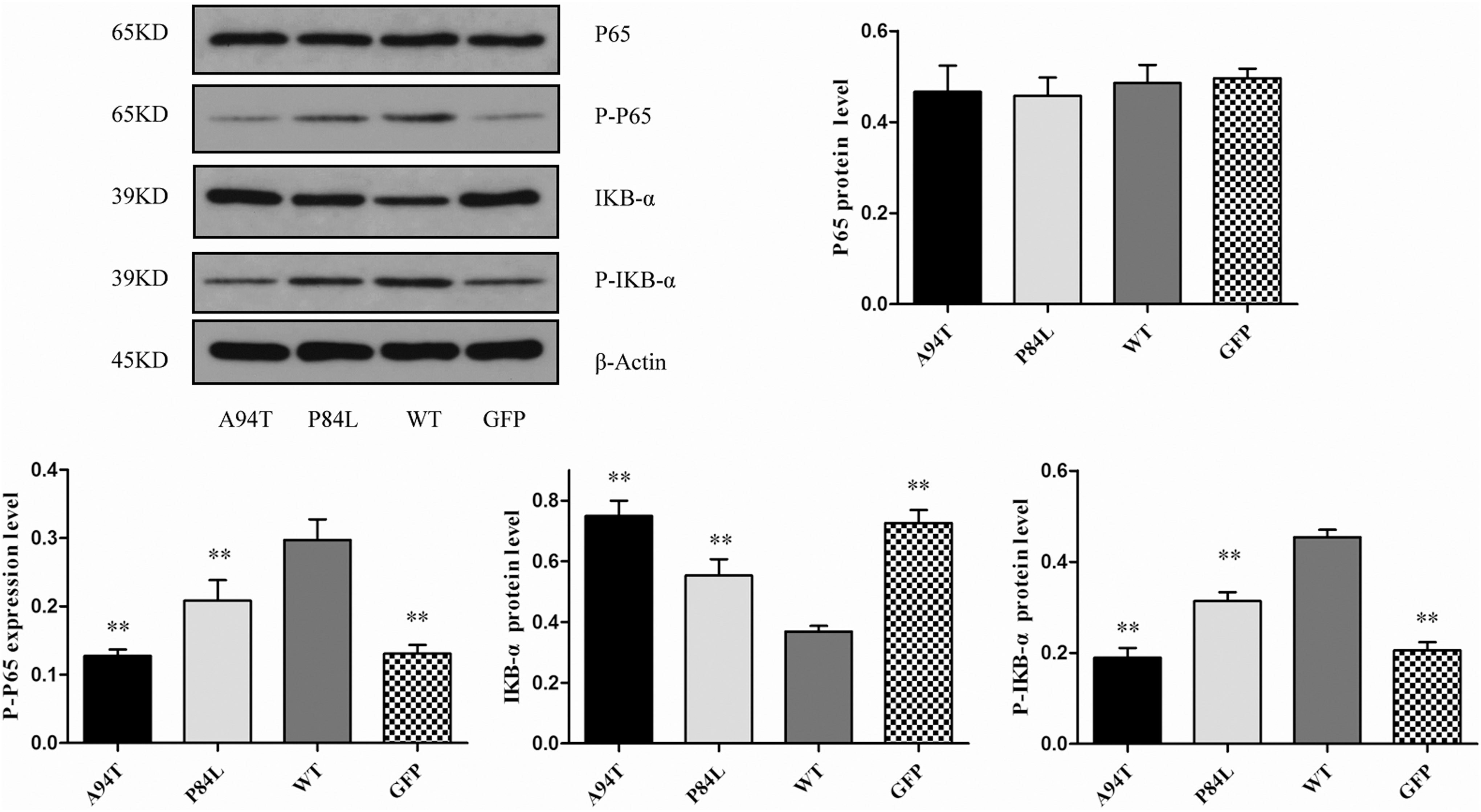
Effects of TNF-α SNPs on NF-κB pathway. The protein expression of P-IκB-α, IκB-α, P65, and P-P65 was measured in stable recombinant LX-2 cells by Western blot. Each column shows the mean ± SD. Significance was set at **p < 0.01 versus WT group. IκB-α, inhibitory kappa B-alpha.
Discussion
This study explores the effects of TNF-α A94T and P84L polymorphisms on the level of TNF-α, proliferation, and activation of HSCs. The experimental results showed that TNF-α SNPs downregulate the level of mRNA and protein of TNF-α, promote HSCs proliferation, promote HSCs activation, upregulate the level of MMP 2 or 9, and decrease the expression of IL-6. We also found that TNF-α SNPs promoted HSCs activation by blocking the activation of NF-κB signaling pathway. This study suggested that A94T and P84L polymorphisms decrease the content of TNF-α and reverse the antifibrosis effect of TNF-α.
It has been observed that all SNPs were located in the TNF-α promoter region. TNF-α enhancer polymorphism influences the transcriptional activity (Kroeger et al., 1997; Wilson et al., 1997), production (Louis et al., 1998), and the serum level of TNF-α, thus affecting the susceptibility to liver diseases (Jeng et al., 2007; Wang et al., 2012; Sghaier et al., 2015). However, many studies produced inconsistent conclusions (Brinkman et al., 1995; Kroeger et al., 2000; Mekinian et al., 2011) caused by small sample size, differences of patient race, lifestyle, and testing methods. Functional SNPs that change the level or structure of TNF-α are the basis of subsequent functional or case–control studies. It is advisable to find out the vital functional SNPs using computational prediction tools and TNF-α dbSNP (Dabhi and Mistry, 2014). Two variants A94T and P84L of TNF-α that were found could directly or indirectly destabilize the amino acid interactions and hydrogen bond networks, thus decreasing the stability of protein structure. In addition, stable recombinant cells, which overexpressed wild-type or the mutation of gene, could effectively explore effects of SNPs on the level of targets gene (Guo et al., 2009; Wei et al., 2016; Huang et al., 2017). So, this study using stable recombinant cells found that two variants A94T and P84L of TNF-α could downregulate the content of TNF-α, which was consistent with Dabhi B's findings. The results enhanced our understanding of the effects of TNF-α SNPs on the level of TNF-α.
The central events of hepatic fibrosis are proliferation and activation of HSCs that are targets of a number of cytokines currently under investigation. Several studies documented that TNF-α mediated inhibitory effects of proliferation in rat (Saile et al., 1999) or human HSCs (Gallois et al., 1998). Taking into account the important role of TNF-α SNPs and the antiproliferation effects of TNF-α on HSCs, this study analyzes the effects of TNF-α SNPs on HSCs proliferation. The results showed that TNF-α inhibited the proliferation of HSCs, which is consistent with previous studies (Knittel et al., 1997; Gallois et al., 1998). We found that TNF-α SNPs decrease the growth inhibitory effects of TNF-α on LX-2 cells by reducing the level of TNF-α.
In addition, HSCs activation plays an important role in hepatic tissue repair, which is characterized by deposition of ECM. A large number of studies have shown that TNF-α inhibits HSCs activation and downregulates the expression of collagen type I in vitro (Varela-Rey et al., 2002, 2007; Novitskiy et al., 2005) and in vivo (Buck et al., 1996). This study suggested that TNF-α inhibited the activation of HSCs by downregulating the level of α-SMA and collagen type I, which was consistent with previous studies (Armendariz-Borunda et al., 1992; Gallois et al., 1998). What is more, A94T or P84L significantly decreased the inhibitory effects of TNF-α on collagen type I. Except for ECM synthesis, HSCs are also correlate with ECM degradation by production of MMPs. It has been found that TNF-α could downregulate the level of collagen type I, accompanied by upregulation of MMP 1, 3, or 9 (Knittel et al., 1997; Robert et al., 2016). We found that both A94T and P84L of TNF-α could downregulate the mRNA level of MMP 9 compared with WT group. A94T, could decrease the expression of MMP 2 mRNA, but P84L couldn't. P84L may affect the interaction between TNF-α and subsequent target molecules, which needs further study.
Dabhi and Mistry (2014) found that the two identified TNF-α variants, that is, P84L and A84T, may alter the inflammatory signaling cascade. Cytokines, including growth factors and inflammatory factors, are involved in the process of hepatic fibrosis, and interact with each other (Robert et al., 2016). Several published articles showed that TGF-β1 could promote HSCs activation and inhibit the expression of TNF-α. In addition, TNF-α decreases collagen production and downregulates the expression of TGF-β1, indicating that there is interaction between TGF-β1 and TNF-α (Buck et al., 1996; Robert et al., 2016). The results showed that A94T and P84L polymorphisms downregulate the level of IL-6 consistent with Robert's study (Robert et al., 2016). However, there was no significant alteration in the mRNA level of TGF-β1 among all groups, which is inconsistent with other studies (Armendariz-Borunda et al., 1992; Buck et al., 1996; Robert et al., 2016). The reason for the contradictory conclusion may be that the construction of the overexpression vector is a stimuli to the cell itself, which needs further study.
Under normal physiological conditions, NF-κB consists of homo- and heterodimers of p50 and p65 protein units and combines with IκB-α, thereby maintaining NF-κB in a quiescent state. Stimuli results in degradation of IκB-α and activation of NF-κB, which translocates to the nucleus and regulates the transcription of target gene (Novitskiy et al., 2005). Many studies (Mann and Smart, 2002; Novitskiy et al., 2005; Varela-Rey et al., 2007) show that NF-κB pathway is closely related to liver fibrosis and HSCs activation. Novitskiy et al. (2005) and Varela-Rey et al. (2007) found that TNF-α-inhibited HSCs activation may be related to the activation of NF-κB pathway by promoting the phosphorylation of IκB-α. In addition, Varela-Rey et al. (2002) found that the activation inhibiting effects of TNF-α was involved in p38 MAPK pathway but not in Jun-N terminal kinase (JNK) or in extracellular-regulated kinase (ERK). This study suggested that TNF-α SNPs significantly upregulated the protein level of IκB-α, downregulated the expression of P-IκB-α, and P-P65 eventually inhibited the activation of NF-κB. The results showed that A94T or P84L polymorphisms inhibited HSCs activation that may be related to NF-κB signaling pathway.
Conclusion
In summary, the present study found that A94T and P84L polymorphisms downregulate the level of TNF-α. NF-α SNPs could inhibit the proliferation and activation of HSCs. The activation inhibiting effects of TNF-α SNPs is related to NF-κB pathway. This study provides two vital SNPs for functional or case–control studies of TNF-α SNPs and provides susceptibility sites for liver-related diseases.
There are limitations to this study. Our study only revealed that SNP could reduce the level of TNF-α, but did not verify the effects of A94T and P84L polymorphisms on the structural stability of TNF-α. Therefore, future studies should verify the effects of TNF-α SNPs on the protein structure of TNF-α. Our future study will enhance understanding of TNF-α SNPs and their role in liver fibrosis.
Footnotes
Acknowledgments
The authors thank their colleagues at the Central Laboratory of Renmin Hospital, Wuhan University, where most of this work was performed.
Disclaimer
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Disclosure Statement
No competing financial interests exist.